|
La
nature*
humaine est
unique... ou
plutôt l'humain
se
différencie
profondément
de l'animal et
forme une
espèce
à part...
Une définition de l'homme, due à Linné, 1758 : animal rationale, loquens, erectum, bimanum (animal rationnel, doué de parole, en position érigée (station droite), pourvu de 2 mains). |
|
*Le terme de nature, à forte connotation métaphysique, peut, si l'on veut éviter une forme de dualisme stérile, être remplacé par "genre humain" ou "humanité". Le terme de genre a pris récemment une connotation bien particulière qu'il est indispensable d'éviter. Les biologistes utilisent le continu de l'espèce ; il y a donc une espèce humaine. voir page sur l'identité biologique de l'homme
**L'anthropologie (du grec anthropos = l'homme) qui s'appuie sur la philosophie est la science de l'homme en tant qu'être vivant semblable à nul autre. |
|||||||||||||||||||||||||||||||||||
|
|
La position anthropologique (très fortement minoritaire dans le monde, mais qui fleurit dans l'Éducation Nationale en France) qui réduit l'homme à un animal comme les autres est une opinion scientiste qui n'a rien de scientifique. Tout scientifique doit pouvoir aborder les questions d'anthropologie avec ses convictions sans être l'objet de suspicion de malhonnêteté ou de parti-pris anti-scientifique. |
|
Dans cette page, comme annoncé ci-dessus, l'homme n'est pas qu'un animal. De plus, l'expérience n'est pas le bon outil pour étudier l'histoire de l'homme. |
||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||
|
La paléontologie n'est pas une science expérimentale |
|
La paléontologie n'est pas une science expérimentale (au sens où l'on ne fait pas des expériences sur le passé : la vie passée n'est pas un phénomène actuel, c'est une "donnée historique", que l'on peut s'efforcer de reconstituer à l'aide d'archives, de témoignages plus ou moins directs). Faire "parler" les fossiles à l'aide d'outils mathématiques faisant appel à des sciences expérimentales (comme la datation isotopique) ne change pas le caractère profondément original d'une science des êtres vivants du passé. |
|||||||||||||||||||||||||||||||||||
|
|
|
La paléoanthropologie n'est pas une science biologique, mais nécessite une approche à la fois anthropologique et à la fois paléontologique (de paléo = ancien, ontos = l'être et logos = parler). La
paléontologie
utilise les
sciences
expérimentales
biologiques
(anatomie,
embryologie,
physiologie...)
dont elle
extrapole les
résultats
dans le
passé. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||
|
|
|
1 . L'évolution
c'est
l'histoire de
la
transformation
des
espèces
:
l'espèce
humaine est
apparue et se
transforme
peut-être |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||
|
1 . L'évolution c'est l'histoire de la transformation des espèces : l'espèce humaine est apparue et se transforme peut-être Cette partie me semble une base essentielle: elle est traitée sous forme de 2 fiches utilisées dès la classe de seconde |
|
|
|||||||||||||||||||||||||||||||||||
|
|
L'évolution
est une
idée
qui est
maintenant
acceptée
par le plus
grand nombre:
les
espèces
dérivent
les unes des
autres. L'homme est une espèce. À ce titre, il est donc le fruit d'une évolution des espèces. Mais l'homme est très récent. Les fossiles que l'on possède sont très peu nombreux. Évolue-t-il encore ? L'évolution n'est-elle pas en voie terminale ? (c'est notamment la thèse de PP Grassé : l'amortissement de l'évolution). La controverse se limite donc souvent à l'origine de l'homme plutôt qu'à son évolution. |
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||
|
Pour
échapper
à cette
vue
étroite
de la
spéciation,
il faut : Pour replacer cette question dans une perspective historique et épistémologique je recommande les travaux d'André Pichot (voir page sur l'évolution). |
|
les mécanismes génétiques de l'évolution proposés dans la théorie synthétique et cités dans le programme Le contenu de cette partie est clairement dépassé, tel qu'il est formulé, et peu convaincant si l'on essaie de l'appliquer à l'évolution de l'homme |
|||||||||||||||||||||||||||||||||||
|
|
|
|
La sélection naturelle était comprise semble-t-il par Darwin comme une reproduction différentielle; elle a ensuite été comprise par les néodarwiniens comme l'ensemble des contraintes sélectives externes qui entourent le développement des populations (et donc directement des contraintes sur le génome qui est au cœur de la notion de population).
Voir quelques citations, anciennes, sur l'idée de sélection naturelle. |
||||||||||||||||||||||||||||||||||
|
|
La microévolution repose principalement sur les mutations. Elle engendre une variation intraspécifique (au sein de l'espèce). Les caractéristiques de l'espèce sont plus ou moins figées dans un pool génétique transmis entre individus dans une population génétique |
|
La mégaévolution, évolution interspécifique (le passage d'une espèce à une autre) reste très discutée. Un
mécanisme
possible
pourrait
être
constitué
par des réarrangements
chromosomiques
(comme ceux
que l'on peut
voir entre le
chimpanzé
et l'homme:
délétion,
translocation,
duplication...). |
|
|||||||||||||||||||||||||||||||||
|
Voici les mots exacts du programme qui constituent un cours (dont le contenu est pour moi inacceptable tel quel...): |
|
Les mécanismes de l'évolution |
|||||||||||||||||||||||||||||||||||
|
|
Les génomes des espèces sont des archives. Ils permettent d'imaginer les événements génétiques moléculaires de l'évolution qui ont conduit à des innovations, à leur diversification et à leur complexification (familles multigéniques, gènes chimères...). Ces innovations génétiques sont aléatoires ; leur nature ne dépend pas des caractéristiques du milieu. L'évolution des génomes résulte d'un bricolage moléculaire qui a conduit à faire du neuf avec du vieux. |
|
Ainsi, l'acquisition de la bipédie dans la lignée humaine ne fait pas intervenir une explication finaliste. À l'origine de la bipédie se trouvent des innovations génétiques. Elles ont dû affecter les gènes du développement.Les conditions de l'environnement peuvent jouer le rôle de crible vis-à-vis des nouveautés phénotypiques engendrées par les innovations génétiques (sélection naturelle). |
|
De ce fait, l'évolution dans la lignée humaine comme dans les autres lignées peut être dépendante de changements dans l'environnement. Elle est contingente. |
||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||
|
2. Les phylogénies sont des classifications évolutives : l'espèce humaine est regroupée avec de proches parents dans le groupe des Hominiens ou des Homininés ou des Hominina selon les classifications |
|
|
|||||||||||||||||||||||||||||||||||
|
Depuis les développements faits par les disciples de Darwin les querelles autour de la place de l'homme dans les classifications tournent trop autour de comparaisons (moléculaires et autres) entre l'homme et les grands singes africains. |
