À la recherche de marqueurs de l'identité biologique de l'homme
|
|
|
|
|
|
|
||
|
Une question d'anthropologie |
|
Le problème de l'identité n'est bien sûr pas biologique mais plus largement anthropologique; il sort même du domaine de l'anthropologie biologique; il atteint le fondement de notre personne (ce que nombre de philosophes ont appelé notre âme, représentée par notre conscience et notre volonté LIBRE). Pour aborder la question d'une autre manière, voici deux pistes: - écouter une émission grand-public de Canal Académie: Qu'est-ce que l'identité ? par le sociologue Jean-Claude Kaufmann http://www.canalacademie.com/ L-identite.html (sur le même média, l'interview de Michel Serres (Corps et identité) n'apporte pas grand chose, étant donné l'attachement de l'auteur à la définition d'une identité basée sur la séquence de l'ADN, ce qui est - un rêve - fortement en désaccord avec les résultats de la génétique - voir ci-dessous, les marqueurs génétiques). Le livre d'Edgardo Carosella et de Thomas Pradeu: "L'identité : la part de l'autre" repose sur la même vision matérialiste même si un niveau supérieur de contrôle est ajouté. - lire le texte d'un académicien (section Biologie humaine et Sciences Médicales): Pierre KARLI : Peut-on parler d'une biologie de la personne humaine ? (dans le n°1 de PLASTIR : http://plasticites-sciences-arts.org/PLASTIR/Karli.pdf) |
|
Mais la certitude que nous sommes une personne unique, différente de toutes les autres, n'est pas forcément inexprimable par la biologie, sans que cette science présente une approche exhaustive (la transcendance de l'homme à laquelle je crois ne sera jamais un résultat de la science). La vision biologique, nécessairement limitée, n'en est pas moins vraie.
|
|||
| |
|
|
|||||
|
Plan
en travaux |
|
Introduction:
une
nature humaine
partagée
... et une
personne,
unique,
douée
d'une
volonté
et d'une
conscience
propres,
à
nulles autres
pareilles, et
avec un corps
mortel |
|||||
|
But de cette page : une fois écrite, il est apparu clairement que mes propos ci-dessous visent à expliquer en quoi la vision de l'identité biologique de l'homme que j'ai trouvé dans les ouvrages scolaires et dans nombre de médias de diffusion-éducation scientifique me semble erronée: l'homme ne possède pas de carte d'identité génétique, moléculaire ou biochimique. La biochimie relie tous les êtres vivants. Ce qui différencie les êtres vivants, ce ne sont pas les molécules, mais les formes dynamiques, qui synthétisent, renouvellent et dégradent les molécules et les structures du vivant. Ainsi, l'homme présente une forme individuelle dont les caractéristiques lui sont propres et peuvent être explorées par différentes techniques. |
|||||||
|
Qu'est-ce qu'un homme ? |
|
Une nature humaine partagée |
|||||
|
L'homme ne se définit pas en opposition avec autre chose mais il est sans aucun doute proche des animaux par bien des côtés, tout en s'en différenciant de façon radicale. L'anthropologie parle de nature humaine ou d'humanité qu'elle différencie de la nature animale ou animalité |
|
Une
réponse
anthropologique
non
scientifique:
Une
réponse
scientifique: |
|
Pour amorcer un débat qui dépasse le cadre de ce cours mais qui est nécessaire dans l'optique de la classe de terminale et moins scolairement pour tout élève curieux, je conseille l'audition de la conférence d'Élisabeth Fontenay , maître de conférence émérite en philosophie à Paris I, dans le cadre de l'éthique à l'ENS accessible sur internet : De l'animal à l'humain, du 13 avril 2005 (accès direct fichier audio mp3 - 17,6Mo) ; son propos, qui se présente comme une négociation pour éviter des positions rigidifiées, est de montrer l'enrichissement de la conception de l'animalité moderne par rapport à une séparation nette des natures (elle cite Plutarque qui utilise le mot philanthropie dans le sens d'une bienveillance de l'homme envers les animaux et non pas seulement envers les hommes). L'animalité commune à l'homme et aux animaux, et qui fonde, à son avis, une nécessaire éthique animale à base d'empathie, n'empêche pas que l'on pose, comme principe non négociable, la profonde différence (philosophique, politique et historique selon ses propos) de l'homme et de l'animal (extrait mp3 sur ce site, 1,3Mo). Je conseille aussi le débat qui a suivi dans lequel sont soulevés deux problèmes majeurs: le caractère carnivore de l'homme (une étape à dépasser ? ou à redécouvrir: manger