|
|
||||||
|
|
|
|
(en travaux) |
|||
|
Plan |
|
|
page simplifiée pour les secondes : Les mutations |
|||
| |
|
|
||||
|
Introduction : des mutations très instrumentalisées |
||||||
|
|
Popularisé par les séries télévisées (des X-files ou X-men en passant par les Spiderman et autres comics....) l'idée de mutants - et surtout de mutants humains - a d'abord été un simple prolongement de science-fiction des interprétations de l'hérédité mendélienne et morganienne : une mouche avec une paire d'ailes supplémentaires, une bactérie capable de résister à un antibiotique... L'imaginaire s'est vite déchaîné et l'on en est venu à considérer que les élucubrations de la science-fiction n'étaient peut-être pas si éloignées que cela d'un futur proche. Ceci une idée scientifiquement erronée. |
|
Il y a de quoi s'alarmer devant la naïveté de certains raisonnements d'élèves, ou même de pédagogues qui s'appuient sur les programmes de l'enseignement secondaire. Ceux-ci, en effet, véhiculent des visions simplistes sur le programme génétique ou sur le programme de développement. On a tenté, depuis les années 2000, de modérer ce travers à l'aide des termes "environnement" et "épigénétique" , mais sans grand succès, étant donné que l'on est resté, dans la plupart des cas, à la présentation d'un être vivant comme machine moléculaire dont le comportement est déterminé par un programme (génétique) sous le contrôle de l'environnement (épigénétique). Il y a une profonde méconnaissance de la richesse de la recherche en épigénétique et de la remise en cause de la théorie de l'information génétique - dans sa version scolaire - qu'elle suppose (voir quelques pistes sur la biologie systémique ou encore la biologie autonome dans la page de biologie théorique et celle sur la science). |
|
http://epigenomique.free.fr/fr/index.php épigénèse/préformation... une riche histoire de ces mots (voir remarque en bas de page)
Dans le cadre de la théorie de l'information génétique, la mutation est un changement de l'information génétique (au sens le plus restreint de séquence de l'ADN), qui détermine donc (par causalité matérielle) un changement plus ou moins important, voire non apparent, chez l'être vivant. |
|
|
|
D'une façon paradoxale, cette vision, même reconnue comme inexacte, est, à mon avis, volontairement colportée par ceux qui y trouvent un appui pour leur attitude philosophique de rejet de la notion de cause (voir les 4 causes) en s'efforçant d'y substituer un hasard (qui joue en fait le rôle de la fin). Ils s'appuient en effet sur le hasard (des erreurs non corrigées...) pour expliquer l'apparition des mutations mais invoquent le plus souvent un déterminisme (qui diffère de la causalité) pour le passage du génotype au phénotype (il y a des exceptions notamment la vision probabiliste de Kupiec, mais sa vision reste, je crois, un matérialisme qui rejette toute causalité autre que matérielle). Il est peu crédible de donner un sens à l'information génétique, tout en refusant d'en donner un à la variation. |
|
Une
autre raison
plus politique
et surtout
économique
des efforts de
certains pour
favoriser
cette vision
étroite
de la mutation
repose sur
l'implication
de ses
promoteurs
dans la
technologie
génétique
: recherche de
gènes
impliqués
dans telle ou
telle maladie,
manipulations
génétiques
et surtout
OGM... la
théorie
de
l'information
génétique
est
défendue
par ceux qui
ont
réalisé
des
investissements
colossaux et
se sont
engagés
dans une
entreprise
parfois
très
immorale dans
le but de
contrôler
l'alimentation
et la
santé
des hommes. |
|
Sans entrer dans l'aspect politique, je vais m'attacher à corriger, scientifiquement, cette manière stérilisante de présenter la mutation, particulièrement faussée, historiquement et expérimentalement. Je présenterai quelques ouvertures après un essai de panorama "historique". |
|
|
|
||||||
|
1 - Une histoire en 3 étapes : mutations héréditaires, géniques et chromosomiques |
||||||
|
Les étapes sont une sorte d'artifice de présentation (je n'ai pas de prétention historique - au sens d'un véritable travail d'histoire des sciences- sur cette question complexe). |
|
1.1 la mutation comme changement brusque héréditaire |
||||
|
ancienne page sur l'histoire de la génétique cours
de terminale
de
spécialité |
|
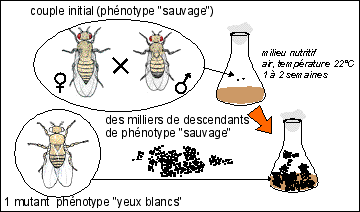
Il semble que ce soit Hugo de Vries (1848-1935) (qui travaille sur Œnothera lamarckiana) qui utilise la première fois le terme de mutation dans un sens biologique en nommant ainsi le changement brusque de caractères observé dans une descendance. Il développe la théorie selon laquelle les mutations président à l'apparition de nouvelles espèces. De Vries utilisera le terme de pangène pour désigner les particules héréditaires qui seront finalement nommées gènes (le sens actuel est différent sauf si on ajoute le qualificatif "héréditaire") selon la proposition du biologiste danois Wilhem Johannsen (1857-1927).
|
|
Je
ne crois pas
me tromper en
affirmant
qu'historiquement
le terme de
mutation
désigne
plutôt,
à
l'origine, un
phénomène
qui atteint
l'ensemble de
l'être
vivant
(qualifié
alors de
muté)
et non pas une
particule ou
une
molécule.
On notera qu'il est sous-entendu que ce nouveau caractère est lui-même héréditaire (héritable ou transmissible à la descendance donc stable). On voit donc combien on est là dans une théorie de la descendance. (voir page sur l'hérédité)
|
||
| |
|
|
Remarque
historique: Même si cette vision saltatoire de la mutation est aujourd'hui abandonnée, la théorie de De Vries a joué un grand rôle dans l'histoire du darwinisme. De nos jours ce type de mutation reste largement inexpliqué par la génétique moléculaire. |
|
«
Il est
très
étonnant
que les
diverses
mutations
Œnothères
offrent un
grand
degré
de
régularité;
il n'y a pas
de chaos de
formes, ni de
déviations
mal
définies
à tous
les
degrés
et dans toutes
les
directions; au
contraire, il
est
évident,
à
première
vue, que le
phénomène
tout entier
est
régi
par des lois
très
simples. (H.
De Vries,
Espèces
et
variétés,
351)» Histoire de la notion de vie, André Pichot, 1993, Gallimard, pp 921s |
|
| |
|
Il est important de noter combien la stabilité est associée à la variabilité dans cette vision héréditaire de la mutation. La mutation est un changement brusque et stable d'un caractère (ou d'un ensemble de caractères) héréditaire(s), apparu chez un nouvel être vivant à l'issue d'une reproduction. |
|
|
||
| |
|
1.2 la mutation comme variation allélique |
||||
|
théorie chromosomique de l'hérédité |
|
Sans que l'on puisse déterminer de façon nette, et surtout datée, un changement de sens, la mutation se comprendra ensuite, dans la théorie chromosomique de l'hérédité (weismannienne), comme un changement au niveau du gène (au sens de gène héréditaire bien sûr). |
|
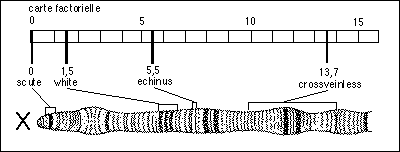
Les mutations sont la cause de la variation allélique: les nouveaux allèles (forme des gènes) apparaissent par mutation; ils sont ensuite transmis fidèlement par la reproduction sans altération. Tous les allèles, sauf les allèles sauvages, sont donc le résultat de mutations dans une telle théorie héréditaire (on voit que pousser ce raisonnement à l'extrême est peu convaincant). Pour les eucaryotes les gènes sont souvent notés avec une seule lettre (minuscule pour les allèles récessifs et majuscule pour les allèles dominants, le sauvage étant souvent agrémenté d'un + en exposant: par exemple, W+ indique la couleur rouge des yeux de la drosophile, w pour white, la couleur du mutant le plus fréquent). Pour les procaryotes les groupes de gènes intervenant pour un même produit sont nommés par trois lettres (par exemple lac), suivies par une lettre majuscule indiquant le gène (lac Z); l'allèle sauvage est lui aussi affecté d'un exposant + (lac Z+) et les allèles mutés par - ou d'autres sigles en exposant. |
|
La théorie chromosomique de l'hérédité (Weismann puis De Vries et Morgan...) repose donc sur un système stable de transmission héréditaire (les gènes, portés par les chromosomes) et un système de variation allélique (les mutations). Si le support matériel particulaire est affirmé (le chromosome comme groupe de liaison, voir cours de TS), le support moléculaire éventuel reste alors inconnu. Inséparables de la variation allélique, les réarrangements au sein des chromosomes après cassure (crossing-over et autres recombinaisons), sont parfois considérés comme des mutations chromosomiques (voir ci-après). |
|
|
|
|
||||
|
|
|
1.3 les mutations géniques dans la théorie de l'information génétique |
||||
|
|
|
|
source principale: Analyse génétique moderne, Griffiths et al., 2001, DeBoeck Université, ch 7 et 8 |
|||
|
voir la partie historique du cours de 1èreS vocabulaire pour les gènes héréditaires et moléculaires : annexe au cours de 1èreS |
|
Vers les années 1940, dans le cadre de la théorie de l'information génétique, les biologistes moléculaires vont tenter de fusionner le gène héréditaire et le gène moléculaire. Cette fois encore on superpose la stabilité de l'expression et de la transmission de l'information génétique à la variation (mécanisme supposé aléatoire, vis-à-vis de ce qui est supposé stable: l'information, représentée par la séquence du gène moléculaire, ce qui est très différent du caractère considéré dans la théorie chromosomique). |
|
Ces mutations géniques ont un sens construit a posteriori qui repose strictement sur la notion de gène moléculaire, avec, comme fondement matériel, la fragilité de l'ADN ou la variabilité des mécanismes cellulaires de synthèse ou de transcription de ce même ADN. Elles n'ont souvent de sens expérimental que pour les bactéries !!!! La généralisation à tout organisme vivant est problématique. La "première" (?) mutation historique présentée par De Vries chez Œnothera, sera associée à une recombinaison génétique par translocation. |
|
Les
mutations des
survivants
à
l'application
d'un agent
mutagène: |
|
La mutation génique est un changement de l'information génétique (au sens le plus restreint de la seule séquence de l'ADN), déterminant un changement plus ou moins important, voire non apparent, chez l'être vivant. |
|
Même si je pense qu'il est dépassé (je reporte la discussion au chapitre suivant), ce type d'explication n'est pas à rejeter totalement, surtout du fait de la cohérence du pouvoir explicatif du modèle proposé. Le passage d'une mutation comme changement phénotypique à une mutation comme changement génotypique s'est fait progressivement. Le terme est maintenant employé avec des sens différents et recouvrant parfois des visions incompatibles. |
|
Les paragraphes ci-dessous présentent différentes approches qui se superposent plus ou moins. |
||
| |
|
a
- les
mutations
selon leur
origine
lointaine |
||||
|
Il est d'usage de considérer deux types de mutations : |
|
*
Les mutations
spontanées
(sans
que l'on ai
soumis
l'organisme
à un
agent
mutagène)
sont
considérées
comme dues
à |
|
*
Les mutations
induites
(avec
soumission
à un agent
mutagène)
:
pouvant
être
dues |
|
Il va sans dire que ces deux types de mutations peuvent être naturelles étant donné que l'on considère qu'il existe dans la nature de nombreux agents mutagènes, à commencer par le rayonnement u.v.. Cette distinction repose donc plutôt sur l'observation de la fréquence des mutations. Une fréquence élevée laissant supposer la présence d'un mutagène. Cependant, certaines souches présentent naturellement une forte instabilité. |
| |
|
b
- les
mutations
selon leurs
conséquences
sur la
synthèse
des produits
génétiques
|
||||
|
Les substitutions dans les gènes de structure du protéome |
|
Les mutations silencieuses, quelle qu'en soit la cause, provoquent un changement de la séquence de l'ARNm qui ne se traduit, du fait de la redondance du code génétique par aucun changement dans l'expression du gène moléculaire (même acide aminé ou autre codon stop). Les mutations faux-sens conduisent au remplacement d'un aa par un autre. Les mutations non-sens conduisent à l'arrêt de l'allongement de la chaîne polypeptidique du fait du remplacement d'un codon correspondant à un aa par un codon stop. |
|
Les
conséquences
phénotypiques
de telles
mutations ne
sont pas
forcément
simples. |
||
|
Les autres mutations |
|
Les additions ou les délétions (ponctuelles ou non) de bases peuvent avoir des conséquences nettement plus importantes que les simples substitutions notamment dans le cas d'un décalage du cadre de lecture, tous les codons étant susceptibles d'être changés en aval d'une addition ou d'une délétion. |
|
Les conséquences des mutations dans les séquences régulatrices et autres séquences non codantes sont difficile à évaluer, même du point de vue théorique. Par exemple les mutations au niveau des sites d'épissage (splice-site : dans ce type de mutation des nucléotides sont insérés ou délétés juste au niveau des sites d'épissage, ce qui empêche le bon déroulement de cette phase de maturation des ARNm). |
|
|
| |
|
c - les réarrangements de l'ADN des procaryotes |
||||
|
Ces réarrangements sont parfois classés avec les "mutations chromosomiques", à tort, puisque le terme de chromosome est tout à fait inadapté pour les procaryotes. |
|
Du
fait des
moyens
investis et du
foisonnement
des
modèles
concernant
l'organisation
dynamique du
matériel
génétique
des
Procaryotes,
je ne
prétends
pas ici
être
à jour
des derniers
développements. Il faut d'abord souligner que l'ADN d'un Procaryote se présente souvent sous la forme de plusieurs (4 à 6 habituellement chez E. coli) boucles d'ADN sans compter les plasmides. Il y a donc une redondance de ce que l'on appelle l'information génétique. Mais savoir quelle information est utilisée n'est pas non plus évident : la présence de l'ADN ne suffit pas à prouver que cette information est exprimée. |
|
Finalement, quelle importance peut avoir l'ordre des gènes sur le nucléofilament ? Cette importance dépend surtout de la présence et de la position des séquences régulatrices ce qui fait que le gène ne serait pas la bonne unité de fonction. On revient à l'ancienne notion d'opéron qui a eu son succès, même si les choses se sont compliquées depuis. Les séquences régulatrices sont-elles spécifiques, mobiles,.. ? Les gènes peuvent-ils être fractionnés tout en étant actifs ? Pire encore, un gène disparu peut réapparaître dans le génome des descendants. À partir des ARN ? |
|
Comment distinguer une origine dans le filament d'ADN bactérien ? Il
y a cependant
des points de
cassure
préférentiels.