|
Précis de Zoologie, Vertébrés (t. 2), PP Grassé, Masson, 1977, p 109, p 268 |
|||||||||||||||||||||||||||||||||||
| |
|
2.1 On classe l'homme dans les Primates |
|
|
|||||||||||||||||||||||||||||||||
|
|
|
Rien n'est plus mal connu ou volontairement occulté que l'histoire du concept de race humaine et de ses relations avec les espèces de singes. Les ressemblances entre l'homme et les singes (chimpanzé, mais surtout orang-outan, ont depuis fort longtemps (au moins le XVIIème siècle) donné lieu à des classifications qui les regroupaient tout en les différenciant (Primates, bi-quadrumanes...). Dès le XVIIIème siècle Jean-Baptiste Robinet propose l'orang-outan comme intermédiaire entre l'homme et le singe en supposant une véritable parenté. Le schéma évolutif deviendra plus ou moins orang-outan -> hottentot (ou negrito)-> noir -> blanc. Des thèses similaires seront reprises par les évolutionnistes au XIXème siècle. |
|||||||||||||||||||||||||||||||||||
|
|
|
Les représentations phylogénétiques reflètent la classification choisie. Il ne faut pas accorder une trop grande importance aux classifications qui sont de toute façon des constructions humaines, même si je crois qu'il existe un ordre dans la nature et donc une classification naturelle. Les querelles autour de l'importance de telle ou telle représentation masquent souvent des positions philosophiques qui, pour le scientifique, ne sont que des points de vue face à une réalité toujours plus mystérieuse, selon le mot de Charles Péguy sur la page d'accueil. |
|||||||||||||||||||||||||||||||||||
|
Les 3 anciens types de classifications |
|
Historiquement, depuis le milieu du XXème jusqu'au début du XXIème siècle, on a distingué 3 types de classifications: |
|
|
|||||||||||||||||||||||||||||||||
|
thèse de Richard Blanchette, Le problème de la classification en zoologie, Université Laval (Québec), PhD, juin 2002) que l'on peut trouver in extenso au format PDF sur internet à l'adresse: http://www.theses.ulaval.ca/2002/20168/20168.pdf) |
|
Les graphes phylétiques (cladogrammes) ne représentent pas l'histoire des espèces, mais des liens de ressemblance. Ils incluent la dimension évolutive (tel caractère dérive phylogénétiquement de tel autre, ce qui n'est pas vrai pour l'espèce) mais ils ne représentent pas le temps (telle espèce est apparue avant telle autre). |
|
Les graphes phénétiques (phénogrammes) ne représentent que des ressemblances sans dimension évolutive : telle espèce ressemble plus telle autre espèce qu'aux autres (par la similitude de davantage de caractères), ni temporelle. |
|
Les arbres évolutifs (phylogrammes) tentent de représenter l'histoire des espèces (généalogies) en hiérarchisant les caractères considérés vis-à-vis de l'évolution. Ils représentent donc à la fois l'évolution et le temps. |
|||||||||||||||||||||||||||||||
|
approfondissement : fiche cladisme |
|
Dans
les deux
premières
méthodes
(à
connaître
pour le bac),
on construit
(mathématiquement)
des graphes
phylétiques
représentant
les
parentés
entre les
espèces
à
partir de
tableaux de
caractères.
La distance
entre deux
représentants
(taxons) est
d'autant plus
grande que la
parenté
est
éloignée.
Aucun de ces
deux types de
graphes ne
devrait
être
représenté
sous forme d'arbres
enracinés
vers le bas
avec le temps
verticalement,
car aucun ne
prend en
compte le
temps. |
|||||||||||||||||||||||||||||||||||
| |
|
Dans
l'enseignement,
forcément
décalé
par rapport
à la
recherche, la
grande
avancée
réalisée
depuis les
années
1990 à
été
l'utilisation
généralisée
de la
méthode
cladiste, avec
des
réussites
plus ou moins
marquées.
On saluera
l'effort de
vulgarisation
scolaire
réalisée
par certains
pédagogues
- G.
Lecointre
est, à
cet
égard,
exemplaire -
(voir site
acces-évolution
de l'INRP,
strictement
dédié
à cette
vue partielle)
. Très
récemment
encore, des
présentations
réalisées
à
partir des
collections
muséographiques,
tentent de
populariser
ces
mêmes
classifications
cladistes, en
surmontant les
difficultés
de
précision
de vocabulaire
et leur
méconnaissance
par le public
des
musées
d'histoire
naturelle.
Pour ma part,
je regrette
que, la
plupart du
temps, le
relativisme
philosophique
soit
véhiculé
avec le
cladisme, ce
qui est,
à mon
avis,
inacceptable,
mais pas
irrémédiable
(voir nommer-classer). |
|||||||||||||||||||||||||||||||||||
|
L'irruption des statistiques |
|
Depuis le début du XXIe siècle, le fort développement des statistiques, notamment du fait des études phénétiques et cladistiques moléculaires, a changé le paysage de la recherche phylogénétique en provoquant une sorte de mélange entre les 3 types anciens de classification, si bien qu'il est devenu très difficile de s'y retrouver. Les statisticiens (des techniciens des mathématiques ?) qui travaillent sur des modèles évolutifs (écologiques notamment....) sont peut-être peu intéressés par les théories biologiques sous-jacentes à leur recherche et il n'est pas facile de décrypter, à partir des articles qui publient des phylogenèses, quels sont les modèles utilisés. De façon quasi générale, le paradigme moléculariste est la norme, mais, pour l'évolution, il y a par contre un flou indubitable. On pourrait même parler d'une absence de théorie évolutive; les mécanismes moléculaires évolutifs étant inférés, par les statisticiens eux-mêmes, à partir de leurs résultats, ce qui est épistémologiquement fort dangereux. |
|
Ainsi, l'article de l'EU2009 de P. Duris et P. Tassy sur la classification du vivant conclut: « Il n'est pas besoin de préciser que la critique mutuelle que se font parfois les partisans des méthodes probabilistes et les partisans des méthodes cladistiques peut se révéler aussi féroce que lors des débats des années 1970 entre les évolutionnistes, les cladistes et les phénéticiens. Certains considèrent que les résultats tirés des approches probabilistes sont un nuage de fumée, d'autres que la morphologie n'est pas fiable et a fait son temps. ... [] Si un siècle après L'Origine des espèces, on a admis que les classifications devaient être phylogénétiques, on s'est vite désaccordé sur les méthodes permettant de reconstruire la phylogénie. Cet aspect n'apparaît pas aux yeux du grand public : un résultat phylogénétique est le plus souvent présenté comme un résultat biologique qui va de soi, s'imposant de lui-même, où la méthode de travail n'est pas censée influencer le résultat. Ce n'est pas tout à fait vrai. Alors que la classification a d'abord été vue comme un système de référence invitant à la stabilité du savoir, la phylogénétique est un domaine en recherche permanente.» |
|||||||||||||||||||||||||||||||||
|
|
|
Dossier Sagascience de Guillaume Lecointre; avec des bémols pour une terminologie inadéquate (comme gènes "lents/rapides") et ses habituelles vues scientistes (il est le héraut d'une classification "laïcisée", selon ses propres mots); l'important est son travail de pédagogue.