non pas de la viande mais un animal, que l'on respecte, voir conférence de Marylène Pathou-Mathis sur la consommation de viande dans la préhistoire) et la position d'une barrière spécifique au niveau de la phylogénie. Même si je ptéfère le débat ci-dessus, on peut aussi noter que de nombreux scientifiques pensent que la différence homme-animal peut être posée scientifiquement. Par exemple, j'ai tenté de suivre le débat entre Alain Prochiantz, neurologue (un brillant matérialiste athée qui défend une rupture scientifique entre l'homme et l'animal) et Dominique Lestel, ethnologue (partisan d'une nouvelle "religion" (sans Dieu !).. je ne suis pas sûr d'avoir tout saisi..., matérialiste où l'homme et l'animal sont réunis): http://www.canal- u.com:80/ canalu/ chainev2/ utls/programme/ 115535685_ y_a_t_il_ une_barriere_entre_l_ homme_et_l_animal_/. |
|
|
|
|
|
|
||||||
|
Tous parents ... |
|
Ce qui nous rapproche des autres : l'espèce humaine |
|
|
|||
|
* il va sans dire que les personnes qui seraient nés d'une fécondation in vitro sont aussi passés par le l'utérus maternel, d'où le problème fondamental des mères porteuses. Pour l'instant, et grâce à Dieu, les développements en milieu artificiel (extra-utero) s'arrêtent au milieu (10 jours sur les 20 jours de gestation chez le souris...). Qu'est-ce
qu'une
espèce? |
|
+ notre origine humaine ou parenté (nous sommes nés d'une femme)... ce que la biologie exprime par la notion d'espèce : nous appartenons à l'espèce humaine à partir du moment où l'on est né d'une femme* (mais on voit bien que la biologie devient impuissante à donner des critères définitifs...car notre mère a eu aussi une mère... et l'on remonte ainsi à l'origine de l'espèce humaine, à la spéciation, l'apparition d'une espèce, à partir d'une autre espèce par transformation, selon le concept de l'évolution (quelle que soit la théorie à laquelle on se réfère). On notera que si l'on est tous parents c'est bien au regard de l'évolution et non au regard de la société. C'est une parenté biologique et non familiale. Une première conséquence est que, du fait même que le fondement de notre première identité est formé par notre mère et notre père, la famille, comme lieu de conception puis de naissance et enfin d'éducation, revêt une importance singulière, du strict point de vue biologique. Changer ce rapport, changer la structure familiale, c'est proposer un changement de l'identité biologique humaine. Ces éléments font partie de l'anthropologie biologique. |
|
Une deuxième conséquence de cette identité de naissance (génétique au sens étymologique, c'est-à-dire celui de la parenté), est que l'étude des caractéristiques qui nous relient à notre espèce tombe dans le domaine de la phylogenèse: ce sont les études phylogénétiques qui nous permettent de trouver les relations logiques entre les caractères qui nous définissent entre nous et par rapport aux autres espèces. C'est ainsi que le cladisme, méthode phylogénétique majoritairement employée, est devenu le moyen presque unique d'étudier nos spécificités. Les caractères matériels (biochimiques, physiologiques, anatomiques...) ou immatériels (comportementaux, culturels...) sont ainsi évalués et comparés en fonction de leur pertinence à nous définir comme espèce originale. Les caractères génétiques ont parfois pris une importance exagérée dans ces phylogénies, nous y reviendrons. |
|||
|
|
|
Mais il est aussi clair que si notre naissance nous rapproche des autres il nous en éloigne aussi puisque, les liens de "sang" (ou plutôt de "parenté") nous font partager la même mère (et généralement le même père) avec nos frères et sœurs, au sein d'une fratrie, mais pas avec des individus éloignés avec lesquels nous n'avons aucun lien familial actuel apparent. La mode des généalogies met en évidence ce besoin de reconstituer un "clan" élargi, avec des liens de parenté plus lâches que dans la famille stricte, mais qui remonte le plus loin possible (on se souvient que dans le judaïsme, il est d'usage de conserver sa généalogie qui remonte si possible à Abraham, père du peuple élu. Ces généalogies étaient aussi bien établies selon la génération par les hommes que par les femmes). Dans la plupart des scénarios évolutifs, la spéciation oblige à remonter à un couple fondateur unique, ce qui rejoint la foi de nombreux hommes religieux qui croient en un couple originel à l'origine de l'humanité (Adam et Ève). |
|||||
|
|