Quels sont
leur lien avec
les
gènes ? Quels sont les points d'attachement des nucléofilaments à la membrane/paroi ? Les échanges de matériel génétique sont très fréquents entre bactéries d'une même espèce mais aussi entre espèces différentes. Les virus participent aussi à ces échanges. Une homogénéisation génétique des populations est ainsi plus ou moins obtenue. |
| |
|
d -les réarrangements à l'échelle du chromosome (donc chez les eucaryotes) |
||||
|
Ces réarrangements sont la plupart du temps qualifiés de "mutations chromosomiques", ce qui se discute. |
|
D'abord interprétés dans le cadre d'une hérédité chromosomique morganienne (voir cours de spécialité) - un chromosome linéaire orienté (du fait de la position du centromère) avec des allèles disposés les uns à la suite des autres dans un ordre fixe dans l'espèce- , les mécanismes proposés permettent de rendre compte de la transmission héréditaire des allèles étudiés chez la drosophile et d'autres organismes modèles (Saccharomyces, Œnothera, Cænorhabditis...) , dans des cas particuliers. Dans l'étape suivante, celle de la théorie de l'information génétique, le chromosome sera ensuite assimilé à une unique molécule d'ADN. Des techniques développées avec les outils du génie génétique (voir cours de spécialité) permettront alors d'intégrer la génétique des procaryotes, avec plus ou moins de réussite. |
|
À l'inverse des mutations géniques considérées ci-dessus, le chromosome est bien pris ici comme support des gènes héréditaires (et non pas de l'information génétique). La mutation ne change pas l'information, mais provoque matériellement un changement dans la structure du chromosome supposée stable, allongée et orientée (de par la position d'un centromère décalé - l'orientation par les bandes colorées (banding) est aussi possible). Ce changement structurel a pour principales conséquences des changements statistiques dans la transmission des allèles (voir ci-dessus), sans que l'on s'intéresse au niveau moléculaire, du mois dans un premier temps. |
|
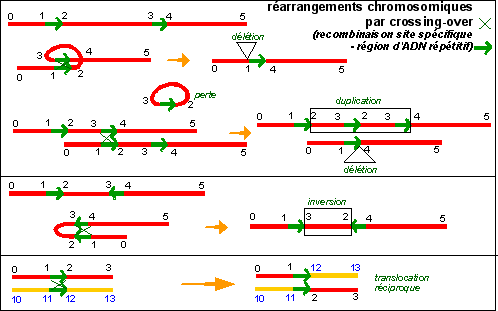
On considère que les cassures suivies d'une réunion sont extrêmement fréquentes, elles sont d'ailleurs à la base de la théorie du crossing-over, ou recombinaison site-spécifique. On pense que ces cassures sont pour la plupart indécelables. C'est dans le cas où la réunion se fait de façon différente d'avant la cassure que l'on peut; parfois, déceler des modifications. On fait, il serait préférable de dire que, la fragilité et la capacité des chromatides à se réunir à nouveau après cassure permet d'envisager des mécanismes explicatifs dont la plupart sont loin d'être autre chose que des hypothèses plus ou moins séduisantes. Même si l'on s'est efforcé de réunir les deux niveaux d'étude il n'est rien moins que très incertain que les sites de cassure / réunion des chromosomes proposés comme explication aient quelque chose à voir avec les sites de recombinaison observés dans l'ADN bactérien. |
| |
|
|
|
En travaux |
||
|
|
|
1.4 les variations caryotypiques cadrant mal avec une théorie des mutations |
||||
|
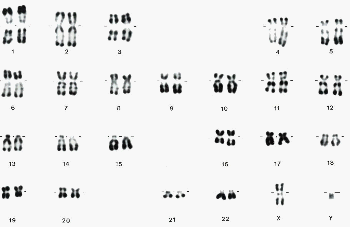
On qualifie habituellement, à tort, les anomalies du caryotype (nombre, taille, forme ou structure des chromosomes) de mutations chromosomiques
cours de seconde : les chromosomes |
|
Si les outils de la biologie moléculaire peuvent être employés pour s'efforcer de comprendre ces phénomènes, il est cependant important de souligner que l'on se place ici à une autre échelle que celle de la molécule. Ici ce sont des corps colorables micrométriques: les chromosomes, que l'on peut observer au microscope (mais souvent très difficilement...voir par exemple Acetabularia qui est loin d'être un cas isolé), marquer avec des sondes fluorescentes, voir, même, manipuler. La théorie mise en jeu dans ces phénomènes va avoir beaucoup de peine à intégrer l'information génétique issue de la biologie moléculaire du gène. Si l'on trouve parfois le terme de cytogénétique, il faut alors bien remarquer que ce terme ne désigne pas une génétique centrée sur l'ADN, et encore moins sur l'ADN procaryotique, mais une cytologie du matériel génétique eucaryote. |
|
Les anomalies caryotypiques ne correspondent pas à des changements brusques de caractères dans une descendance car tout d'abord, elles ne sont pas héréditaires, du fait de la stérilité qui les accompagne souvent lorsqu'elles touchent les cellules sexuelles. Ensuite, les modifications "structurales" que l'on fait porter la plupart du temps sur l'ADN contenu dans le chromosome (alors que le contenu protéique du chromosome est majoritaire dans la structure colorable) sont fortement tributaires de la représentation que l'on a de cette structure: voir le chromosome comme une simple pelote d'ADN dont le seul rôle est de contenir deux immenses molécules d'ADN force implicitement à fusionner la génétique chromosomique avec la génétique moléculaire. Ce n'est certes pas la seule voie et nous choisirons bien sûr ici de présenter le chromosome comme une structure originale (composée d'ADN et de protéines mais surtout comme une forme dynamique) qui ne se résume pas au modèle moléculaire. Une difficulté supplémentaire, d'ordre méthodologique , vient des techniques de marquage chromosomique - ces techniques utilisent uniquement l'ADN comme cible des marqueurs- . Elles fusionnent donc implicitement les représentations cytologiques et moléculaires, souvent de façon indue. Le marquage d'un gène moléculaire, parfois possible directement sur un chromosome traité, fait incontestablement penser au chromosome comme à une pelote de gènes, ce qui laisse de côté les aspects dynamiques proprement structuraux. |
||
| |
|
|
|
Les variations du nombre de chromosomes ne sont pas des mutations |
||
|
Le nombre de chromosomes est normalement fixe dans une espèce. |
|
La
plupart des
organismes
possèdent
, au moins
lors d'une génération
de leur
cycle de
développement,
plus d'un jeu
de chromosomes
: on les
qualifie d'euploïdes
(2n=diploïdes,
3n=triploïdes,4n=tétraploïdes,
5n=pentaploïdes,
6n=hexaploïdes,
etc.). Des individus monoploïdes apparaissent aussi chez certaines espèces (gamétophyte de très nombreuses plantes, reproducteurs mâles chez les abeilles par exemple...). |
|
Un exemple typique de polyploïde naturel est représenté par le blé; l'espèce, utilisée majoritairement pour la panification est Triticum æstivum, un hexaploïde naturel issu de la fécondation entre une espèce tétraploïde (Triticum dicoccum) et une espèce diploïde (Aegilops squarrosa ou Æ. ovata ou Triticum tauschii). on parle d'allopolyploïdie (voir ancienne page sur le blé, dont on a pu retracer l'histoire génétique probable depuis sa domestication). Mais l'on peut manipuler le nombre de chromosomes d'un individu (que l'on reproduit ensuite) dans un but commercial. Par exemple, de nombreux individus triploïdes (toujours stériles) ont été obtenus à partir du croisement d'une espèce tétraploïde avec une espèce diploïde. |
|
Chez certaines espèces on peut augmenter le degré de polyploïdie par fécondation entre individus de polyploïdie différents (que l'on trouve naturellement dans les cultures) mais de la même espèce (autopolyploïdie), comme chez le tabac. On a noté qu'habituellement la plante (et ses cellules) était alors d'autant plus grande que son degré de ploïdie était élevé. On a aussi fabriqué des polyploïdes artificiels en augmentant la ploïdie de gamètes par l'utilisation de la colchicine (voir les expériences de Taylor pour l'utilisation de cette toxine). |
| |
|
De la même manière il est possible de réaliser des monoploïdes artificiels par culture de grains de pollen (androgénèse) qui, pour certaines espèces, peuvent conduire à des plants complets monoploïdes (soja, tabac...). On peut ainsi sélectionner certains plants directement à partir de la culture in vitro de cals appelés embryoïdes (par exemple pour la résistance à telle ou telle substance). On peut ensuite passer du monoploïde au diploïde par utilisation de la colchicine (ou un autre bloquant partiel de la séparation anaphasique des chromatides) lors des premières divisions. |
|
Ces techniques, très aléatoires, ont cependant donné des résultats probants pour la sélection de plants destinés à l'agriculture "contrôlée". Il va sans dire que la stabilité problématique de leurs caractères et la nécessité de réaliser à nouveau les manipulations pour chaque génération, du fait de la stérilité des hybrides, font que ces manipulations sont d'une tout autre nature que celles réalisées par les sélectionneurs qui travaillaient uniquement à partir des semences constituées de plantes matures. |
|
Les cas de polyploïdie ne sont pas du tout réservés au royaume des plantes. On trouve de nombreux animaux polyploïdes. Tout le monde connaît la parthénogenèse du puceron ou des abeilles, mais il est plus rare que l'on sache que de nombreux invertébrés, poissons, amphibiens et surtout reptiles peuvent présenter habituellement des polyploïdies, plus ou moins stables. |
| |
|
Les aneuploïdies, anomalies dans le nombre de chromosomes, touchent une ou quelques paires de chromosomes seulement. Les plus connues sont, chez l'homme, les trisomies dont seules les 21, 13 et 18 sont viables, au moins jusqu'à la naissance (130 jours de vie moyenne pour les enfants atteints de trisomie 13 ou syndrome de Patau). La trisomie 21 ou syndrome de Down fut la première maladie à être associée à un caryotype anormal par l'équipe du professeur Lejeune. On est loin de bien comprendre la cause des symptômes des maladies associées à ces trisomies qui sont toutes associées à un retard mental. |
|
La théorie la plus courante concernant la cause des anomalies phénotypiques associées aux anomalies chromosomiques reprend la théorie de l'information génétique en supposant qu'il existe un déséquilibre dans le dosage des produits associés aux gènes présents sur les chromosomes excédentaires. Cette thèse est assez mal confirmée par les récents dosages réalisés massivement (à l'aide de puces) sur des cellules trisomiques qui ne montrent que de faible taux de surexpression (voir de baisse du niveau d'expression) des gènes présents en 3 exemplaires dans la cellule trisomique. Il me semble bien plus prometteur de s'intéresser à des explications faisant intervenir le niveau supérieur d'organisation dans la cellule. Les anomalies chromosomiques se retrouvent fréquemment associées à des cellules cancéreuses et peuvent même être transmises entre organismes adultes comme une maladie infectieuse (voir page sur les chromosomes - partie 6). |
|
On peut aussi remarquer que les anomalies concernant les chromosomes sexuels observées chez une vingtaine d'espèces de mammifères dont l'homme (XO, XXX pour des femelles, souvent stériles, et XXY et XYY pour des mâles, toujours stériles) sont loin d'être généralisables. De plus, il existe de très nombreuses espèces - y compris de vertébrés - chez lesquelles la présence de chromosomes sexuels n'a pu être mise en évidence (reptiles (tous les crocodiliens, nombreuses tortues, quelques lézards), amphibiens (plus de 99%), poissons (la plupart)). |
|
voir aussi page sur l'identité |