un exemple récent (juin 2008) d'approche statistique avec des caractères morphologiques : clade de la lignée humaine sur la page du cladisme |
|
Actuellement des mots, dont le sens original est pourtant si différent, comme phylogénie, généalogie et classification, sont donc devenus quasiment synonymes ou du moins sont employés indifféremment lorsqu'il s'agit de parler d'une méthode scientifique moderne de classification évolutive (voir nommer-classer). |
|
||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||
|
L'homme
a une histoire
qui s'enracine
dans l'arbre
phylogénétique |
|
|
|||||||||||||||||||||||||||||||||||
|
Les Mammifères
(classe)
sont des animaux,
vertébrés,
possédant
des mamelles
(ce qui a
donné
le nom au
groupe : du
latin mamma
= la mamelle
et fereo =
je porte) mais
aussi de
nombreux
autres
caractères: |
|
Les Primates
(ordre) sont
des euthériens
(à
développement
embryonnaire
long se
déroulant
dans
l'utérus,
à
mandibule sans
processus
angulaire
recourbé
en dedans,
à
coracoïde
réduite
soudée
à
l'omoplate,
à corps
calleux,
à
denture
diphyodonte;
ces
caractères
sont
partagés
avec la
plupart des
Mammifères
qui ne sont
pas Marsupiaux
(kangourous...)
ou
Monotrèmes
(fourmiliers...)...),
à
très
grande
majorité
arboricole
(vivent dans
les arbres)
qui se
caractérisent
par: |
|||||||||||||||||||||||||||||||||||
|
L'homme au sein des primates |
|
|
|||||||||||||||||||||||||||||||||||
|
L'évolution des Primates, Marc Godinot, Pour la Science, L'évolution, Belin, 1998, p 94-100 |
|
Darwin,
Huxley,
Haeckel et les
premiers
"évolutionnistes"
placèrent
l'origine de
la
lignée
hominienne
chez les
grands singes
anthropomorphes
: les
hommes
n'étaient
que des grands
singes
évolués,
eux-mêmes
provenant des
Lémuriens. |
|
Dans
l'ancienne
classification
phylogénétique
l'homme
était
classé
dans le
sous-ordre des
Hominiens
qui
étaient
séparés
des 3 autres
sous-ordres de
Primates:
Tarsiens,
Lémuriens
et Simiens.
Les grands
singes
anthropomorphes
(ce
qui signifie
"de même
forme que
l'homme") ou
Simiens
Catarhiniens,
étaient
donc
classés
à part;
ils
comprennent
les Gibbons
(Hylobates),
les Orangs-Outans
(Pongo),
les
Chimpanzés
(Pan)
et les Gorilles
(Gorilla). |
|||||||||||||||||||||||||||||||||
|
d'après
E.U. Primates
fossiles
(Denis
Geraads), v 12
- 2006 http://www.hominides.com/html/ dossiers/grands_singes.htm La planète des singes du Miocène, David Begun, Pour la Science, dossier 57, oct-déc 2007, pp 104-111 |
|
Dans les classifications phylogénétiques modernes, les caractères chromosomiques et moléculaires (ADN mitochondrial, certains gènes...) ont été très largement intégrés aux matrices de caractères qui permettent de construire les arbres. Les phylogénies cladistes qui ne comprennent que des caractères paléontologiques, anatomiques et physiologiques amènent PARFOIS à des relations logiques incompatibles avec les phylogénies établies à partir des seuls caractères moléculaires. Depuis la généralisation des classifications cladistes relativistes on est loin d'avoir une unanimité sur le sens des noms: c'est plus que déroutant pour les enseignants et que dire des élèves.
|
|
La classification phylogénétique la plus courante :
Le
terme d'Homininæ
(homininés)
désigne
alors
l'ensemble des
grands singes
africains
(gorille,
chimpanzé
et
fossiles
apparentés)
et de l'homme
(et
des fossiles
apparentés)
séparé
ainsi des
orangs-outans
(et
des fossiles
apparentés)
au sein de la
famille des Hominidæ
(hominidés). Une magistrale mise au point de Véronique Barriel (maître de conférences au Muséum National d'Histoire Naturelle) pour s'y retrouver dans les classifications (Hominés notamment) http://www.snv.jussieu.fr/vie/ dossiers/evolution/ ligneehumaine/ homo.htm.;
(super-famille) Hylobates
(gibbon) et
fossiles (famille) Pongo
(orang-outan)
et
fossiles Gorilla
et
fossiles (tribu) Pan
(chimpanzé)
et
fossiles Homo
et
fossiles et
australopithèques |
|||||||||||||||||||||||||||||||||
|
pour une approche exhaustive des terminaisons des groupes (æ, i,... voir le site http://www.iczn.org/iczn/index.jsp - en anglais). pour une approche statistique récente (juin 2008) voir le clade de la lignée humaine sur la page du cladisme |
|
||||||||||||||||||||||||||||||||||||
| |
|
2.2 L'homme est proche du chimpanzé |
|
|
|||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||
|