+ notre corps, notre matérialité (non pas au sens de matière aristotélicienne, pure puissance, mais comme composé sensible), ce que la biologie exprime par la biochimie mais aussi l'anatomie, la physiologie... |
|
Remarque: |
|
C'est aussi ce qui fonde les possibilités de greffes entre individus de la même espèce. Avec une identité plus large, par exemple notre appartenance au groupe des mammifères, les possibilités de greffes sont encore davantage étendues mais réussissent plus rarement; on parle de barrière spécifique pour désigner cette limitation des processus à l'espèce. |
||
| |
|
|
|
voir
Canguilhem: |
|||
| |
|
Une personne, unique, douée d'une volonté et d'une conscience propres, à nulles autres pareilles, et avec un corps mortel |
|||||
|
... tous différents |
|
Ce qui nous différencie des autres : |
|
|
|||
|
voir
Canguilhem: |
|
+ des particularités individuelles physiques qui peuvent être prises en compte par des mesures morpho-anatomiques, biochimiques, physiologiques.... ... mais il ne faut pas oublier que la plupart des sciences expérimentales cherchent à s'affranchir de l'individu pour atteindre le général. Il n'y a guère de science de l'individu... Il y a une sorte de répugnance de tout scientifique à considérer l'individuel. C'est une question essentielle de nos jours où l'on essaye de mettre en place des modèles reposant sur l'émergence et l'autonomie. Il y a alors une redécouverte de l'individu. + nos choix libres... |
|
Une
des
réflexions
actuelles les
plus
pertinentes
sur ce sujet
nous est
donnée
par Nissim
Amzallag: Du
sens de la
variabilité
in Génétiquement
indéterminé
- le vivant
auto-organisé,
2007,
éditions
Quæ
(Cemagref,
Cirad,
Ifremer,
Inra). |
|||
| |
|
Remarque: les molécules sont communes aux êtres vivants même si on trouve des spécificités dans tel ou tel groupe (cellulose chez les plantes, certaines bactéries; chitine chez les arthropodes, les mycètes...; tout en sachant que ces molécules sont des types qui présentent de nombreuses variantes chimiques). L'idée selon laquelle c'est la combinaison originale des différentes molécules qui caractérise l'individu recèle la même erreur. Car il faut alors chercher un support à l'individualité qui dirigerait la synthèse et l'ordonnancement de ces molécules. Et l'on revient ainsi à la notion d'information (le plus souvent génétique). Or cette information manque. L'information génétique trouvée (un gène du protéome) est au contraire une unité fonctionnelle stable et plus ou moins transmissible. Le reste c'est de l'imagination. |
|
|
|||
|
|
|||||||
|
De
l'information
à la
forme |
|
À l'instar de la carte d'identité individuelle, issue de l'occupation -lors de la dernière guerre - en France, on a cru que le concept d'information, et principalement d'information génétique, pourrait nous mener à une identification de la personne. |
|
Maintenant plus personne ne croît à cette notion d'information (l'information génétique n'est qu'une information pour une séquence d'une molécule). Les réseaux d'interactions que l'on invente ne sont pas stockés dans l'ADN. On retourne donc progressivement vers ce qui était le point de départ de la définition de l'identité biologique: la mesure des formes: la biométrie ou plutôt la morphométrie. |
|
Ainsi les résultats des techniques de caryotypage ou du polymorphisme de longueur des séquences répétitives de l'ADN ne sont plus considérés comme une information qui serait notre propriété et qui nous définirait, mais bien comme la forme - individuelle- que prend l'ensemble de nos fonctions vivantes dans une espace de grande dimension. (voir Qu'est-ce que la vie ? et la modélisation avec René Thom) |
|
|
|
|||||||
|
1. l'ADN, les chromosomes et les marqueurs génétiques |
|
en travaux |
|||||
| |
|
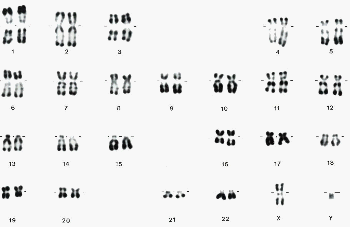
1.1 Les chromosomes disposés en caryotype sont utilisés comme marqueurs de l'espèce |
|
|
|||
|
|
|
||||||
| |
|
|
Les chromosomes ne sont pas des pelotes d'ADN : leur formation, leur disparition et surtout leurs rôles restent bien mystérieux |
||||
|
|
|
Les chromosomes sont de gros "corps colorables" allongés (de quelques micromètres de long pour 0,7 µm de diamètre de chromatide) qui apparaissent dans le noyau des cellules eucaryotes lors de la division cellulaire. |
|
On appelle caryotype, d'une cellule, le classement des chromosomes colorés, fixés et étalés, obtenus par blocage de la division au stade de la métaphase. |
|