|
Ainsi, les euploïdies naturelles stables ne peuvent guère être considérées comme des mutations, car elles ne modifient pas vraiment les caractères d'un individu (ou le font de façon harmonieuse). D'autre part, les aneuploïdies ne sont pas transmissibles héréditairement et ne correspondent pas à un changement stable. |
|
Au niveau cellulaire, une cellule cancéreuse, présentant une aneuploïdie, est certes anormale, et peut transmettre dans certains cas cette anomalie aux cellules issues de sa division, mais il ne s'agit pas d'hérédité au niveau de l'organisme. Le terme de mutation cellulaire ne peut être employé que de façon plus ou moins analogique. |
|
Il n'y a donc pas vraiment de raison sérieuse pour utiliser le terme de mutation pour les variations du nombre de chromosomes. |
| |
|
|
|
Les variations caryotypiques fines |
||
|
délétions |
|
Les délétions (du latin deletere=détruire), ou pertes ne sont pas toutes de même ampleur. Les macrodélétions peuvent aller jusqu'à toucher l'ensemble d'un bras d'un chromosome. Les fragments contenant le centromère sont aussi parfois perdus ce qui empêche l'accrochage des fibres kinétochoriales et le déplacement du chromosome à l'anaphase. Les cellules résultant de la division n'auront pas alors le même caryotype. Les macrodélétions sont parfois mises en évidence par des boucles (boucles de délétion) que font les chromosomes appariés lors de la 1re prophase de la méïose. La zone non appariée formant une boucle que l'on peut observer au microscope. C'est aussi le cas dans les chromosomes polyténiques où de telles boucles, très fréquentes, sont interprétées comme des boucles de délétion. |
|
On peut mettre en évidence des modifications plus ou moins fines dans l'organisation des chromosomes à l'aide des techniques de coloration des chromosomes en bandes claires et sombres (banding, voir fiche chromosomes). Le syndrome le plus connu est celui du "cri-du-chat", nommé ainsi du fait des sons émis par le nouveau-né atteint de cette maladie (les symptômes sont nombreux et sont associés à un retard mental prononcé). Il est associé à la délétion de l'extrémité du bras court du chromosome 5 (bandes 5p15.2 et 5p15.3). Cette délétion est donc de très grande ampleur (macrodélétion) et est quasiment un exemple viable unique. Par contre il existe des microdélétions, beaucoup plus difficiles à mettre en évidence, mais aussi beaucoup plus fréquentes. Elles ne sont pas toujours associées à des différences phénotypiques. |
|
On a observé que des microdélétions identifiées et localisées n'ont pas les mêmes conséquences phénotypiques dans toutes les cellules, notamment les cellules sexuelles. Seules les cellules polliniques semblent très sensibles aux délétions (comme aux aneuploïdies). Par contre, les spermatozoïdes, tout comme les ovules des plantes, les ovocytes animaux et les cellules somatiques, pourraient par contre comporter des délétions sans qu'apparaissent des modifications phénotypiques. |
|
duplications |
|
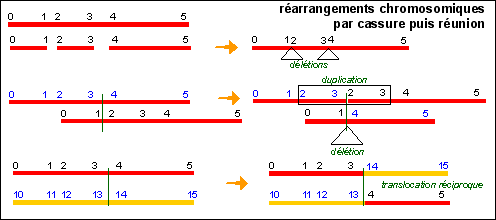
Il n'est pas rare que l'on trouve une copie supplémentaire d'une région chromosomique dans le génome des organismes. Chez les diploïdes la région concernée est effectivement en double exemplaire (on parle de duplication en tandem lorsque les deux zones sont voisines, et de duplication par insertion dans le cas contraire). Chez les diploïdes, en supposant que la duplication intervient après la phase de synthèse de l'ADN et avant la phase de condensation -donc en phase G2 - on peut considérer que la zone dupliquée se trouve en 3 exemplaires dans le génome. |
|
|
||
|
d'après Analyse génétique moderne, De Boeck, 2001 |
|
... en effet, toutes ces interprétations théoriques s'inscrivent dans une théorie de l'information génétique linéaire, associée à une molécule d'ADN unique de très grande taille dans chaque chromosome. La fusion, pour ne pas dire la confusion, entre les mécanismes moléculaires proposés pour les Procaryotes et la très hypothétique structure linéaire du chromosome des eucaryotes, rend le travail de l'historien des sciences ou du philosophe extrêmement compliqué, voire impossible. |
||||
| |
|
|
|
Les variations caryotypiques comme mécanisme évolutif |
||
|
Origine de l'homme, Jean Chaline, Encyclopedia Universalis v12, 2007 |
|
Jean Chaline et ses coauteurs (J. Chaline, B. Dutrillaux, J. Couturier et al., Un modèle chromosomique et paléobiogéographique d'évolution des primates supérieurs, Géobios, Lyon, 1991) se sont efforcés de reconstituer les caryotypes probables des Primates de la lignée des hominidés en comparent les caryotypes d'organismes actuels et en faisant des hypothèses concernant les modifications possibles (duplications, insertions, délétions, fusions...). Ils ont ainsi proposé une reconstitution phylogénétique de cette lignée à partir de ce qu'on peut appeler les distances chromosomiques (voir évolution de l'homme). |
|
|
||
|
2 - Discussion |
||||||
| |
|
2.1 un article fondateur peu convaincant: Luria et Delbrück, 1943 |
||||
|
|
|
|
|
On lira avec profit un chapeau à la publication de l'ESP écrit par Robert J. Robbins en 2001 qui tente de montrer l'importance de cette expérience replacée dans une perspective évolutionniste (mutationnisme, supposé darwinien, contre adaptationisme, supposé lamarckien). Cette interprétation est à mon avis discutable (dans le sens où l'on cherche à nouveau des pères fondateurs au mouvement darwinien). Mais je pense que ce commentaire manque surtout l'essentiel de l'article qui porte sur la méthode de sélection supposée pertinente pour connaître la physiologie de ces micro-organismes (voir ci-dessous Nissim Amzallag). En 1943 la théorie de l'information génétique est déjà dans certains esprits, au moins aux Etats-Unis (voir cours de 1èreS partie historique). Les expériences de Griffith (voir page spéciale) ont ouvert la voie à la recherche d'un principe transformant chimique pour les bactéries. Cette expérience est, à mon avis, bien davantage la preuve de l'importance de la vision chimique de la vie que d'un darwinisme. |
||
| |
|
|
Voici quelques pistes d'interprétations plus ouvertes, sortant du cadre de la théorie de l'information génétique. Dans cette conception ancienne (voir historique dans le cours de 1èreS), mais qui persiste dans l'imaginaire de beaucoup, le matériel génétique d'une bactérie (sélectionnée et repiquée) est absolument identique à celui de ses descendants. Ceci est actuellement beaucoup moins sûr : - chaque bactérie est plutôt une cellule d'un organisme pluricellulaire (voir Shapiro dans le cours de seconde) et possède une grande quantité d'ADN (environ 6 exemplaires de son matériel génétique); donc, les phases de multiplication cellulaire (et non de reproduction) de l'individu peuvent être considérées comme des phases critiques où peuvent se mettre en place des métabolismes nouveaux (voir ci-après). - le métabolisme repose sur un réseau génétique dynamique et fluctuant avec des redondances, l'activité de certains gènes peut être relayée par d'autres en cas de déficience.... une cellule présentant une résistance n'implique pas que les autres cellules n'auraient pas pu présenter cette résistance, plus progressivement, par adaptation à des conditions changées progressivement (voir suite de cette page). |
|||
| |
|
|
|
Analyse de Nissim Amzallag Voici un extrait de l'homme végétal qui souligne les enjeux de l'extraordinaire retentissement de cet article qui influe encore aujourd'hui la conception de la variabilité du vivant pour de nombreux biologistes. J'ai ajouté quelques intertitres personnels. |
||
|
L 'homme végétal, Nissim Amzallag, Albin Michel, 2003, pp 58-62 -Ch 2 : Les fabuleux pouvoirs de l'horloger aveugle
|
|
Contexte
historique (... suite ---->) |
|
«La
grande
innovation de
Luria et
Delbruck fut
la mise au
point d'une
expérience
permettant de
vérifier
si de rares "innovations
avantageuses"
étaient
susceptibles
d'apparaître
au hasard des
mutations.
L'enjeu
était
de taille, et
le
résultat
fut à
la hauteur de
leurs
prétentions:
l'expérience
en question
devint la
démonstration
historique de
l'origine
fortuite des
mutations
avantageuses. |
||
| |
|
2.2 pour sortir de la vision chimique de la vie |
||||
|
Cours de 1ère S: A 1 - De la genèse à l'abandon d'une vision chimique de la vie |
|
Je me suis déjà efforcé autre part de documenter l'histoire de la vision chimique de la vie qui en est venue, jusqu'à la fin du XXème siècle, à orienter tout l'effort de recherche en science de la vie dans une direction qualifiée de moléculariste, allant de pair avec un réductionnisme physico-chimique. |
|
Des changements importants sont apparus depuis et la vision la plus courante actuellement fait cohabiter un déterminisme avec un indéterminisme (voir la science), mettant en avant un principe d'émergence que l'on appelle hasard mais qui n'est plus la hasard salvateur incriminé par Nissim Amzallag (les 4 causes et page de biologie théorique). |
|
Par contre, dans le domaine de l'évolution, et notamment de la phylogénétique moléculaire, la théorie est restée la même et le mutationnisme mécaniciste semble être la règle de compréhension de la variabilité. Il n'est donc pas inutile de revenir sur cet indéterminisme que Nissim Amzallag qualifie d'autonomie. |
| |
|
|
|
adaptations versus génétique/épigénétique |
||
|
L 'homme végétal, Nissim Amzallag, Albin Michel, 2003 |
|
- l'exposition d'une population à des substances toxiques conduit l'ensemble de la population (et non quelques individus préadaptés) à une tolérance progressive (et variable selon les individus). |
|
Viktor Jollos, 1930 - population de paramécies, tolérance à l'arsenic, à l'acétate de plomb notamment |
||
|
|
- des cellules de plantes clonées présentent une grande variabilité dans leur sensibilité aux hormones végétales; ce phénomène, appelé habituation, n'a rien à voir avec une sélection. Pour assurer la croissance d'un cal, les cellules de plantes isolées ne se divisent pas, en effet, spontanément, il est nécessaire de les stimuler par des hormones (auxines, cytokines...). Leur réponse à ces hormones, qu'elles sécrètent elles-mêmes, est complexe. Les résultats obtenus avec les outils de la biologie moléculaire, utilisés pour comprendre cette complexité, n'ont fait que l'augmenter (action sur les gènes des hormones, action épigénétique (notamment par les ARN), action directe sur les protéines actives dans la division...). On a ainsi perdu de vue l'autonomie de chaque cellule, qui est une donnée, qui s'est noyée dans un fouillis de réseaux d'interactions plus ou moins déterministes. |
|
F. Meins Jr, 1989, Habituation : heritable variation in the requirement of cultured plant cells for hormones, Annual Review of Genetics, 23, pp 395-408 (article de synthèse) |
|||
| |
|
-
dans les
années
1980-1990,
pour cadrer
avec
l'hypothèse
d'une
information
génétique
déterministe
on a
considéré
que les
phénomènes
d'adaptation
réversibles
(qui pouvaient
disparaître
- si
l'organisme
était
ramené
dans les
conditions
initiales ou,
même de
façon
spontanée,
sans
changement des
conditions de
milieu) -
seraient
considérés
comme épigénétiques
, parce que
réversibles.