Les querelles les plus persistantes concernant les arbres phylétiques viennent de l'importance accordée aux données moléculaires ou chromosomiques ou de l'origine des espèces ou sous-espèces ayant servi à établir les parentés moléculaires Les données moléculaires ne sont bien sûr accessibles que pour les seuls individus actuels. La parenté s'exprime dans les arbres phylogénétiques par les distances génétiques et chromosomiques (source : Origine de l'homme, Jean Chaline, Encyclopedia Universalis v12, 2007) |
|
distance
génétique
: distance
chromosomique |
|||||||||||||||||||||||||||||||||||
|
|
Personnellement j'ai toujours pensé que puisque la similitude moléculaire et chromosomique entre les grands singes et l'homme était si forte, c'était bien la preuve qu'il fallait chercher ailleurs la cause de leur profonde différence biologique. L'universalité
des
génomes |
||||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||
|
source
: Origine
de l'homme,
Jean Chaline,
Encyclopedia
Universalis
v12, 2007 Anne Dambricourt-Malassé: Évolution du chondrocrâne et de la face des grands anthropoïdes miocènes jusqu'à Homo sapiens, continuités et discontinuités, article de Palevol (Compte-Rendu de l'Académie des Sciences , Palevol, 5, 2006). Une copie html est accessible sur internet à l'adresse: http://carpediemcom.free.fr/ ADMPalevol05.htm *Voir aussi une ancienne page sur les modèles; pour la notion d'attracteur *voir la page sur les modèles thomiens ainsi que la page de compléments mathématiques et la page sur le continu
N.B. Le texte ci-contre, extrait de l'encyclopédie Universalis, est de Jean Chaline qui rend hommage au très intéressant travail de Mme Dambricourt-Malassé (voir référence ci-dessus par exemple et bibliographie). Cette chercheuse fait encore souvent l'objet de médisances éhontées, venant toujours des mêmes personnes, et qu'il serait bon de voir cesser dans l'éducation nationale. Cette institution n'a pas à faire la propagande du scientisme athée (néopositivisme). |
|
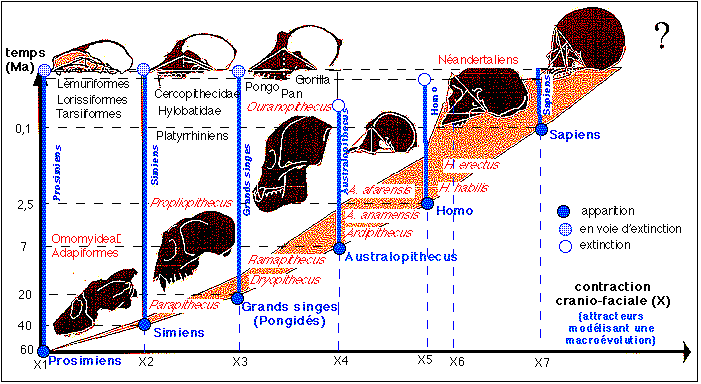
« La comparaison des développements des singes supérieurs et de l'homme a montré que, si on se limite aux seules modifications du crâne, on constate que l'évolution des primates est contrôlée par un phénomène majeur : l'augmentation du volume cérébral entraîne une diminution de la face et un élargissement de la mandibule. Ce phénomène dynamique, connu sous le nom de contraction craniofaciale, a été étudié par Antoine Delattre et Raphaël Fenart, et plus récemment par Anne Dambricourt-Malassé et Marie-Josèphe Deshayes. Comme ce phénomène est inhérent à l'ensemble du groupe des primates, et reconnaissable dans la plupart des lignées de mammifères, on doit tenir compte de cette contrainte de développement si l'on veut comprendre les changements évolutifs qui se produisent lorsque l'on passe des singes supérieurs (gorille et chimpanzé) aux australopithèques, ou singes bipèdes, et enfin aux hommes. Anne Dambricourt-Malassé a pu montrer que dans l'histoire des primates on pouvait distinguer six étapes majeures, correspondant à six plans d'organisation morphologique crânienne distincts, ce qu'elle appelle des ontogenèses fondamentales. Ces plans sont caractérisés par des contractions crânio-faciales de plus en plus fortes, excluant tout état intermédiaire. Or les étapes de contraction crânio-faciale sont conditionnées par des décalages de la chronologie du développement [voir hétérochronies] pouvant toucher toutes les étapes de la formation, qu'elles soient embryonnaires, fœtales, lactéales (première dentition), de substitution (remplacement des dents de lait) ou adultes. Si l'on compare les développements respectifs d'un chimpanzé et d'un homme, on constate un ralentissement généralisé du développement humain et un doublement de la période de croissance.» (suite ci-dessous) |
|
«
Trois
changements
majeurs sont
intervenus: le
premier entre
l'ancêtre
commun et
l'australopithèque,
le
deuxième
entre
l'australopithèque
et l'homme
archaïque,
le
troisième
entre l'homme
érigé
et l'homme
moderne. |
|||||||||||||||||||||||||||||||||
|
Les animations parlantes qui illustrent la dynamique d'une contraction crânio-faciale selon le modèle de Vincent Fleury (un des co-auteurs de Génétiquement indéterminé, 2007, Quæ), biophysicien à Paris 7, sont à nouveau accessibles par internet. Légendes à lire absolument. passé
-> erectus
-> futur (fichier
.avi - 356 Ko) Vincent Fleury - De l'œuf à l'éternité - Le sens de l'évolution, Flammarion, 2006. |
|
«
La phase
embryonnaire,
qui dure deux
semaines chez
le
chimpanzé,
est
prolongée
jusqu'à
huit semaines
chez l'homme,
et c'est
durant cette
seule phase
que se
constituent
les cellules
nerveuses.