Le nombre et la position des bandes colorées/non colorées (dont on ignore l'origine mais que l'on sait obtenir de façon reproductible avec une technique standardisée) qui alternent sur les chromosomes dépendent de la technique de coloration (voir cours de seconde: les chromosomes). |
|
| |
|
|
|
le caryotype est stable au sein de l'espèce |
|||
|
voir le chapitre 1.4 Les variations caryotypiques dans la page sur les mutations |
|
Tous les individus d'une même espèce présentent le même caryotype (même nombre, même taille, même forme et mêmes bandes des chromosomes), sauf anomalie (et variabilité dans le cadre d'un polymorphisme plus ou moins héréditaire, voir ci-dessous)). Les anomalies sont RARES car on considère qu'elles sont la marque (et non à l'origine) de phénomènes gravissimes. |
|
Les
anomalies
caryotypiques
(chromosomiques)
peuvent
être des C'est au nom de cette affirmation que l'on compare les caryotypes des primates entre eux . Par exemple il y a 24 paires de chromosomes pour le chimpanzé et 23 pour l'homme. Mais le chromosome 2 de l'homme semble pouvoir provenir de la réunion de deux chromosomes du chimpanzé. 13 paires de chromosomes ont exactement la même taille, forme et coloration de bandes chez l'homme et le chimpanzé et 7 chromosomes présentent des différences légères de bandes. |
|||
|
Une référence (accès restreint): Cytogénétique humaine. De 1956 à 2006, R. Berger, Pathologie Biologie 55 (2007) 1-12 |
|
« L'obtention des bandes pose des questions diverses. Une première est de définir le caryotype normal. On s'est vite aperçu en effet que certaines régions chromosomiques, en particulier celles qui contenaient l'hétérochromatine constitutive et les bras courts des chromosomes acrocentriques (13 à 15 et 21-22) pouvaient varier d'un individu à un autre, définissant un polymorphisme chromosomique. Ces polymorphismes se retrouvent dans une même famille et sont transmis selon le mode dominant mendélien [voir plus bas]. Les frontières entre polymorphisme et anomalie ne sont pas toujours aisées à établir, notamment dans le cas des inversions partielles dont les plus fréquentes concernent un chromosome 9, ou le bras court d'un chromosome acrocentrique qui pourraient être confondus avec le résultat d'une translocation à ce niveau. L'intérêt des polymorphismes chromosomiques est qu'ils peuvent servir (et ont servi) de marqueurs génétiques pour suivre des ségrégations de chromosomes, identifier l'origine paternelle ou maternelle d'une anomalie, et définir le stade auquel a pu se produire une non-disjonction au cours de la méiose. Le polymorphisme chromosomique peut également être utilisé pour identifier rapidement une lignée cellulaire par caryotype en flux, permettant ainsi de détecter rapidement une contamination de lignées cellulaires cultivées au long cours, accident trop souvent ignoré dans les laboratoires. Une deuxième série de questions posée par l'existence de polymorphisme chromosomique est sa responsabilité éventuelle dans la survenue d'anomalies chromosomiques constitutionnelles ou acquises en particulier des non-disjonctions ou même des remaniements de structure liés par exemple à des segments d'hétérochromatine de taille inhabituelle. Malgré de nombreux travaux, le rôle de polymorphismes chromosomiques dans l'apparition d'anomalies constitutionnelles, même à titre de prédisposition, n'a pu être prouvé de façon formelle ». (Berger, 2007) |
|||||
|
corollaire
: |
|
des différences caryotypiques peuvent être la marque de dérèglements du développement |
|||||
| |
|
Les chromosomes sont des organites spécifiques de la division des cellules eucaryotes. Ils sont devenus un moyen commode de détecter des maladies graves DU DÉVELOPPEMENT qui sont associées à des anomalies chromosomiques depuis le travail fondateur du professeur Lejeune sur le trisomie 21 (voir ancienne page). Mais il ne faudrait pas faire le raisonnement simpliste, issu du paradigme moléculaire (voir paragraphe suivant sur l'ADN), selon lequel les anomalies chromosomiques et partant, les chromosomes habituels, seraient À L'ORIGINE du fonctionnement plus ou moins déréglé de l'organisme. Plusieurs observations tendent au contraire à nous indiquer une direction bien différente et plus biologique que biochimique: des maladies transmissibles affectent le caryotype de certaines cellules de certains organismes. |
|
Les
résultats
les plus
probants nous
viennent de
cellules
cancéreuses.