Étant
sous-entendu
que les
phénomènes
génétiques,
comme les mutations
mis en
évidence
par
sélection,
étaient
irréversibles. |
|
|||
|
des travaux allant dans une direction opposée à celle de Luria |
|
travaux
de Hinshelwood - Colony Formation by Bact. lactis aerogenes on Solid Media Containing Antibacterial Agents, A. C. R. Dean and Cyril Hinshelwood, Proceedings of the Royal Society of London. Series B, Biological Sciences, Vol. 140, No. 900 (Nov. 20, 1952), pp. 339-352 (abstract)
|
|
- The Development of Drug Resistance in Strains of Saccharomyces cerevisiae: Resistance to 2.4-Dinitrophenol and to Brilliant Green, G. Wild and Cyril Hinshelwood, Proceedings of the Royal Society of London. Series B, Biological Sciences, Vol. 144, No. 916 (Nov. 29, 1955), pp. 287-297 (abstract):«En considérant la différence de comportement entre les souches habituées et les non-habituées, nous suggérons que le développement progressif de l'habituation des cellules de levure à ces conditions, corresponde davantage à une réponse adaptative à l'inhibiteur qu'à la sélection de mutants pré-existants. (On the basis of the time relations for the trained and the untrained strains it is suggested that the gradual development of training of the yeast cells under these conditions is more likely to be an adaptive response to the inhibitor than the selection of pre-formed mutants).» |
||
|
L 'homme végétal, Nissim Amzallag, Albin Michel, 2003, p 71-75 |
|
|
|
émergence d'une nouvelle fonction chez Escherichia coli |
||
| |
|
Hall et Hartl, dans les années 1975, utilisèrent des bactéries "mutées" dans lesquelles un segment entier du génome avait été éliminé par sélection (cette portion fonctionnelle, contenant plusieurs gènes, était ce que l'on connaissait sous le nom d'opéron lactose). La souche "mutée" était donc incapable de se nourrir de lactose. Quelques jours après la mise culture de cette souche sur un milieu dont le lactose devenait progressivement la seule source de nutriment, ils virent apparaître des individus ayant récupéré la totalité de la fonction (d'utilisation du lactose). Ils notèrent que cette nouvelle fonction était due à l'activité d'une nouvelle enzyme capable d'utiliser le lactose, et adaptée à la concentration en lactose du milieu. En fait, cette enzyme était déjà présente dans la souche initiale, mais présentait alors une faible activité. Il s'agit clairement d'un cas de redondance métabolique, qui invalide la méthode de sélection des mutants. Mais il ne faut pas croire que ces résultats firent perdre pied aux tenants du mutationnisme salvateur. On inventa un mécanisme d'erreurs d'autant plus nombreuses dans les gènes que ceux-ci étaient exprimés, doublé d'un processus de duplication qui permettait d'envisager une compétition entre gènes pour retomber sur le hasard sélectif.... ces explications ont toujours cours, malgré la simplicité de l'hypothèse de la redondance. |
|
travaux de B.G. Hall des années 1975-1985 (travaux de cet auteur dans Pubmed dont un bon nombre sont en libre accès |
||
| |
|
|
|
phénocopies |
||
|
Une phénocopie est un phénotype considéré d'abord comme modification environnementale (non héréditaire) mais qui mime un phénotype obtenu par mutation d'origine génétique. |
|
Le terme de phénocopie est souvent mal employé et mal compris. Étymologiquement le mot est composé des racines phéno = visible (du grec phainô= paraître) et copie (du latin co-opia = abondance). |
|
Le summum de l'inadéquation de l'emploi du mot est celui que l'on trouve dans le dictionnaire de l'Encyclopedia Universalis : « en biologie, individu sur lequel on a provoqué des mutations génétiques». |
||
|
|
Un des premiers à intégrer le terme dans sa biologie théorique fût probablement Waddington (j'ai présenté par ailleurs des éléments que René Thom lui a emprunté, voir aussi le cours de seconde sur l'effort). Voilà une description de ses intéressantes thèses par Saunders. |
|
||||
|
L'évolution des formes biologiques, Peter.T. Saunders, Intellectica, 1993/1, 16, pp. 61-83 (http://www. intellectica. org/archives/ n16/16_06_ Saunders.pdf) |
|
« Ainsi, puisque le processus de développement est non-linéaire, il est à la fois stable et capable de changements organisés importants. Le biologiste Conrad Waddington a illustré cette idée par son fameux "paysage épigénétique" (figure 1) : le développement d'un organisme y est représenté par une bille qui descend une vallée représentant un chemin ontogénique (Waddington 1957 ; Saunders 1992). Les gènes contrôlent la forme du paysage, et une mutation peut être vue comme une modification de ce paysage. La mutation n'aura un effet perceptible que si la modification est suffisante pour détourner la bille vers une autre vallée. Puisque ce qui compte est le choix de la vallée, autrement dit du chemin, différentes mutations peuvent avoir le même effet sur le développement bien que les changements de paysages seraient probablement un peu différents pour chacune. Une perturbation extérieure sur le système aura un effet similaire à celui d'une mutation : peu ou pas d'influence si la bille reste dans la même vallée, mais la possibilité d'un développement entièrement différent si elle passe au-dessus d'une "ligne de partage des eaux". Nous savons aujourd'hui que ces propriétés sont caractéristiques des systèmes non-linéaires, mais Waddington a abouti à cette vision des choses empiriquement, à partir de ce qu'il savait des propriétés des systèmes en cours de développement. Un exemple particulièrement clair du type de comportement qu'il avait à l'esprit est le phénomène bien connu de la phénocopie. Perturber le développement d'un embryon génétiquement normal peut avoir le même résultat qu'une mutation. Ainsi, un embryon de drosophile normal traité avec de la vapeur d'éther peut produire une mouche adulte avec une paire d'ailes supplémentaire, exactement comme le mutant bithorax. Cet exemple n'est pas isolé. Dans les années quarante, Richard Goldschmidt réussit à obtenir une phénocopie de tous les mutants connus de la Drosophile. Les phénocopies de papillons mutants sont suffisamment courantes pour que les lépidoptéristes se sentent obligés de les exclure explicitement de leurs collections. La phénocopie montre clairement l'importance réduite accordée aux gènes dans la conception de l'évolution qui émerge actuellement. La source de variations n'est plus la mutation, mais l'ensemble des potentialités de l'organisme, les mutations ou les perturbations de l'environnement ne jouant que le rôle d'un stimulus. Cette idée a quelques conséquences importantes. Si le facteur crucial réside dans une mutation particulière, il est peu probable que la même variation apparaisse chez plusieurs individus en même temps. Et si tel est le cas, alors à moins qu'elle ne procure un avantage sélectif vraiment important elle disparaîtra probablement par "dérive génétique", autrement dit selon le hasard. Si, au contraire, la variation correspond à un changement vers un chemin déjà existant dans le paysage épigénétique, lequel est commun à tous les individus, il est bien plus probable qu'elle ait lieu dans suffisamment d'individus pour subsister.» |
|
Notes: |
||
|
|
Remarque: |
|||||
| |
|
Piaget revendique une acception du terme personnelle dans son effort de redonner au vivant un certaine autonomie (détails dans le chapitre III (Les phénocopies, p7) d'un article de Susan OYAMA : Penser l'évolution : l'intégration du contexte dans l'étude de la phylogenèse, de l'ontogenèse et de la cognition, Intellectica, 1993/1, 16, pp. 133-150). L'auteur critique cette utilisation et donne une définition exacte du terme (p 10): « La signification réelle de la phénocopie réside dans le fait que le même phénotype peut provenir d'une modification soit génétique,soit environnementale». |
|
Antoine Danchin (voir son site à l'Institut Pasteur), connu pour sa participation au débat de biologie théorique (ses échanges avec René Thom dans le CD-ROM des œuvres complètes de René Thom sont toujours courtois, même si je suis loin d'être d'accord avec sa vision de la biologie) a écrit, il y a déjà de nombreuses années une Note critique sur l'emploi du terme phénocopie, In: "Théories du langage, théories de l'apprentissage" (CRSH), Le Débat Chomsky-Piaget, Le Seuil (1979) pp 109-114 (traduit en anglais en 1981). |
|
|
| |
|
2.3 la méthode sélective en accusation |
||||
| |
|
Au niveau phénotypique et non plus génotypique (théorique donc) qu'est-ce qui permet d'affirmer que l'on a affaire à une mutation ? |
|
La démarche est une démarche analytique : dans une population homogène, on isole un ou des individus qui présentent un trait de caractère rare (qu'il corresponde à une perte ou à un gain de fonction). Ces individus sont isolés, et sont considérés comme mutants s'ils conservent ce caractère sur plusieurs générations. Dans certains cas simples, notamment chez les procaryotes on associe le phénotype muté à un génotype muté grâce à un séquençage très partiel du génome (au niveau de gènes que l'on suppose impliqués). |
|
On voit que dans cette méthode l'interprétation génétique reste en toile de fond (c'est une hypothèse explicative énoncée dès le départ), et ne peut donc pas être prouvée par l'expérience, qui consiste en une simple sélection. On peut en effet imaginer que les "mutants" sont des organismes capables de s'adapter et que les variations génotypiques sont présentes chez de nombreux autres individus non testés, car ne présentant pas le changement de caractère, ou un changement moindre. |
|
la sélection remplace la compréhension |
|
Nissim Amzallag pointe de façon très claire l'erreur que l'on fait dans le raisonnement lorsque l'on cherche à prouver la présence de mutation dans le cadre du paradigme moléculaire. La
raison
malmenée |
|
«
La
sélection
a
posteriori d'individus
modifiés
n'est pas
seulement la
méthode
exclusive en
biotechnologie,
mais
également
la voie
d'investigation
privilégiée
du vivant.
C'est par
l'analyse de
mutants,
individus
reconnus comme
déficients
pour une
fonction, que
les
biologistes se
proposent de
comprendre le
fonctionnement
d'un organisme
normal. Il est
possible, au
moyen
d'irradiations
ou d'autres
techniques,
d'altérer
la structure
ou
l'expression
de petites
régions
du
génome.
L'analyse des
modifications
observées
en
parallèle
sur
l'organisme et
sur les
gènes
affectés
permet
d'établir
un lien de
causalité
entre
gènes
et fonctions,
génotype
et
phénotype.
Le cas
idéal
est bien
entendu celui
où il
est possible
d'établir
un lien direct
entre une
déficience
fonctionnelle
et
l'altération
d'un seul
gène,
comme c'est le
cas dans de
nombreuses
maladies dites
génétiques.
C'est la
première
mise en
évidence
d'une telle
relation qui
valut, en
1958, le prix
Nobel à
Georges
Beadle et
Edward Tatum.
Depuis lors,
cette approche
est devenue la
méthode
privilégiée
d'investigation,
celle qui
ouvrit la voie
à la
biologie
moléculaire
et à la
correspondance
étroite
entre science
et technologie
dans le
domaine du
vivant. |
||
| |
|
|
||||
|
Une approche du syllogisme, formalisation du raisonnement par Aristote |
|
Tout
être
vivant est
mortel (P1) moyen terme= être vivant, sujet en P1 puis prédicat en P2 (1ère figure) P1(majeure) affirmative universelle (mode A) P2 (mineure) affirmative particulière (mode I) C conclusion affirmative particulière (I) Ce syllogisme s'appelle AII ou Darii, selon la terminologie aristotélicienne. |
|
Le
syllogisme
est
basé
sur deux
propositions (prémisses,
P1 et P2)
dont on
déduit
une
conclusion (C).
L'idée
géniale
et novatrice
d'Aristote est
de s'appuyer
sur le
structuration
du
raisonnement
et non pas sur
le contenu
(c'est
d'ailleurs
pourquoi de
nos jours la
science du
syllogisme est
incluse dans
la logique
formelle). Le
côté
séduisant
du syllogisme
vient de ce
que le rapport
entre la
conclusion et
chacune des
prémisses
est
caché
dans le terme
moyen,
commun aux
deux
prémisses,
la
première
étant
qualifiée
de majeure
et la seconde
de mineure.
Les variantes
du syllogisme
viennent du
type
(quantité)
de proposition
(universelle
ou
particulière),
de leur
qualité
(affirmative
ou
négative)
ou encore de
la position du
moyen terme
dans les
prémisses. |
||
| |
|
Dans notre petit "jeu", si nous appliquons ce raisonnement à la méthode sélective, nous avons la conclusion: (C) Nous avons sélectionné une bactérie résistante, parce que mutée (sous-entendu au niveau de son génome, ce qui cause son phénotype particulier). Efforçons nous de restituer les propositions cachées du raisonnement. Quelques propositions sur le mode IAI (en inversant la prémisse universelle et la prémisse particulière): (P1) Cette bactérie apparaît comme résistante. Comme (P2) toutes les modifications de phénotypes (et donc les résistances) sont dues à des mutations, (C) cette bactérie est donc mutée. (P1) Cette bactérie survit. (P2) Ne peuvent survivre que les individus adaptés. (C) Cette bactérie est préadaptée. (P1) Cette bactérie peut survivre avec un nutriment original. (P2) La nutrition nécessite des gènes adaptés pour chaque type de nutriment. (C) Cette bactérie possède donc des gènes originaux qui lui permettent d'utiliser ce nutriment original. |
|
Il
est clair
qu'à
chaque fois
c'est la
prémisse
universelle
qui est fausse
dans sa
généralité
: |
||
| |
|
Il ne faut pas confondre les syllogismes non concluants (qui ne permettent pas d'énoncer de proposition) et les faux syllogismes ou encore sophismes (volontairement trompeurs). |
|
un
sophisme bien
connu
d'Anaxagore,
qui caricature
le
syllogisme : |
|
En n'énonçant pas les prémisses universelles de leurs raisonnements, les biologistes qui se meuvent au sein d'un paradigme font œuvre de sophistes. |
| |
|
|
||||
| |
|
Il est donc à craindre que l'on ai indûment construit un échafaudage imaginaire en biologie moléculaire sur la stabilité de l'information génétique et sur sa variation assimilée aux mutations. |
|
Construire de nouveaux modèles à partir d'un autre regard sur la variation est indispensable, mais demande un travail, qui en plus d'être franchement à contre-courant, s'avère être extrêmement difficile, car nécessitant de construire les outils avant de pouvoir obtenir des résultats. Nissim Amzallag s'est attelé à cette tâche... je vais m'efforcer de présenter quelques aspects de son travail dans la quatrième partie de cette page. |
||
| |
|
2.4 - des statistiques assourdissantes |
||||
|
|
Dans
la
démarche
scientifique
qui
établit
des modèles
plutôt
que des lois
(voir
René
Thom, une théorie
générale
des
modèles),
les
statistiques
interviennent
soit dans
l'élaboration
du
modèle
(statistiques
descriptives
comme les
nomme N.