L'allongement
de cette
période
chez l'homme
entraîne
donc une
hypertrophie
du cerveau,
car nos
neurones sont
deux à
trois fois
plus nombreux
que ceux du
chimpanzé. |
|||||||||||||||||||||||||||||||||||
|
|
2.3 Mais l'homme n'est pas un singe |
|
|
||||||||||||||||||||||||||||||||||
|
Caractères de l'homme (anthropologie... biologique)
page sur l'identité biologique de l'homme |
|
|
|||||||||||||||||||||||||||||||||||
|
|
Du
point de vue
biologique
voici quelques
points qui
distinguent
l'homme des
autres
primates : « L'hominisation, c'est-à-dire l'introduction dans l'histoire de la vie du phénomène humain, n'est point l'apparition d'une espèce nouvelle, mais celle d'une forme nouvelle de la vie. L'enveloppe humaine de la biosphère est du même ordre de grandeur que la biosphère elle-même. L'humanité n'est pas une partie de la vie, mais l'équivalent, l'homologue de la vie. (...) Événement d'une portée considérable qui ne peut être comparé qu'à celui de l'apparition de la vie sur le globe. Vitalisation de la matière il y a plus de trois milliards d'années, l'hominisation de la vie il y a sans doute plus de deux millions d'années, tels sont les deux événements qui dominent l'histoire du Cosmos. » Jean Piveteau; article "hominisation" de l'Encyclopedia Universalis Une autre approche: quelques notes d'un séminaire de Michel Morange: L'HOMME ET LE SINGE, 17 Mars 2005: http://www.college-de-france.fr/media/phi_sci/UPL49356_Delmas17mars05.pdf |
|
Squelette d'un cours sur le site du laboratoire d'anthropobiologie de Toulouse: http://www. anthropobiologie. cict.fr/ Enseignements/ AgesdelaVie.pdf Homme
/ Autres
Primates et
Mammifères
: Ages de la
vie
très
différents la référence à l'ontogénèse et particulièrement aux hétérochronies est essentielle (voir ci-dessus et page de ce site sur les hétérochronies) |
||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||
|
Pour
le bac : |
|
L'homme
est le
chimpanzé
possèdent
des caryotypes
(description
du nombre, de
la forme et de
la coloration
en bandes des
chromosomes)
très
voisins:
24 paires de
chromosomes
pour le
chimpanzé,
23 pour
l'homme mais
le chromosome
2 de l'homme
semble
provenir de la
réunion
de deux
chromosomes du
chimpanzé,
13 paires de
chromosomes
ont exactement
la même
taille, forme
et coloration
de bandes chez
l'homme et le
chimpanzé,
7 chromosomes
présentent
des
différences
légères
de bandes. |
|
Les chimpanzés (et surtout les bonobos, Pan paniscus ou chimpanzés nains) utilisent fréquemment la bipédie, bien que principalement arboricoles. Les chimpanzés sont capables d'utiliser des outils (comme de nombreux autres animaux) : pour ouvrir une noix en la cassant entre deux pierres, pour éponger de l'eau retenue dans une cavité d'un tronc... |
|||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||
|
3. Les fossiles sont des traces des êtres vivants conservées dans les roches : les traces d'homme et de ses proches parents sont peu nombreuses et très discutées Voilà enfin le cœur du sujet, c'est par le contact avec les fossiles que les passions peuvent se développer... laissons les querelles de classification aux spécialistes... |
|
à conseiller: http://www.didac-tic.fr/ evolution/hominids_ history1.htm |
|||||||||||||||||||||||||||||||||||
|
L'espèce biologique ne coïncide pas avec l'espèce paléontologique |
|
|
|||||||||||||||||||||||||||||||||||
|
. |
|
Si l'espèce humaine (espèce biologique) est une (Homo sapiens), on a donné le nom de genre Homo à plusieurs espèces paléontologiques fossiles (Homo erectus, habilis, neanderthalensis... mais aussi sapiens), on aurait tendance actuellement à leur donner un autre nom de genre pour éviter la confusion avec la seule espèce humaine. Certains paléontologues préfèrent parler de la sous-espèce H. sapiens sapiens pour l'homme actuel, ce qui brouille encore davantage le discours. |
|
Une
espèce
paléontologique
est
définie
par des critères
morphologiques
ou
morpho-anatomiques
(par
comparaison
avec d'autres
fossiles et
avec des
espèces
actuelles)
mais aussi des
critères
paléogeographiques
ou
stratigraphiques,
c'est-à-dire
tous les
éléments
qui permettent
de
connaître
l'environnement
dans lequel
pouvait vivre
l'organisme
qui a fourni
le fossile. |
|||||||||||||||||||||||||||||||||
|
La même remarque peut être faite au sujet de l'outil: |
|
L'outil biologique (utilisé de façon incontestable par de nombreuses espèces animales actuelles) n'est pas l'outil paléontologique.
|
|
Qu'est-ce
qui
distinguerait
une branchette
utilisée
par un
chimpanzé
pour
récolter
des fourmis,
d'une autre
branchette ?
La nature
d'outil d'un
objet
paléontologique,
doit
être
prouvée
paléontologiquement,
c'est-à-dire
par des
preuves ou
indices
paléontologiques
(situation de
découverte,
association
avec d'autres
restes,
débit
et autres
traces
d'utilisation....).
Aucun outil
paléontologique
n'est
désormais
associé
aux
Australopithèques.
Les outils
paléontologiques
sont donc
désormais
réservés
au genre
-paléontologique
lui aussi -
Homo.
|
|||||||||||||||||||||||||||||||||
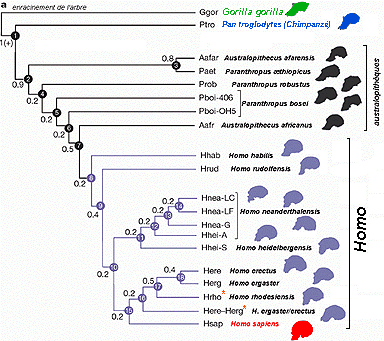
|
3 lignées : australopithèques, homo et sapiens sous-tribu
des hominina cladogramme récent de la lignée (page cladisme) |
|
Pour faire simple, et en suivant la proposition d'A. Dambricourt-Malassé reprise par J. Chaline, il existe 3 lignées distinctes (correspondant à de nombreux genres et espèces PALÉONTOLOGIQUES): Australopithèques (des formes robustes aux formes graciles), Homo (des formes habiles, érigées aux néandertaliens*, robustes) et enfin Sapiens (homme actuel ou moderne, qui pourrait être assimilé à l'espèce humaine biologique). * j'ai adopté l'orthographe le plus courante: Neanderthalensis prend un "h" alors que néandertalien ou Néandertal, s'écrit sans "h", mais je crois qu'on peut accepter les deux orthographes pour tous les dérivés du nom. <<<<-- pour placer les espèces dans le temps et l'espace voir par exemple une page du site de Tautavel à l'adresse: http://www.culture.gouv.fr/ culture/arcnat/ tautavel/fr/ hom_conq_pg.htm |
|||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||
|
Manuels
scolaires et
article Hominidés,
Brigitte
Senut,
Encyclopédia
Universalis,
v12, 2007 |
|
|
|
|
|
|
|||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||
|
Taille |
|
(H. neanderthalensis plus petits :1,5 à 1,7 m) |
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||