Mais
très
peu de
recherches ont
été
réalisées
dans le but de
démontrer
la
variabilité
caryotypique
au sein de
l'organisme en
fonction des
types
cellulaires et
du
métabolisme,
ou encore du
développement. |
|||
|
ancienne page sur les trisomies
*trisomie en mosaïque = ne touche que quelques cellules ou quelques organes et non l'organisme entier ** disomie = anomalie chromosomique correspondant à la réunion de deux chromosomes paternels ou maternels dans une cellule, à la place d'une paire d'homologues (un paternel, l'autre maternel).
photos
de la
technique
d'amniocentèse
avec culture
des cellules
fœtales:
Une référence (accès restreint): Cytogénétique humaine. De 1956 à 2006, R. Berger, Pathologie Biologie 55 (2007) 1-12 |
|
Les trisomies sont principalement des anomalies du développement : Il est malheureusement assez rare de présenter une trisomie (et particulièrement la trisomie 21) comme une anomalie de développement et pourtant c'est ce qu'elle est. À mon sens l'anomalie chromosomique est la marque de l'anomalie du développement et non l'inverse (ce qui est une évidence si l'on raisonne au niveau cellulaire mais qui est plus difficile à formaliser au niveau de l'individu, faute de modèle). La preuve indirecte la plus flagrante vient de ce que l'on parvient à des traitements très efficaces en adaptant l'éducation et l'aide au développement psychomoteur de ces enfants. Cette anomalie est suffisamment précoce pour impliquer la quasi-totalité de l'embryon. Le fait que l'on puisse la détecter précocement chez des cellules fœtales (par caryotypage) montre que ces cellules ont déjà acquis un caryotype anormal et donc qu'elles ont hérité d'un dysfonctionnement. Il est fréquent de dire que ce caryotype est hérité de l'ovocyte ou du spermatozoïde comme en témoignent les exercices scolaires sur l'origine de la triploïdie lors d'une méïose anormale. Les marqueurs génétiques utilisés pour tracer l'origine paternelle ou maternelle (et méïotique ou mitotique) de la trisomie ne font que reprendre cette hypothèse sans la remettre en question. J'ai vainement cherché dans la littérature un caryotypage d'embryons au stade morula ou blastula (rejetés ou morts) reporté comme étant trisomique. À ma connaissance (et je serai très intéressé par des données sur cette question : pierre.stouff at libertysurf.fr) on ne rapporte pas de caryotype trisomique chez un embryon avant le développement du cœur (30-40% des trisomiques 21 ont une malformation cardiaque) et même du cerveau à 5 cavités (le retard mental est un trait omniprésent et il est difficile de ne pas placer son origine dans le cerveau) c'est-à-dire entre la 3ème et la 5ème semaine de développement. D'autre part, le relativement grand nombre de cas de trisomies placentaires (alors que l'embryon présente un caryotype normal - ce que l'on interprète comme des trisomies en mosaïques*, et que l'on rapporte le plus souvent à des disomies** plus ou moins corrigées-) tend à nous indiquer que l'on généralise peut-être bien hâtivement, dans notre enseignement, des phénomènes infiniment variés et complexes. Une autre manière d'aborder le problème vient des travaux récents réalisés avec l'idée que la trisomie est une maladie de dosage génique (50% de matériel génétique en trop pour une trisomie complète) et non pas une anomalie génétique. Or « les premiers résultats de l'emploi des puces d'expression paraissent montrer que la situation est plus complexe puisque, dans des cultures de fibroblastes de cellules foetales trisomiques 21 (comme de cellules trisomiques 13), le niveau moyen d'expression des gènes est augmenté seulement de 1,1 fois au lieu de 1,5 par rapport à des cellules normales. De plus, la majorité des gènes dont l'expression est dérégulée, ne sont pas localisés sur les chromosomes trisomiques. Ces résultats suggèrent que la trisomie constitutionnelle met en jeu un système complexe de régulation de l'expression génique et il est évident que d'autres études du transcriptome sont nécessaires.» (in Berger, 2007) Remarque: |