Amzallag) soit
dans la
justification
(a
posteriori)
du
modèle
(statistiques
qualifiées
de justificatives). |
|
*La Raison malmenée. De l'origine des idées reçues en biologie moderne, Gérard Nissim Amzallag, Préface d'André Pichot, CNRS Editions 2002, p 194-196 (Les statistiques justificatives) |
|||
| |
|
|
|
les analyses de variance (ANOVA) |
||
|
RM*p195 |
|
Il est d'usage de décomposer les caractères dans les études de populations en trois parties: le génotype (supposé constant et héritable, s'exprimant quelles que soient les conditions de milieu), l'environnement (la part du phénotype sous la dépendance directe du milieu) et la composante d'interaction entre ces deux premiers termes. |
|
Les
analyses de
variance (ou
ANOVA)
permettent, en
comparant des
populations
exposées
à des
environnements
différents
de
déterminer
STATISTIQUEMENT
et
quantitativement
la part
respective des
trois
composantes. Ainsi, dans le modèle de contrôle génétique du développement, majoritairement considéré, on exerce une «pression sur le réel» de type «autocatalytique, en renforçant toujours davantage la réponse positive, parce que celle-ci fait déjà partie intégrante de la méthode d'investigation». |
|
Amzallag relie ce parti-pris à la comparaison que fait Lewontin: «Par exemple, si deux hommes construisent un mûr en posant des briques l'une sur l'autre, nous pouvons aisément quantifier leur contribution respective en comptant le nombre de briques posées par chacun. Par contre, si l'un prépare le ciment et l'autre pose les briques, il est absurde de mesurer leur contribution respective en comparant le nombre de briques posées avec le volume de ciment produit» (R.C. Lewontin, The analysis of variance and the analysis of causes, American Journal of Human Genetics, 1974, 26, 400-411 - cité dans RM p 196). |
| |
|
|
|
du bruit à la surdité |
||
|
RM*p200s |
|
Une irréductible variabilité est inhérente à toute mesure expérimentale. Vis-à-vis de la valeur absolue, c'est l'incertitude. Vis-à-vis du sens de la mesure les statistiques s'efforcent de chiffrer la fiabilité de la mesure vis-à-vis d'un phénomène, soit approximé à partir d'un traitement statistique des données (régression par exemple), soit déduit à partir des propriétés du système (fonction continue par exemple dans un modèle thomien).
|
|
Poussé à l'extrême le raisonnement déterministe statistique considère que la variabilité est non orientée (isotrope) dans sa cause comme dans son effet. La variation est alors évaluée à l'aide de l'écart-type qui représente l'amplitude des variations par rapport à la valeur moyenne. On définit alors une limite arbitraire à partir de laquelle on considère que les variations autour de la moyenne sont plus importantes que le bruit de fond inhérent à l'expérience. Une mesure très excentrique a donc toutes les chances d'être considérée comme un artefact expérimental. Et le chercheur est fondé à recommencer son expérience jusqu'à ce qu'il obtienne une majorité de résultats qui s'inscrivent dans les limites de la loi statistique choisie, avec une variabilité considérée comme acceptable.
° la causalité redondante est définie par N. Amzallag par opposition à la causalité simple (existence d'une voie unique de régulation du paramètre étudié; par exemple un substrat, une enzyme, un produit), comme un cas où le paramètre étudié est influencé par au moins deux voies redondantes de régulations (par exemple, un produit avec deux substrats et deux enzymes). |
|
Dans un travail de biologie moléculaire, avec l'arsenal technique (et donc le coût) que représente une publication - par exemple pour les résultats de l'expression d'un gène ciblé -, on considère que l'obtention d'un seul résultat significatif est probante, sans même exiger une reproductibilité de l'expérience, impossible à réaliser d'ailleurs la plupart du temps étant donné la destruction du matériel vivant aux fins d'analyse. En recherchant des résultats conformes (paradigmatiquement corrects) on se ferme à une complexité considérée comme inaccessible, par exemple celle reposant sur des cas de redondance des réseaux de régulation: ce qu'Amzallag nomme la causalité redondante°.
|
| |
|
|
|
|
||
|
|
|
«...Il est impossible de déceler, au moyen de la méthode d'investigation traditionnelle, si l'effet observé sur la moyenne est un artefact (c'est-à-dire une transformation artificielle d'une causalité redondante° en causalité dominante) ou bien s'il reflète une réalité biologique, puis que la méthode classique d'investigation ne permet d'étudier un système qu'après avoir transformé ses régulations selon un mode de causalité dominante». |
|
«... les mutants ne sont détectés que grâce aux différences qu'ils affichent par rapport aux individus normaux, c'est-à-dire par le résultat même de la [supposée] mutation. Par voie de conséquence, les mutations détectables sont uniquement celles relatives à une causalité dominante. Dans le cas d'une causalité redondante d'un réseau fonctionnel, les mutants sont décelés uniquement si la mutation affecte un tronçon en chaîne du réseau, d'un nœud ou dans les dernières étapes (parfois linéaires) de la production d'un composé ou de la régulation d'une variable. Dans les autres cas, la déficience d'un gène passera complètement inaperçue, même si le produit du gène en question est impliqué dans le processus étudié». |
|
On
ne pourra
dépasser
ces limites
qu'en
élargissant
le cadre
épistémologique
afin
d'intégrer
l'hypothèse
déterministe
et la
causalité
redondante°,
tout en
trouvant le
moyen de
distinguer
entre ces deux
modes de
régulation. |


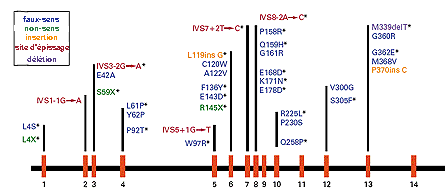



|
|
|
|
|
|
|
|
||
|
3. Comprendre la variation en biologie ... une question ouverte |
||||||||
|
sans prétendre balayer la question, voici quelques approches... |
|
3.1 l'approche fonctionnelle mathématique |
||||||
|
|
|
C'est Leibniz qui, dans un manuscrit de 1684, utilise le mot fonction dans le sens où on le comprend actuellement (ou presque*): « toute quantité qui varierait d'un point à un autre d'une courbe » . On considère qu'on lui doit les mots de variable et constante ainsi que de paramètre. |
|
En mathématique une variable est à la fois une grandeur (mesurable) et un symbole formel. En biologie paramètre et variable sont employés dans le même sens d'élément mesurable ou contrôlable d'un système (voir cours de seconde sur les paramètres de l'effort). |
|
*d'après M. Kline, Mathematical Thought from Ancient to Modern Times, New York, Oxford University Press, 1972, p. 339-340.; cité dans les notes de Paraboles et catastrophes, René Thom |
||
| |
|
|
|
|
||||
|
|
Ainsi, on pourrait ramener le problème de la variabilité à celui de la description du phénomène vivant à l'aide de fonctions et donc de constantes et de variables. |
|
Dans cette approche le phénomène vivant est variation et il n'y a pas de méthode particulière pour étudier ou exprimer la variation qui est l'expression même de la vie. La compréhension de la variation dépend alors principalement de l'échelle à laquelle on se place. |
|
Mais il existe une autre approche, plus philosophique, qui repose davantage sur les temps du vivant. |
|||
| |
|
3.2 l'approche par le changement |
||||||
| |
|
|
|
revenir à Aristote, puissance et acte.... |
||||
|
le secours d'Aristote |
|
Aristote est considéré comme le premier philosophe grec qui a résolu le problème du mouvement / changement à l'aide de la division de l'être en acte et puissance. L'unité vient de l'acte, la multiplicité de la puissance. Un être vivant est à la fois en acte, un être, unique dans son être corporel, et à la fois multiple, divisé, car matériel. |
|
|||||
| |
|
|
|
la variation comme différence et la variation comme changement |
||||
|
étymologie |
|
Une
fois encore
revenir
à
l'étymologie
permet de
débusquer
des a
priori :
variation
vient du latin
varius
qui a deux
sens que l'on
retrouve dans
l'ancien
français
vair ou
vairon (de
couleur
différente
mais aussi
changeante,
par exemple
pour les
yeux): |
|
En
biologie la
variation peut
donc
désigner Une cellule et, a fortiori, un organisme sont des ensembles hétérogènes. Et l'on peut aussi bien les étudier comme formes stables que comme formes changeantes. |
|
Cette oscillation continuelle entre ces deux sens brouille notre perception de la variation que l'on croît saisir comme le mouvement d'une forme vivante stable (la vie étant considérée comme un processus dynamique, mais stable) ou comme une propriété intrinsèque des organismes vivants que l'on sait en perpétuelle évolution. |
||
|
|
|
la variation comme différence |
|
|
|
|
||
|
|
La catégorie qui représente le mieux cette stabilité est l'espèce. Une espèce biologique est la forme à laquelle appartient un être vivant de par sa naissance à partir d'un autre être vivant. |
|
Les êtres vivant (individus) naissent, vivent et meurent au sein de l'espèce. L'espèce représente le continu et les individus des formes saillantes*. La différence entre les individus au sein de l'espèce c'est la variation intraspécifique. |
|
* pour le vocabulaire de René Thom voir la page sur les 4 causes d'Aristote en SVT |
|||
|
|
|
la variation comme changement |
|
|
|
|
||
|
|
Les espèces évoluent. L'évolution c'est l'idée selon laquelle les espèces vivantes se transforment et dérivent les unes des autres. |
|
Les individus naissent, croissent, vivent, meurent. Les individus changent tout d'abord au cours de leur vie (morphogénèse, croissance, sénescence...) et ensuite en fonction de leur environnement (adaptation...). |
|
Même si c'est l'espèce, qui est le sujet théorique de l'évolution, seuls les individus sont accessibles expérimentalement. La fonction de l'individu qui le fait participer de l'espèce est la reproduction, c'est donc probablement là que se joue l'évolution au niveau local. |
|||
|
|
|
L'espèce
est un
continu. Mais
il y a une discontinuité,
lors de la spéciation,
apparition
d'une nouvelle
espèce,
et lors de l'extinction. Contrairement à ce que l'on pourrait penser, l'étude des populations nous permet d'atteindre la stabilité de l'espèce, par l'étude des variations intraspécifiques, alors que l'étude de phylums nous permet de comprendre les mécanismes de la spéciation. Vouloir confondre ces deux ordres de grandeur est une tentation extrêmement prégnante mais que je crois qu'elle conduit à des erreurs. |
|
En
comparant les
individus au
sein d'une
espèce
on peut
évaluer
la variation
intraspécifique,
À UN
INSTANT
DONNÉ,
mais non la
variation AU
COURS DU
TEMPS, pour
laquelle il
faut
accéder
à des
individus
à des
époques
différentes,
ce qui ne peut
se faire que
par la
paléontologie,
qui n'est pas
une science
expérimentale. |
|
La variation interspécifique ne peut guère se faire qu'en comparant des espèces et donc des individus d'espèces voisines. |
||
| |
|
|
|
|||||
| |
|
|
La stabilité de l'espèce et donc les petites variations intraspécifiques s'étudient à l'échelle de la population pour des temps de l'ordre de quelques générations (temps de reproduction). C'est, de loin, le sujet le plus fréquent des études sur la variation. Ces études concernent donc davantage la stabilité de l'espèce, malgré les variations individuelles, que l'évolution. |
|
L'évolution, dans son approche interspécifique, s'étudie à l'échelle des phylums (embranchements et autre regroupement classificatoire évolutif). La méthode la plus universellement employée est la méthode cladistique. Cependant, cette méthode n'inclue aucune théorie de la variation spécifique, elle est, en cela, inachevée. La plupart du temps les chercheurs considèrent que cette variation résulte d'un mutationnisme (moléculaire au départ) généralisé aux fonctions. Nous nous sommes efforcés de démonter cette interprétation simpliste ci-dessus. |
|||
| |
|
|
||||||
|
Remarque |
|
Mes
propos
pourraient
être mal
interprétés.