|
âge des fossiles
|
|
2 Ma à 1,5 Ma (H. habilis) 1,8 Ma à 0,15 Ma (H. erectus) 0,4 Ma - 0,03 Ma (H. neanderthalensis) |
(site de Qafzeh en Israël) depuis 400.000 ans (0,4 Ma) si l'on considère les formes les plus archaïques |
||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||
|
volume encéphalique |
|
800-1700 cm3 (H. erectus et neanderthalensis) |
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||
|
squelette |
premières traces de bipèdie incontestable (Lucy) mais un peu claudicante; déplacement (de type grimper) encore majoritairement arboricole |
bipèdie de type humain (mais pas de station érigée) |
bipédie humaine permanente et surtout station érigée ; articulation de la hanche (fémur-bassin) particulière ; position particulière des ligaments et des muscles qui les prolongent; col du fémur très allongé. |
||||||||||||||||||||||||||||||||||
|
remarques sur la bipédie et la station érigée |
La bipèdie est le mode de locomotion habituel de l'homme sur ses deux pieds (du latin bi = deux et pedes = pieds). Il ne faut pas confondre la bipèdie avec la station verticale, redressement du corps, et notamment la station érigée de l'homme. La bipèdie est considérée comme acquise dans de très nombreux groupes fossiles de façon indépendante: reptiles Thécodontes, nombreux dinosaures (Théropodes carnivores dont les fameux Tyrannosaures, Ornithopodes dont les Iguanodons herbivores). Actuellement certains lézards comme le Basilic pratiquent une course bipède active. Mais il suffit de penser aux Oiseaux pour retrouver des bipèdes. On n'oubliera pas les kangourous chez les mammifères marsupiaux actuels. |
|
La station érigée de l'homme a été récemment associée à une rotation du basi-sphénoïde à la base du crâne (avec une contraction crânio-faciale) résultant d'une flexion du tube neural au cours de l'embryogénèse (voir les travaux d'Anne Dambricourt-Malassé ci-dessus). |
||||||||||||||||||||||||||||||||||
|
|
|
|
|
||||||||||||||||||||||||||||||||||
|
industries et culture
Pour une belle réalisation multimédia sur les outils et les techniques voir le site de Tautavel: http://www.culture. gouv.fr/culture/ arcnat/ tautavel/fr/ comp_mat _acc_pg.htm, qui contient d'ailleurs plein d'autres trésors. |
On penche maintenant pour la thèse selon laquelle aucun australopithèque n'aurait fabriqué d'outil (les plus anciens sont datés de 2,5Ma). Aucun australopithèque ne semble avoir chassé même si l'on ne peut exclure qu'ils aient pu tuer de petits animaux. Ils avaient une nourriture coriace (attestée par les striations dentaires) principalement d'origine végétale (racines, baies dures...). Ils étaient la proies de prédateurs qui ont accumulé leurs ossements dans des grottes. |
Les chopper (= galets aménagés) sont les premiers outils incontestables. Les premières industries sont très stables dans les formes et continues dans le temps (on connaît des chopper depuis 2,5 Ma en Afrique jusque vers 0,2 Ma en Extrême-Orient. Les premiers outils aménagés sur les deux faces (bifaces) sont associés à H. erectus et datés de 1,6 Ma; à partir de 0,25 Ma les industries évoluent vite et se succèdent dans le temps. Les plus anciens outils en os ont été découverts à Swartkrans et Sterkfontein en Afrique du Sud dont certains ont été brûlés attestant l'usage du feu vers 1,5 Ma. Ces grottes karstiques sont d'une richesse incroyable mais posent des problèmes de datation (nouvelles méthodes U/Pb, LR, dossier 32 p 36). Les sépultures sont connues depuis 0,9 Ma Le feu est connu depuis 0,45 Ma mais ne semble s'être généralisé que vers 0,2 Ma |
Au Paléolithique moyen (entre 0,2 et 0,04 Ma) il existe encore des industries lithique importantes (industrie moustérienne et technique Levallois...). À partir du Paléolithique supérieur (0,04 à 0,016 Ma) les outils sont extrêmement variés (industrie magdalénienne). Le Néolithique (normalement placé en France entre 5.000 et 2.000 ans avant J.C.) est encore appelé "âge de la pierre polie". Les premiers mégalithes en Europe datent d'environ 6.000 ans alors que les premières villes apparaîtraient au Moyen-Orient vers 8.000 ans. Les plus anciennes traces d'agriculture sont rapportées au croissant fertile (Irak, Syrie, Turquie) vers 7.000 ans. Le cuivre travaillé ferait son apparition vers 8.000 ans en Orient et 5.000 en Europe. Le bronze suivrait vers 5.000 ans et le fer vers 2.000 ans. L'écriture cunéiforme daterait de 3.300 ans. La plupart des hommes "modernes" trouvés récemment au Proche-Orient et datés de plus de 100.000 ans sont assez bien conservés car ils proviennent de sépultures. |
||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||
|
localisation Dossier LR 32, 2008, pp 64-68; |
|
|
(depuis Israël mais berceau africain très discuté pour des "hommes modernes archaïques") |
||||||||||||||||||||||||||||||||||
|
genres et espèces PALÉONTOLOGIQUES
E.U.
article "Australopithèques",
José
Braga, (1) voir aussi Les premiers hommes hors d'Afrique, dossier La Recherche, 419, mai 2008, 28-42 et Dossier LR, 32, 2008, pp 58-62; Les outils trouvés sur le site (plus de 5.000) sont plutôt de culture oldowayenne (éclats, chopper, galets aménagés sur une face...) connue en Afrique 1 Ma auparavant. La faune fossile retrouvée (mammouth archaïque, chevaux, rhinocéros, daims, chiens, hyènes) est européenne, malgré quelques spécimens (tigre à dents de sabre, girafoïde du genre Paleotragus ou l'autruche géante...) qui sont aussi africains. La datation biostratigraphique est assez fiable. (2) les paléontologues chinois ont entrepris depuis la fin du XXème une recherche systématique de traces d'hominidés en Chine; 3 dents de très grande taille, datées de 2 Ma ont été retrouvées dans le comté de Jianshi (province du Hubei) associées à de très nombreux outils de type chopper; elles ont été attribuées à un genre proche d'H. erectus : Meganthropus. (Dossier LR n°32, p 47-49) (3)
À
écouter
: Un intéressant entretien sur Canal Académie (radio de l'Académie des Sciences morales et politiques): Les « migrations » des hommes préhistoriques, avec Henry de Lumley (52 min, peut être téléchargé : 35,8Mo, mp3)
Dossier La Recherche: La nouvelle histoire de l'homme, août 2008, 32 |
1 ou 2 (voire 3) genres, tous fossiles: * Australopithecus africanus , Afrique du Sud (sites de Makapansgat, Gladysvale, Sterkfontein (1947) et Taung (1ère espèce décrite: 1925, c'est le fossile de référence du genre) - 3,2 - 2,8 Ma Australopithecus barelghazali, Tchad (site de Koro Toro - 2000 km des sites éthiopiens, 1995; une mandibule) - 3 - 3,5 Ma Australopithecus garhi, Éthiopie (Bouri, 1999), 2,5 Ma Deux espèces sont très discutées : Australopithecus anamensis, Kenya (sites de Kanapoi et d'Allia Bay 1995), 4,4 - 3,2 Ma * Australopithecus afarensis, Éthiopie (sites de Belohdelie, Maka et Hadar (Lucy, 1974)) et Tanzanie (sites de Laetoli et Garusi), 4,4 - 2,6 Ma Les trois espèces qui suivent sont parfois (notamment par B. Senut) rattachées au genre Paranthropus nez plat, grande face légèrement concave avec orbites très marquées formant un rebord, front bas et légèrement creux, grandes molaires et prémolaires avec cuspides plates, grosses mâchoires massives, incisives et canines assez petites, crête sagittale au niveau du crâne permettant l'insertion de muscles masticateurs puissants (nourriture coriace ?) Australopithecus aethiopicus, Éthiopie (vallée de l'Omo, 1967) et Kenya (à l'ouest du lac Turkana, 1995), 2,5 Ma Australopithecus boisei (hyperrobuste), Éthiopie (vallée de l'Omo et Konso), Kenya (à l'est du lac Turkana) et Tanzanie (à Olduvai, Zinjanthrope, 1959), 2,2 - 1,2 Ma Australopithecus robustus, Malawi (site d'Uraha) et Afrique du Sud (sites de Kromdraai, Drimolen et Swartkrans) - 2 Ma
Ce genre n'est rattaché aux australopithèques que par certains auteurs (voir ci-dessous : hominidés du Miocène terminal) Ardipithecus ramidus, Éthiopie (région de Gona), 4,51-4,32 Ma (2005) ; un squelette assez complet bien que fragmenté en 135 morceaux; l'équipe de Tim White propose (2009) de rattacher les 3 hominidés ci-dessous à ce fossile pour faire un unique genre qui serait le premier hominidé doué d'une bipèdie "archaïque"; cette hypothèse reste très controversée. |
Le genre Homo a d'abord été défini à partir de fossiles regroupés maintenant dans une espèce robuste : * Homo erectus, Kenya (Nariokotome, garçon de Turkana), dès 1,7 Ma, Tanzanie (Olduvai), Ouganda (Nyabusosi), Éthiopie (Bouri, calotte crânienne, 2002, 0,8-1 Ma), Afrique du Sud (Swartkrans), Afrique du Nord (homme de Rabat, Algérie (Ternifine), vers 500.000 ans), Tanzanie (bords du lac Ndutu) et Éthiopie vers 400.000 ans. Chine (Zhoukoudian, 500.000 ans); Java (Pithécanthrope, 1891, 1,7 à 0,8 Ma...), Israël (Oubeideiya, 1,4 - 0,7 Ma), Syrie (0,55 Ma), Allemagne (Mauer, 0,7 Ma, H. heidelbergensis), France (Tautavel, Caune de l'Arago, 0,45 Ma; crâne, 120 ossements, nombreux avec stries de découpage (cannibalisme), feu inconnu... voir note 3) Hongrie (Vértesszöllös, 0,3 Ma), Chine (2 Ma (voir note 2) ; 0,4 -0,2 Ma) ; encéphale assez réduit/habilis, voûte crânienne plus basse, bourrelet sus-orbitaire assez développé, couches externes et internes des os crâniens épaisses, os du squelette des membre avec couche externe épaisse, fémurs et tibias aplatis, bipédie adaptée à de longs trajets. Une
espèce
créée
en 1975 qui
aurait
cohabité
avec H.
erectus au
Kenya:
Les formes primitives, moins robustes qu'erectus, sont rattachées à l'espèce habilis; certains voudraient les classer dans des genres (et donc des clades) différents (sans parenté directe) Homo habilis, Tanzanie (Olduvai, une mandibule, 2 pariétaux, des dos des mains et un pied presque complet, 1960), Kenya (est du lac Turkana), Éthiopie, Afrique du Sud, 1,8 Ma Homo rudolfensis, nouvelle dénomination des pièces du Kenya anciennement rattachées à H. habilis, Malawi (Uraha), 2,5 Ma La mandibule de Dmanisi (Géorgie) trouvée en 1991 a été suivie peu après par des crânes quasi complets trouvés non loin. D'autres pièces, venant des membres et de la colonne vertébrale, se sont ajoutées. Les squelettes (1 adolescent et 3 adultes ?) présentent une mosaïque de caractères à la fois primitifs (petite taille, faible capacité crânienne, absence de torsion humérale: rattachés à H. habilis ?) et évolués (proportions du squelette, morphologie des membres inférieurs indiquant la possibilité de longs trajets... rattachés à H. erectus, principalement à partir des caractéristiques du garçon de Turkana). Ils sont datés de 1,77 Ma ce qui pourrait faire penser que des espèces africaines d'H. habilis aient pu directement venir d'Afrique. En tout cas ces fossiles, qui correspondent aux plus vieux Homo européens, ne possèdent pas l'ensemble des traits évolués caractéristiques des H. erectus africains (voir note1). Homo
antecessor,
Espagne (Gran
Dolina,
sierra Atapuerca,
1994), 0,78 Ma Deux espèces les plus anciennes mais définies avec un seul fossile chacune: Praeanthropus africanus, Tanzanie (fragment maxillaire rattaché à A. anamensis, 1930 réinterprété 1995), Kenyanthropus platyops, Kenya (lac Turkana, Lomekwi, un crâne écrasé), 3,2-3,5 Ma La troisième espèce d'Homo n'est jamais africaine: Homo neanderthalensis voir aussi colonne suivante ----------> les traits néandertaliens apparaissent indépendamment sur des fossiles présentant des traits d'H. erectus en Europe (caune de l'Arago, Tautavel) vers 0,4 Ma ; ils sont assez typiques à partir de 0,25Ma sur de nombreux fossiles européens (Biache-Saint-Vaast, Pas-de-Calais ; Saccopastore, banlieue de Rome); enfin, à partir de 0,125 Ma les caractéristiques néandertaliennes sont pleinement développées en Europe et la population néandertalienne atteint son maximum d'extension géographique en Asie et au Moyen-Orient (débutée vers 0,2 Ma) : crâne de Tabun I (Mont Carmel, Israël), 0,175 Ma. Un inclassable: Homo floresiensis, Indonésie (île de Flores), 18.000 ans, 7 individus, un squelette presque complet... une nouvelle espèce, un descendant de l'H. erectus ? un nain d'H. sapiens ? |
Homo sapiens sapiens ou Sapiens, "homme moderne", les plus anciens indiscutables : Israël (site de Qafzeh), 100 000 ans, Afrique du Sud (site de Klasie) 90.000 ans. Des formes archaïques (fort développement encéphalique associé à de fortes superstructures crâniennes) très voisines d'H. erectus. Tanzanie (Ndutu), 400.000 ans), Afrique du Sud (Klasies River Mouth, 80,000 ans). Depuis 1998 nombreuses découvertes en Afrique. Namibie (Oranjemund, 1998, calotte crânienne, 100.000-50.000 ans), Éthiopie (hommes de Herto, découverte 1997, publication 2003, 160.000-154.000 ans ; vallée de l'Omo, formation de Kibish, 2 crânes découverts en 1967, datés à nouveau en 2005 de 198.000-104.000 ans).