Je ne dis pas
que la
variation des
individus est
un bruit
indésirable,
comme le
fustige si
bien Nissim
Amzallag,
mais, bien au
contraire, que
la variation
individuelle,
qui est
vraiment une
variation de
l'individu,
n'est,
profondément,
que
l'expression
de la
stabilité
de
l'espèce. |
|
J'appelle de tous mes vœux, comme je vais essayer de le présenter plus bas, et comme pédagogue ouvert à toutes les inventions de l'esprit humain qui cherche à comprendre la vie, un développement de la conception d'une individualité émergente qui supporte un mécanisme original de spéciation qui pourrait être élargi au sein d'une nouvelle théorie évolutive (la dissociation autonome pour Nissim Amzallag). |
|
N. Amzallag, Du sens de la variabilité, 2007, in Génétiquement indéterminé, ch 1 (pp27-58), Quæ (Cemagref, Cirad, Ifremer, Inra) (table des matières: http://www.quae.com/livre/ ?GCOI=27380100948540) L'homme végétal : pour une autonomie du vivant, Gérard Nissim AMZALLAG, Albin Michel, 2003 (préface de Bernard Werber) |
||
| |
|
3.3 points de vue |
|
|
||||
|
celui de Charles Darwin dans : The Variation of Animals and Plants under Domestication, 1868
|
|
C. Darwin en observateur minutieux ou conteur intarissable décrit des variations héréditaires de façon très détaillée chez un grand nombre d'espèces domestiques (chats, chiens, chevaux, ânes, porcs, bovins, ovins, caprins, lapins, pigeons (exhaustif !!!), volailles, vers à soie, céréales, plantes potagères, fruitiers, fleurs...). Il se rapporte parfois à sa théorie de la descendance avec modification que l'on retrouve dans L'origine des espèces mais il y a dans cet ouvrage une faconde stupéfiante qui déborde généreusement l'étroitesse des développements théoriques modernes plus ou moins inspirés de cet auteur. |
|
Une
traduction
française
est disponible
gratuitement
sur Gallica: http://gallica.bnf.fr
/ark:/ 12148/
bpt6k406270g |
|
le texte original en anglais (formal html avec chapitrage et figures séparées) en téléchargement gratuit est disponible sur la librairie virtuelle Gutenberg: http://www.gutenberg. org/ etext/ 3332. Pour de plus amples détails sur les différentes éditions voir désormais Darwin online qui offre l'intégralité de l'œuvre de Darwin. |
||
| |
|
|
||||||
|
bergsonisme
téléchargement gratuit des livres d'H. Bergson (très utile pour faire des recherches intratext automatisées) page sur le temps (terminales S) |
|
Bergson, par sa démarche métaphysique originale (à partir de l'intuition) réconcilie le corps et l'esprit dans la durée (Matière et mémoire : essai sur la relation du corps et de l'esprit, 1896, cité MM). Sans développer une ontologie de la matière, qu'il considère impossible, il renvoie dos à dos les positions tranchées matérialistes et idéalistes. Il met ainsi en évidence l'inanité d'une théorie de la variation puisqu'elle est l'expression du jaillissement permanent de nouveauté qu'est la vie. |
|
C'est donc dans l'Évolution créatrice (1907, accès gratuit au livre, cité EC) qu'il faut aller chercher la signification de la variation. Si L'évolution créatrice est très ancrée dans son temps par la discussion des théories (Lamarckisme, Darwinisme, travaux de Spencer, qui furent les premières amours de Bergson) et de philosophies (le ch IV est un excellent résumé de la vision des sciences de la vie de nombre de systèmes philosophiques), cet ouvrage n'apporte pas moins une vision originale, lumineuse et cohérente de l'évolution. Le premier point fondamental est, à mes yeux, que le milieu ne se distingue pas de façon nette de l'organisme vivant, à la manière de ce que Bergson dit du corps et de l'esprit ou de la matière et de la conscience (ci-contre quelques phrases; voir extraits plus complets sur la page sur Bersgon). |
|
Il
ne s'agit de
s'arrêter
ni au
réalisme
naïf, ni
à
l'idéalisme
naïf : «
Les
mêmes
besoins, la
même
puissance
d'agir qui ont
découpé
notre corps
dans la
matière
vont
délimiter
des corps
distincts dans
le milieu qui
nous
environne. S'il n'y a pas de milieu indifférent (on retrouve ces termes chez Canguilhem), on pourrait même dire avec Bergson que le milieu proche n'est séparé de l'être vivant que dans la durée et non pas dans l'espace. Je voudrais aussi faire le rapprochement avec le discours d'André Pichot sur la disjonction d'évolution entre l'être vivant et son milieu. |
||
| |
|
|
|
|||||
| |
|
« Que la condition nécessaire de l'évolution soit l'adaptation au milieu, nous ne le contestons aucunement. Il est trop évident qu'une espèce disparaît quand elle ne se plie pas aux conditions d'existence qui lui sont faites. Mais autre chose est reconnaître que les circonstances extérieures sont des forces avec lesquelles l'évolution doit compter, autre chose soutenir qu'elles sont les causes directrices de l'évolution. Cette dernière thèse est celle du mécanisme. Elle exclut absolument l'hypothèse d'un élan originel, je veux dire d'une poussée intérieure qui porterait la vie, par des formes de plus en plus complexes, à des destinées de plus en plus hautes. Cet élan est pourtant visible, et un simple coup d'œil jeté sur les espèces fossiles nous montre que la vie aurait pu se passer d'évoluer, ou n'évoluer que dans des limites très restreintes, si elle avait pris le parti, beaucoup plus commode pour elle, de s'ankyloser dans ses formes primitives. Certains Foraminifères n'ont pas varié depuis l'époque silurienne. Impassibles témoins des révolutions sans nombre qui ont bouleversé notre planète, les Lingules sont aujourd'hui ce qu'elles étaient aux temps les plus reculés de l'ère paléozoïque. (--->) |
|
La
vérité
est que l'adaptation
explique les
sinuosités
du mouvement
évolutif,
mais non pas
les directions
générales
du mouvement,
encore moins
le mouvement
lui-même. Là est la première erreur du finalisme. Elle en entraîne une autre, plus grave encore. (--->) |
|
Si
la vie
réalise
un plan, elle
devra
manifester une
harmonie plus
haute à
mesure qu'elle
avancera plus
loin. Telle,
la maison
dessine de
mieux en mieux
l'idée
de
l'architecte
tandis que les
pierres
montent sur
les pierres.
Au contraire,
si
l'unité
de la vie est
tout
entière
dans
l'élan
qui la pousse
sur la route
du temps, l'harmonie
n'est pas en
avant, mais-en
arrière.
L'unité
vient d'une
vis a
tergo :
elle est
donnée
au
début
comme une
impulsion,
elle n'est pas
posée
au bout comme
un attrait.
L'élan
se divise de
plus en plus
en se
communiquant.
La vie, au
fur et
à
mesure de son
progrès,
s'éparpille
en
manifestations
qui
devront sans
doute à
la
communauté
de leur
origine
d'être
complémentaires
les unes des
autres sous
certains
aspects, mais
qui n'en
seront pas
moins
antagonistes
et
incompatibles
entre elles. |
||
|
4 . Explorer la variation individuelle par l'expérience |
||||||||
 Nissim Amzallag (Photo trouvée sur le site de Nakim) En travaux *Du sens de la variabilité, Nissim Amzallag, in Génétiquement indéterminé - le vivant auto-organisé, ch1, éditions Quæ (Cemagref, Cirad, Ifremer, Inra), 2007 (partiellement accessible sur http://books.google.com) Rejet de souche (cinquième partie) in La Raison malmenée. De l'origine des idées reçues en biologie moderne, Gérard Nissim Amzallag, Préface d'André Pichot, CNRS Editions 2002, ISBN : 978-2-271-05998-7 (une note de lecture de Girolamo Ramunni) |
|
Si j'éprouve une forte attirance pour la philosophie (je ne cache pas mon attirance pour les propos de Bergson ci-dessus), philosophie qui, je crois, peut faire gagner du temps, je suis cependant admiratif du profond enracinement de Nissim Amzallag dans l'expérience. Ce petit chapitre va s'efforcer de rendre compte, progressivement, des travaux qu'il a lui-même vulgarisés dans La Raison malmenée. De l'origine des idées reçues en biologie moderne (cinquième partie: Rejet de souche) et dans le chapitre1 de Génétiquement indéterminé. J'ai du parfois aller chercher quelques figures dans ses publications scientifiques... |
||||||
|
|
4.1 - Historique |
|||||||
|
|
|
|
La variation n'est pas un bruit |
|||||
|
|
«...il existe un choix qui n'a pour ainsi dire jamais été remis en cause en biologie : celui considérant la variabilité comme l'expression d'un bruit». DSV*p 27 |
|
À
la suite de
Poincaré
nous avons
cherché
à
modéliser
les
phénomènes
par des
fonctions
(voir Maths-SVT).
Il y a
peut-être
eu une erreur
faite avec la
tendance
à
considérer
que ce qui
n'était
pas la
fonction
était
un bruit
(phénomène
= fonction
(loi) +
bruit). |
|||||
|
|
|
|
||||||
|
|
« Un simple aperçu de la réalité biologique révèle l'existence d'au moins deux options épistémologiques. La première (la transposition de l'approche de la physique classique à la biologie) s'appuie sur le postulat de validité de la condensation des données au travers de la moyenne. Dans ce cas, l'écart-type représente la quantité de bruit dans l'expression de la valeur moyenne. Au contraire, la variabilité n'est pas un simple bruit isotrope dans la seconde approche. Elle est l'expression d'une individualité de la réponse». DSV* p 30 |
|
Travaux de Pascale Mentré sur l'eau (voir Au-delà de la cellule). Comme les éléments du vivant (molécules, individus...) sont singuliers, les réponses sont singulières (individuelles). |
|||||
|
|
La biométrie (qui utilise l'outil statistique) à poussé les expérimentateurs à travailler de plus en plus sur des populations homogènes, avec des comportements prédictibles... ce qui a éliminé la variation et a renforcé la légitimité de l'outil statistique (voir ci-dessus). |
|
||||||
|
Ce passage est géant. Il invalide la relation de cause à effet de nombre de résultats expérimentaux puisqu'on a normalisé les résultats et excluant toute individualité. |
|
«
En toute
rigueur,
l'ignorance de
la nature de
la
distribution
des valeurs
mesurées
dans les
populations
devrait
conduire
à
l'usage (...)
de
statistiques
(...) dites non-paramètriques.
Cependant,
fondés
sur une
hiérarchisation
qualitative
des individus
selon un
critère
donné,
ces outils
statistiques
n'utilisent
qu'une faible
partie de
l'information
contenue dans
les
données,
si bien que
leur usage
aboutit
à une
généralisation
de
portée
très
limitée.
Elle
réduit
ainsi le
"rendement" de
la machine
scientifique.
C'est
pourquoi, en
biologie, la
condition de
normalité
est non pas
vérifiée
par
l'expérimentation,
mais
pratiquement
tout le temps
implicitement
entendue. |
|
Pour un comportement d'un ensemble de molécules, c'est donc une fonction globale statistique que l'on cherche et non le comportement individuel qui lui repose sur la cause-effet (local). Mais la fonction globale n'est pas justifiée par des entités individuelles (les molécules) mais par le tout (organisme). C'est pour cela qu'il applique son raisonnement aux populations et non pas aux molécules. On peut parler d'individualité à partir des cellules (et encore...). |
||||
| |
|
|
|
de la difficulté de publier des résultats sur l'individualité |
||||
|
2 exemples |
|
« Et pourtant, la variabilité demeure une réalité incontournable en biologie. En témoigne, par exemple, la variation inter-individuelle de l'ordre de 1000 fois qui est mesurée dans la sécrétion de l'endomannanase par l'extrémité de jeunes racines de tomates pourtant génétiquement identiques et exposées aux mêmes conditions (15). Un tel écart rend impossible l'usage de moyennes, du moins tant que l'hypothèse de normalité n'est pas vérifiée. Mais une telle vérification n'est généralement même pas envisagée pour deux raisons. Tout d'abord, elle exigerait d'effectuer la mesure sur plusieurs centaines d'individus, ce qui représente un travail fastidieux. Ensuite, une absence de normalité n'est mène pas concevable dans la perspective réductionniste servant généralement de support à l'hypothèse testée. Au-delà
des
fluctuations
immenses de
variabilité
parfois
enregistrées,
il peut
également
apparaître
des
distributions
plurimodales
(c'est-à-dire
l'existence de
plusieurs pics
dans une
distribution
de
fréquences),
et ce,
même sur
des
systèmes
biologiques
très
homogènes.
Par exemple,
dans les
embryons de
graines de
tournesol, la
quantité
d'ADN (1C) par
cellule suit
une
distribution
bimodale.
Celle-ci
correspond au
fait que les
plantules
issues de
graines
situées
en
périphérie
du capitule
contiennent en
moyenne 20 %
d'ADN en moins
que les
plantules
issues de
graines
développées
au centre
(16). Ce
phénomène
ne peut
être
regardé
comme un
artefact parce
qu'il se
reproduit
systématiquement
sur chaque
inflorescence.
C'est pourquoi
le calcul
d'une
quantité
moyenne d'ADN
(1C) ne
reflète
ici rien
d'autre qu'un
état
virtuel. Non
seulement la
valeur moyenne
ne
représente
aucun des
individus de
la population,
mais en plus
elle occulte
un
phénomène
remarquable
d'autocompensation
de la
quantité
d'ADN à
un moment
donné
du
développement,
dont
l'existence
est
révélée
par
l'impossibilité
de
sélectionner
des
lignées
de faible ou
haute
quantité
d'ADN (17). |
|
Notes: (16) Cavallini et al., 1989,, Nuclear DNA changes within Helianthus annuus L. : origin and control mechanism, Theor. appl. Genet. 77, 12-16 ; une pareille différence se retrouve chez Festuca arundinacea entre graines situées en haut et en bas de l'épi, voir Ceccarelli et al., 1992, Variations in genome size and organization within hexaploïd Festuca arundinacea. Theor. appl. Genet. 83, 273-278. (17) Cavallini et al., 1996, Nuclear DNA changes within Helianthus annuus L. : Variations in the amount and methylation of repetitive DNA within homozygous progenies, Theor. appl. Genet. 92, 285-291 |
||||
| |
|
4.2 - Deux exemples |
||||||
|
|
|
|
les brassinostéroïdes peu efficaces |
|||||
|
DSV* p 35-37 |
|
Les brassinostéroïdes (BRs) sont les phytostéroïdes, stéroïdes végétaux. On a donc voulu leur attribuer des rôles hormonaux calqués sur ceux connus chez les animaux. Mais ils ne circulent pratiquement pas et semblent pouvoir mimer l'effet de pratiquement toutes les substances informatives (comme l'éthylène, voir Trewavas, 1999 accessible en image sur books.google.fr). |
|
Trewawas A., 1999. The importance of individuality, in H.R. Lerner (ed.), Plant response to environmental stresses. From phytohormones to genome reorganization. marcel Dekker Inc. New-York, 730p |
||||
| |
|
«
Les BRs ont
réellement
commencé
leur
carrière
de
régulateurs
de croissance
à
partir du
moment
où l'on
a pu
identifier
quelques
situations
dans
lesquelles
l'effet des
BRs
était
proportionnel
à la
concentration
administrée.