Pour certains auteurs ces formes archaïques constituent les premiers éléments d'une seule lignée qui donnera soit des Sapiens (H. sapiens sapiens) soit des néandertaliens (H. sapiens neanderthalensis). Pour d'autres, comme je l'ai présenté ici, avec 2 colonnes, il y a deux lignées distinctes : les néandertaliens (H. neanderthalensis), forme évoluée des H. erectus, et les Sapiens qui forment une espèce à part (ou même un genre et plusieurs espèces paléontologiques si l'on considère des formes archaïques) qui prendrait son origine en Afrique vers 400.000 ans (pour des données anatomiques et embryologiques voir le travail d'Anne-Dambricourt-Malassé ci-dessus ...) . En tout cas, tous les auteurs s'accordent maintenant pour parler de coexistence entre les deux espèces et le scénario le plus souvent proposé POUR L'EUROPE est celui du remplacement d'une espèce (néandertalien) par une autre (Sapiens), venue de l'Est. Mais les preuves d'interactions entre les deux populations manquent. |
||||||||||||||||||||||||||||||||||
| |
|
|
|||||||||||||||||||||||||||||||||||
|
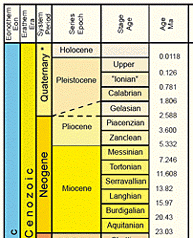
Des
hominidés
du
Miocène
terminal échelle
stratigraphique:
http://www.stratigraphy.org/cheu.pdf, page sur le climat du Quaternaire |
|
Ces fossiles sont très discutés. Certains les considèrent comme faisant partie des hominina (voir cladogramme), d'autres les rattachent aux grands singes (chimpanzés : sous-tribu des panina). Ils datent de la fin du Miocène, et ont été trouvés en Afrique. Ces fossiles sont peu nombreux et très fragmentaires. Dossier La Recherche: La nouvelle histoire de l'homme, août 2008, 32 Une idée particulièrement intéressante dans l'article de Robert Martin : « Plus on se rapproche de la divergence des hominidés et des chimpanzés, plus les fossiles des deux lignées se ressemblent. Les classer correctement nécessite des analyses attentives. En particulier, ces études sont souvent victimes d'un biais systématique : on considère que tous traits par lesquels les grands singes africains actuels différent des humains sont primitifs. Cette tendance à traiter les chimpanzés et les gorilles comme des 'fossiles vivants' peut nous induire en erreur : pour chaque caractère nous devons établir la condition [l'état] primitive [f] par une analyse détaillée. Ainsi les singes africains ont un émail dentaire fin, tandis que les hommes ont un émail épais. On a souvent considéré que l'émail fin, caractéristique des grands singes africains, était primitif, et qu'un émail dentaire épais, comme le nôtre, indiquait qu'un fossile donné faisait partie de l'évolution des hominidés. Toutefois, les orangs-outans (dont la branche a divergé avant celle des grands singes africains) ont un émail dentaire modérément épais. Nous devrions donc au moins envisager la possibilité que l'ancêtre commun des chimpanzés et des hommes avait [ait eu] un émail assez épais.» |
|
Ardipithecus kadabba, 5,2-5,8 Ma, Éthiopie (Middle Awash), 1997, fragments mandibule, dents, fragments squelette membres ; les 17 fragments rattachés à ce nouveau genre ont permis de mettre en évidence un foramen magnum en position relativement antérieure, ce qui est un indice qui pourrait faire penser à une stature érigée, mais la courbure des phalanges suggère plutôt à des doigts préhensiles intervenant dans une locomotion arboricole par grimper et suspension. De même les fragments du crâne le rapprochent plutôt du chimpanzé, tout comme les dents. Orrorin tugenensis, 5,8-6 Ma, Kenya, 2000, fragments mandibule, dents, fragments squelette membres; serait le plus ancien bipède connu (col du fémur allongé, et dont la face arrière présente un sillon qui pourrait être la trace d'attachement du muscle locomoteur : obturator externus, caractéristique exceptionnelle chez les grands singes (comme les gorilles) mais répandue chez tous les hominidés récents) mais on a des incertitudes sur les conclusions faites à partir de fragments assez abîmés. Les fragments d'humérus et de phalange indiquent plutôt une adaptation au grimper. Sahelanthropus tchadensis, 6-7 Ma, Tchad, 2001, crâne "Toumaï ", fragments de mâchoires et dents isolés sur le même site; le crâne est presque complet mais écrasé et tordu; foramen magnum en position relativement centrale avec muscles d'attache en position assez verticale, ce qui indique une stature érigée de l'axe céphalique-colonne vertébrale, indice d'une possible locomotion bipède. Ce crâne présente à la fois des caractères proches des australopithèques et des caractères partagés avec les chimpanzés (faible volume, arcades sourcilières proéminentes...). |
|||||||||||||||||||||||||||||||||
| |
|
|
|||||||||||||||||||||||||||||||||||
|
Dossier LR, 32, 2008, 85 |
|
Ces traits sont réunis ici dans un tableau mais ils ne correspondent pas à un fossile complet et typologique. Bien que les données morphologiques soient plus accessibles, il ne faut pas oublier l'aspect embryologique (complexité de l'origine des os du crâne, voir par exemple l'annexe de l'ancienne page sur les modèles et le travail d'Anne Dambricourt-Malassé référencé ci-dessus). |
|||||||||||||||||||||||||||||||||||
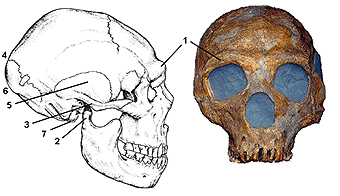
|
Comparaison
TRÈS
CRITICABLE des
silhouettes
squelettiques
entre un
néandertalien
et un sapiens
(origine
inconnue)
supposés
caractéristiques. |
|
|
|||||||||||||||||||||||||||||||||||
| |
|
|
|||||||||||||||||||||||||||||||||||



{kind=link}