Les BRs
seraient donc
bien des
hormones dont
l'action se
combinerait
avec celles
des autres
régulateurs
de croissance,
ce qui
justifie le
vaste spectre
d'influence
observé.
Initialement
réfractaires
au
modèle
traditionnel
de l'hormone,
les
phytostéroïdes
semblent
lentement s'y
plier. Dans cet exemple, l'effet sur la moyenne est reproductible à loisir, à condition de travailler sur des populations suffisamment grandes. Il peut de ce fait être utilisé afin de déceler la présence de BRs dans un extrait. Mais cette prédictibilité de l'effet ne préjuge en rien de l'action des BRs dans le gravitropisme. Cet exemple illustre la coexistence de deux niveaux de réalité complètement indépendants, un niveau ingéniérique (la mise au point d'un essai biologique fiable) et un niveau scientifique (le mode d'action des BRs sur la racine). Ainsi, la restriction de l'analyse à une comparaison de moyennes revient à extrapoler le niveau ingéniérique à la réalité biologique. Il n'est pas étonnant que cette approche conduise à une représentation des transformations propres au vivant sous la forme de «mécanismes» ressemblant à s'y méprendre à ceux développés par les ingénieurs cybernéticiens». |
|
 Figure 1.2. Effet des brassinostéroïdes sur le temps nécessaire à l'initiation de la réponse gravitropique chez le pois. PIGR : pourcentage de la population initiant la réponse gravitropique dans l'intervalle de temps (en min) indiqué en abscisse. D'après Amzallag G.N. et Vaisman J., 2006, |
||||
| |
|
|
|
la tolérance au NaCl, une réponse adaptative au milieu |
||||
|
DSV*p39s |
|
N. Amzallag a déjà publié de nombreux articles sur la tolérance des végétaux (notamment le sorgho) au NaCl. Certaines idées ont été vulgarisées dans son livre La Raison malmenée. De l'origine des idées reçues en biologie moderne , dans la cinquième partie (CNRS éd, 2002). Amzallag, 2001a: Developmental Changes in Effect of Cytokinin and Gibberellin on Shoot K+ and Na+ Accumulation in Salt-Treated Sorghum Plants, G. N. Amzallag, Plant biol (Stuttg) 2001; 3: 319-325 DOI: 10.1055/s-2001-16465;(abstract) |
|
La
tolérance
au NaCl peut
être
induite chez
le sorgho
grâce
à
l'exposition
des plants
à une
concentration
de 150mM
durant trois
semaines, et
ce,
uniquement, pendant
une
période
de
compétence
(cette notion
de
période
de
compétence
a
été
développée
en embryologie
par Rosine
Chandebois),
durant le
développement
précoce.
Les plantes
sont ensuite
capables de
résister
à des
concentrations
de 300mM de
NaCl,
létales
en absence
d'adaptation
induite
à la
salinité. |
|
Mais
cet exemple
peut aussi
montrer les
dangers de
l'analyse
statistique
trop rapide. |
||
| |
|
4.3 - Mesurer la variabilité |
||||||
|
DSV*p41s |
|
On peut considérer qu'il existe 4 sources de variabilité: |
|
-
l'individuation
(ou effet du
à l'
individualité
qui « résulte
de
l'orientation
de l'individu
vers une
trajectoire
particulière
dans un champ
de
possibilités
[je
préfère
parler de
champ de
dynamiques]
qui s'ouvre
à un
moment
précis
du
développement.»
(p 39) «
... si la
variabilité
reflète
une
réorganisation
individuelle
des
régulations
développementales
en
réponse
à un
changement
abrupt, alors
les
différences
de
viabilité
[dans
le cas
où des
organismes
sont
exposés
à un
stress]
devraient
être
relativement
réduites,
le
développement
aboutissant
à
l'émergence
de formes
généralement
viables et
harmonieuses». |
||||
| |
|
|
|
le coefficient de variation (CV) N. Amzallag propose de se servir de l'écart-type (noté ici S) comme indicateur de «quantité de variabilité» d'un caractère. Plus exactement il utilise le coefficient de variation défini comme suit: |
||||
|
CV (coefficient de variation)= 100 X SX(écart-type) / Xm (valeur moyenne) |
|
|
||||||
|
N. Amzallag propose de comparer les CV dans une série de données homologues afin de détecter une fluctuation de la stabilité d'un caractère ou encore sa canalisation****. |
|
« Même en conditions naturelles, où l'hétérogénéité génétique et environnementale est importante, le CV peut refléter une réalité biologique. Ce point est illustré par l'analyse d'une Labiée pluriannuelle, Origanum dayi, plante endémique du versant est des montagnes de Judée, vivant à la limite de la zone désertique. Les plantes de celle espèce produisent une huile essentielle dont il est possible d'analyser les composés principaux (22 molécules volatiles distinctes) dans chaque individu. En
échantillonnant
les individus
dans dix
stations
dispersées
sur l'aire de
répartition
de cette
espèce,
il est
possible de
calculer la
moyenne de la
représentation
de chacun des
22
composés
volatils de
l'huile
essentielle.
L'amplitude
des
différences
environnementales
entre stations
n'est pas
connue, mais
on constate
qu'elle agit
exactement de
la même
façon
sur tous les
composés
volatils
mesurés.
C'est pourquoi
on peut
considérer,
en
première
approche, que
la fluctuation
inter-stations
dans la
moyenne (soit
encore le CV
calculé
sur la base
des 10
moyennes
intra-station)
reflète
le
degré
de
plasticité
des
composés
volatils. Par
ailleurs, le
niveau de
canalisation
dans
l'accumulation
d'un
composé
volatil peut
être
considéré
comme
inversement
proportionnel
au CV
intrastation
(ou plus
exactement
à la
moyenne des CV
intra-stations). |
|
|
||||
|
Soil microvariations as a source of variability in the wild : the case of secondary metabolsime in Origanum dayi, Post, G. N. AMZALLAG, O. LARKOV, M. BEN HUR, and N. DUDAI, Journal of Chemical Ecology, Vol. 31, No. 6, June 2005, DOI: 10.1007/s10886-005-5283-4 |
|
|
****canalisation = le terme est utilisé par Conrad Waddington en 1942 pour désigner une contrainte sur une trajectoire définie. L'exemple le plus simple est celui donné par les chréodes ("routes obligatoires" selon Thom) dans son paysage épigénétique (voir ci-dessus et page sur les modèles). Il est aussi utilisé comme traduction de chanelling dans le métabolisme (voir Au-delà de la cellule, 2.1.2.c). |
|||||
| |
|
|
|
le degré de redondance des réseaux de régulation |
||||
|
Amzallag G.N., Seligmann H., Lerner H.R., 1995. Induced variability during the process of adaptation in Sorghum bicolor. J. Exp. Bot. 45, 1017-1024 réedité in Salinity: Environment - Plants - Molecules, 291-312,Springer Netherlands, DOI 10.1007/0-306-8155-3, 2004 (abstract) |
|
Dans
le cas
théorique
d'une double
chaîne
métabolique:
soit les
substrats XA
et XB,
les produits
intermédiaires
A et B, et C,
le produit
final unique.
Les
flèches
ab et ba
indiquent des
voies de conversion
de A en B et
réciproquement. |
||||||
|
|
|
|
Cas
I Cas
II Cas
III |
|||||
| |
|
On peut donc appliquer ce raisonnement à l'étude du gravitropisme de la racine de pois sous l'action des BRs vu ci-dessus. |
|
« ... il existerait deux voies parallèles de stimulation de la réponse de la racine mettant chacune en jeu un mécanisme plus ou moins rapide de courbure, la première des deux voies exprimée éliminant l'effet de l'autre. Dans ce cas, l'effet des BRs serait l'annulation de l'inhibition de la réponse rapide par la réponse lente.» |
||||
|
|
effet silencieux N. Amzallag fait judicieusement remarquer que dans le contexte d'un réseau redondant de régulation, un changement de moyenne indiquerait davantage la perte de la variabilité du réseau (qui tend à stabiliser la moyenne) par transformation du réseau en une chaîne de régulation, qu'une réelle variabilité. Il appelle ce paradoxe «l'effet silencieux». |
|
« Les BRs appliquées à des concentrations de 0,1 à 10 nM ne modifient pas de façon significative le rapport entre les deux types de racines (adventives et séminales) chez le sorgho. On est donc porté à conclure que les BRs n'ont aucune influence sur l'allocation des ressources entre ces deux organes. Mais c'est là ignorer les changements induits par le traitement aux BRs dans la variabilité: le CV augmente considérablement après application de 0,1 nM BRs, alors qu'il diminue après addition de BRs à 1 et 10 nM. Il y a donc ici un effet silencieux. Il suffit de changer le sens des flèches dans la figure 1.3 (ci-dessus) pour le représenter, puisque les deux types de racines dépendent pour leur croissance des mêmes ressources produites par les parties aériennes. Ainsi, l'étude de la variabilité révèle que les BRs jouent d'une façon très subtile sur les relations source-puits entre tiges et racines. À faible concentration (0,1 nM), ils accentuent l'asymétrie de distribution des tiges vers les deux types de racines, mais ils imposent ensuite (à partir d'une concentration de 1 nM) un mode de relation stabilisé. Les BRs n'exercent une influence sur le rapport tige/racine qu'à partir d'une concentration de l'or de de 10 nM. Or cette concentration est très éloignée de la concentration physiologique (sauf exception, les concentrations de BRs mesurées dans les tissus végétaux sont généralement en deçà de 0,1 nM). II semble donc que l'effet sur la moyenne reflète une linéarisation des processus de contrôle, qui est ici probablement un artefact. C'est donc en fait l'influence silencieuse sur le CV qui révèle ici le véritable mode d'action des BRs.» DSV*p44 |
|||||
|
|
exercice d'application : effet des BRs sur la vigne (Vigna radiata) dont les racines adventives sont exposées au sel. |
|
On observe (au niveau des valeurs moyennes) une stimulation de croissance à 20 nM de BRs et une inhibition après exposition à 500 nM de BRs. L'analyse des CV montre de plus que les racines dont la croissance est la moins inhibée sont celles pour lesquelles la valeur du CV est la plus faible. |
|
||||
| |
|
|
|
la connectance |
||||
|
de r, coefficient de corrélation d'une régression, à z, indicateur biologique de la variabilité |
|
Le
coefficient de
corrélation
d'une
régression
est
censé
représenter
le
degré
de
représentativité
de la fonction
de
régression
par rapport
à la
dispersion des
points de
mesure. |
|
Mais il existe un moyen de transformer le coefficient de corrélation r (surtout pour les faibles valeurs de r, dont la limite de représentativité est habituellement fixée aux valeurs de probabilité P < 0,05) afin qu'il présente une distribution quasi-normale et puisse donc être considéré comme une variable du système biologique étudié au moyen des paramètres corrélés. |
|
z est défini par où r est le coefficient de corrélation. C'est la valeur absolue de z que l'on utilise comme indicateur biologique d'une variation. N. Amzallag reprend ici l'exemple des 10 populations sauvages d'Origanum dayi présentées plus haut. |
||
|
|
|
Soil microvariations as a source of variability in the wild : the case of secondary metabolsime in Origanum dayi, Post, G. N. AMZALLAG, O. LARKOV, M. BEN HUR, and N. DUDAI, Journal of Chemical Ecology, Vol. 31, No. 6, June 2005, DOI: 10.1007/s10886-005-5283-4 |
|
«
Après
avoir
mesuré
la
quantité
de
matière
organique dans
le sol de
chacune des
dix stations,
il devient
possible de
calculer le
coefficient de
corrélation
entre le taux
de
matière
organique du
sol et la
valeur moyenne
(calculée
sur la base de
toutes les
plantes de la
station) de
chacun des 22
composés
principaux de
l'huile
essentielle. |
||||
|
|
|
|||||||
|
Celle-ci ne résulte pas uniquement de la position des coordonnées des 3 composants pour lesquels la valeur de r révèle une corrélation significative. Ce sont toutes les valeurs de z qui s'alignent de façon cohérente sur un tel graphe. Cela révèle que les valeurs de z peuvent servir à mettre en évidence un phénomène biologique, et ce indépendamment du seuil de signification de la corrélation. Dans le cas présent, il apparaît que les matières organiques présentes dans le sol conditionnent, dans son ensemble, le métabolisme secondaire générant les composés volatils. Cette conclusion n'a rien de très étonnant dans la mesure où le métabolisme secondaire est généralement organisé en un réseau d'interactions entre voies métaboliques. (---->) |
|
Par
l'usage des
statistiques
qualitatives,
l'expérimentateur
tend à
réduire
progressivement
son champ
(dans ce cas,
la
réduction
de 22 à
3
composés)
avant de
prolonger
l'investigation,
alors que le
contraire se
produit le
plus souvent
au travers de
l'approche
explorative
illustrée
ici. Ainsi,
les
conclusions de
l'expérience,
mais
également
le type de
questions se
trouvent
conditionnées
par le mode,
qualitatif ou
quantitatif,
d'approche de
la
régression. |
||||||
| |
|
La «connectance» d'une variable est la valeur moyenne de l'intensité de ses relations avec les autres variables exprimées en valeurs absolues de z. Dans RM* (p210), Amzallag utilise le terme légivité à la place de connectance (mot anglais). Définie pour une régulation en réseau , la légivité d'un paramètre mesure «combien la variation du paramètre en question est proportionnelle à la variation de toutes les autres variables étudiées». |
|
La
«connectance»
globale d'un
système
est la moyenne
de la valeur
absolue de
tous les z. «Dans
ce cas elle
reflète
la structure
du
réseau
de relations
entre
unités
biologiques
homologues
(individus,
organes,
cellules,
organites,
protéines,
molécules,
etc.). Sa
valeur est
influencée
par le
degré
de
ramification,
de redondance
du
réseau,
ou encore
l'intensité
des relations
entre
unités». |
||||
|
|
|
Exemple: |
|
« Comparons la connectance et le CV pour une série de caractères de fins de cycle mesurés sur des populations homogènes de sorgho (descendant chacune par autofécondation d'un seul parent) exposées au même milieu. Une relation non-linéaire, en forme de V, apparaît sur le graphe. La canalisation maximale d'un caractère (les valeurs du CV les plus faibles) correspond à une valeur intermédiaire de la connectance (autour de 0,5), tandis qu'un fort CV, reflétant une faible canalisation (soit encore une forte influence du milieu sur l'expression du caractère) s'observe pour de faibles ou très fortes connectances. Ce résultat correspond au fait qu'une faible canalisation (forte influence des perturbations diverses dans l'expression d'un caractère) se manifeste dans deux cas distincts: une faible connectance (isolement de la régulation du caractère par rapport au réseau), ou une très forte connectance (réorganisation du réseau autour d'un seul et unique facteur chapeautant l'ensemble). (----->) |
|
Connectance in Sorghum development: beyond the genotype-phenotype duality, G.N. Amzallag, BioSystems 56 (2000) 1-11 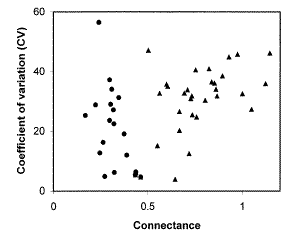 Fig. 2. Relations entre la connectance et la variabilité (coefficient de variation = CV). On observe une corrélation positive signifiante (r = 0.412, PB = 0.001, 46 df). Les données ont été séparées selon la valeur de connectance 0,5 : des cercles pour les valeurs inférieures à 0,5 et des triangles pour les valeurs supérieures de la connectance. |
||
| |
|
Remarque: |
|
|||||
| |
|
Dans les deux cas, l'expression du caractère devient conditionnée par un nombre minimum de facteurs, eux-mêmes influençables par les diverses variations susceptibles de perturber son expression.» |
||||||
| |
|
4.4 - Une nouvelle épistémologie à construire Les propos qui suivent peuvent paraître s'éloigner du thème de cette page (les mutations) mais il faut garder à l'esprit que la plupart des résultats expérimentaux en biologie du développement qui présente des phénomènes non-linéaires sont interprétés à grand renfort de mutants imaginaires surgis au hasard et sélectionnés par le milieu. Dans cette partie N. Amzallag nous emmène sur d'autres pistes. |
||||||
|
DSV*p 51s |
|
Les exemples proposés ci-dessus restaient au sein du paradigme génétique avec un réseau de régulation, même redondant, qui reposait sur des gènes supposés être responsables des différents éléments du réseau. On est toujours dans une vision linéaire du développement de l'organisme, sous le contrôle d'un programme génétique. Mais il est évident que cela n'est pas l'interprétation la plus simple ni la plus évidente. |
|
« Comment imaginer un mode darwinien d'évolution de gènes codant pour des voies parallèles, alors que la redondance contrecarre toute possibilité de les sélectionner de façon spécifique ?» |
|
Au contraire, il est patent que les outils d'analyse expérimentaux mis en place précédemment peuvent nous aider à caractériser de façon plus exacte les changements de phase lors du développement, plutôt qu'à les considérer comme des changements arbitraires dans le programme de développement. En effet, la stabilité d'un réseau, et plus encore celle d'un réseau redondant, porte à penser que l'émergence d'une nouvelle phase de développement ne peut se faire que par interruption du réseau existant et mis en place d'un nouveau réseau. Si l'on utilise la connectance entre organes comme indicateur de la stabilité du réseau de régulation, on devrait pouvoir noter, au cours du développement, des modifications importantes de cette connectance, lors des interruptions des réseaux existants. |
||
|
|
|
« Il est difficile de mettre en évidence un tel changement par la mesure de corrélation entre poids des différents organes, parce que les modifications se produisant sur un faible intervalle de temps n'y laissent que peu de traces visibles. Par contre, l'expansion foliaire, s'effectuant sur un bref intervalle de temps, est un indicateur sensible, susceptible de rendre compte des brefs changements dans les régulations. Par exemple, chez le sorgho, il est possible de calculer la corrélation entre la taille de la gaine des feuilles successives et le poids de la plante à un jour donné. Si la croissance était un phénomène continu, alors on devrait observer une corrélation d'intensité croissante entre la taille de la gaine et le poids de la plante au fur et à mesure que l'on considère des feuilles les plus récemment développées. Mais en réalité, on observe que la connectance baisse progressivement de la 3e à la 5e feuille, pour remonter ensuite à la 6e et à la 7e feuille. Cela implique qu'il existe un moment, celui de l'émergence de la 5e feuille, durant lequel la croissance est relativement déconnectée des événements ultérieurs. Cette observation appuie l'idée qu'une discontinuité dans la régulation de la croissance peut se manifester au cours du développement (note 59). L'événement observé durant l'émergence de la 5e et 6e feuille correspond justement à une phase où, chez le sorgho, les racines adventives deviennent l'appareil racinaire principal de la plante (note 60). Peu avant l'émergence de la 5e feuille, l'ensemble du réseau de relations semble se défaire pour se reconstituer à partir de l'expansion de la 6e feuille (note 61). Observé sur des plantes exposées à des conditions optimales, ce phénomène semble inhérent au développement normal. Il témoigne de la refonte de l'ancien réseau de régulation (parties aériennes et racines séminales) inhérent à l'intégration d'un nouveau type d'organes (les racines adventives). Cette intégration coïncide justement avec la période d'adaptation au sel (note 62). L'augmentation induite de tolérance ne serait que le produit de l'intrusion de facteurs perturbateurs supplémentaires (en l'occurrence le NaCI) dans un processus d'adaptation de la plante à une perturbation endogéne (l'émergence de racines adventives) générée par son propre développement. Confirmée par certaines observations (note 64) cette interprétation conduit à une représentation dans laquelle l'échelle de temps propre au développement n'est plus continue. On y découvre un processus composé de périodes de stabilité (les phénophases) durant lesquelles un réseau de régulation régit les relations entre entités. et de périodes critiques durant lesquelles s'opère une restructuration de ces réseaux de régulation conduisant à leur auto-réémergence (fig. 1.4 ci-dessous). |
|
note 59 Ce phénomène est même amplifié par l'addition de BRs dans le milieu, suggérant que ces hormones sont impliquées dans l'expression de cette non-linéarité dans la régulation de la croissance) note
60 note
61 Figure
3. Relations
entre la
connectance
des feuilles
et la
compétence
pour
l'adaptation
au sel. (a)
Connectance
moyenne pour
les feuilles 3
et 4. (b)
Connectance
moyenne pour
les feuilles
5-7. Une
corrélation
non
signifiante
(r=0.254,
P>0,50) est
observée
en (a). Une
corrélation
signifiante et
positive
(r=0,724,
P<0,05) est
observée
en (b). note
62: note
64: |
||||
|
|
|
Cette réalité est généralement éludée parce que le fort degré de redondance des régulations biologiques masque l'expression des processus auto-organisés, et parce que la variabilité est une dimension négligée de l'étude des organismes. Or pour mettre en évidence une période critique, il est nécessaire d'appliquer à un moment précis une perturbation susceptible par son intensité de conditionner le devenir du systéme, phénomène qui se traduit avant tout par une augmentation du niveau d'individualité.» |
|
|||||
|
|
|
Cette vision du développement replace correctement la notion d'information génétique en la libérant de l'imaginaire d'un contrôle fonctionnel (des gènes pour des produits et non des gènes pour des fonctions, voir cours de 1èreS). |
|
N. Amzallag souligne que l'on comprend ainsi comment l'expression de certains gènes, qualifiés un peu vite de morphogénétiques, peut revêtir, durant certaines périodes critiques, une signification morphogénétique alors que durant une phénophase, ils "codent" pour des protéines n'ayant rien à voir avec un phénomène d'induction de la morphogenèse. Ainsi, si l'on perturbe un de ces pseudo-gènes-morphogénétiques, on provoque une perturbation grave de l'émergence d'un nouveau réseau lors d'une phase critique, qui peut conduire à une morphogenèse anormale ou décalée. On notera que les expériences de N. Amzallag ont été menées sur des plantes mais qu'il paraît tout à fait légitime de tenter une extrapolation au développement des animaux qui présente des phases larvaires et juvéniles homologues de la phase végétative des plantes. Le contrôle de l'expression des gènes durant les périodes critiques permet de comprendre la stabilité du développement, malgré la variabilité inhérente à une espèce. Alors que les variations des processus auto-émergents, associés à ces mêmes phases critiques, permettent de comprendre pourquoi l'individuation, lorsqu'elle est observée, est si étroitement liée à l'adaptation. |
||||
|
|
|
|
||||||
|
|
||||||||
|
Remarque
philosophique:
d'après E.U. (article "épigenèse" par Alain Delaunay) |
|
On rapporte souvent À TORT ces deux théories à une différence dans la matière alors qu'elles concernent la forme substantielle qui donne vie et structure à la matière indifférenciée (aristotélicienne). L'épigénomique et surtout l'épigénétique moderne s'inscrivent dans cette lignée où le sens des mots ne cesse d'évoluer. |
|
Les épigénistes considèrent que la forme individuelle n'est pas visible au début de la vie (que l'on placerait maintenant à la fécondation) et se manifeste progressivement au cours du développement (au sein de la mère). Il est bien sûr évident que cette forme existe dès le début puisque c'est elle qui donne l'être à la matière. Cette théorie sera reprise par exemple par Piaget ou Freud dans le développement psychique. Les préformistes considèrent que l'individu a déjà une forme complète dès le début de sa vie. cette forme est héritée de la mère, pour les ovistes, et du père, pour les animaculistes. La forme se déplie ensuite ou s'agrandit. |
|
Au
XVIIème
siècle
on retrouve
ces deux
termes mais
qui
désignent
des processus
différents
: Quand on lit : « Leeuwenhoek (1675) croyait que le spermatozoïde contenait un individu en réduction», c'est un peu juste. Par son observation des spermatozoïdes dans la semence masculine, il apportait des arguments aux animaculistes préformationistes, mais il ne semble pas d'ailleurs qu'il se soit douté de leur rôle, ni de la querelle sur la forme. C'est Spallanzani qui démontrera la rôle des spermatozoïdes dans la fécondation chez la grenouille (1777) alors qu'il était paraît-il un oviste convaincu; la fusion des cellules n'étant observée qu'en 1875 par Hertwig. |
||
| |
|
|
|
|
|
|
||
|
Réponse de l'exercice d'application sur les BRs |
|
Plutôt que de considérer qu'il existe un unique effet moyen pour toutes les racines, mais variable et opposé selon la concentration, on peut d'abord considérer qu'il existe deux effets : l'un activateur, l'autre inhibiteur. |
|
L'effet activateur s'exprime aux deux concentrations de BR puisque le CV (qui exprime la réduction de la variabilité autour de la valeur moyenne) est plus faible lorsque la croissance est moins inhibée dans les racines adventives qui globalement ont une croissance inhibée. Le sens de la relation entre moyennes et CV révélerait donc un effet activateur direct (permanent). |
|
L'effet inhibiteur, apparent seulement aux fortes concentrations de BRs (et ce, bien au-delà des concentrations physiologiques), serait un artefact expérimental qui masquerait l'effet activateur. Cet effet inhibiteur serait indirect puisque non permanent. |
||


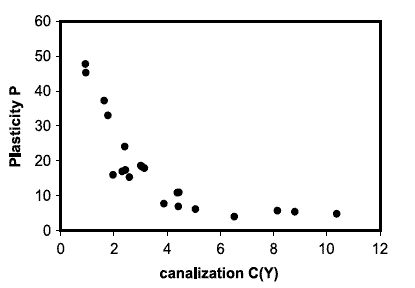
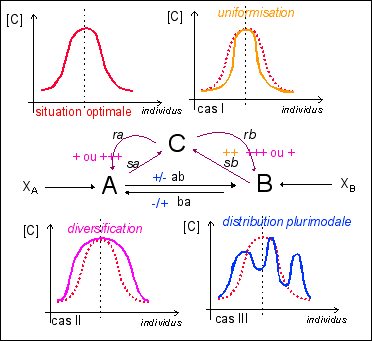
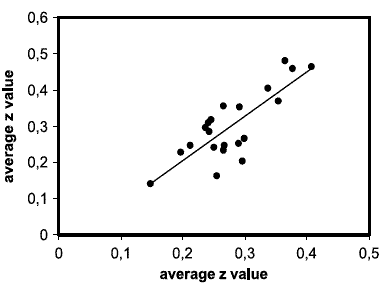
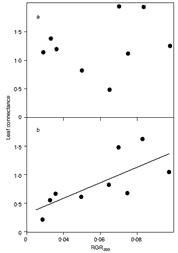
{kind=link}