|||||
|
|
|
Voici un résultat récent qui peut conforter cette vision et s'interpréter comme une modification du génome par la cellule cancéreuse elle-même : brève La Recherche, 404, janvier 2007, 31 (A.-M. Pearse et K. Swift, 2006, Nature, 439, 549) |
|
certaines cellules cancéreuses chez le diable de Tasmanie présentent des anomalies chromosomiques caractéristiques (sur 14 chromosomes: perte des chromosomes 2 , des chromosomes sexuels et d'un seul des chromosomes 6, délétion du bras long des chromosomes 1) que l'on retrouve dans les tumeurs d'animaux différents mais pas bien sûr dans les cellules saines. L'idée que cela puisse être la cellule elle-même qui modifie son caryotype semble tellement peu naturelle aux auteurs qu'ils proposent l'idée d'un cancer contagieux par allogreffe (transmission et greffe d'une cellule étrangère). |
|||
| |
|
|
|
Comment distinguer une anomalie caryotypique d'une différence spécifique, témoin d'une spéciation ? |
|||
|
article
E.U. : |
|
Bernard Dutrillaux, Jérôme Couturier et Jean Chaline ont étudié les caryotypes dans la lignée humaine. Ils proposent un modèle chromosomique et climatique en 3 étapes. Les comparaisons entre caryotypes, et les hypothèses émises pour passer de l'un à l'autre, reposent toutes sur l'idée selon laquelle le caryotype est la marque de l'espèce et que toute spéciation conduit forcément à une empreinte caryotypique nouvelle. Bien évidemment ces études caryotypiques ne se font pas sur de l'ADN fossile (trop dégradé et partiel... voir plus bas) mais sur des espèces actuelles vivantes. |
|
Mais il est clair que si l'on ne dispose que du seul caryotype, sans données morpho-anatomiques, on ne peut spéculer sur une spéciation étant donné que l'on ne possède pas de modèle qui ferait le lien entre la mise en place du caryotype et l'espèce. Dans l'état actuel des connaissances il me semble qu'il n'y a donc pas de limite claire entre variation caryotypique intraspécifique et interspécifique. |
|||
| |
|
1.2 la séquence de l'ADN comme marqueur de l'individu et de l'espèce L'idée d'une séquence de notre ADN (suite des bases composant la chaîne double brin d'une molécule d'ADN théorique que l'on trouverait dans chacune de nos cellules) qui nous caractériserait ou - pire encore - qui nous définirait (au niveau de nos structures et de nos fonctions), si elle a été un rêve pour certains, n'est pas en accord avec les résultats de la génétique. |
|||||
|
|
|
|
|||||
| |
|
|
|
Les empreintes d'ADN sont une signature individuelle |
|||
|
|
|
Analyse génétique moderne, Griffith et al., 2001, DeBoeck Université, ch12 page sur les génomes |
|
La génomique est la caractérisation moléculaire de génomes entiers (c'est-à-dire de l'ensemble de l'ADN nucléaire - et/ou mitochondrial ou plastidien - extrait). Elle utilise des technique particulières de manipulation de très longues séquences d'ADN plus ou moins purifié. La génomique structurale étudie la nature physique des génomes, alors que la génomique fonctionnelle s'intéresse au protéome (gènes associés aux protéines) et aux différents modes d'expression des gènes (voir cours de 1èreS). |
|||
|
Pour le vocabulaire de cette partie voir cours de terminale (spécialité) : les outils des biotechnologies génétiques |
|
|
|||||
|
5
tracés
d'empreintes
ADN (Southern
Blot) |
|
Les empreintes d'ADN sont habituellement réalisées à partir d'une sonde de VNTR (variable number tandem repeats = répétitions en tandem en nombre variable) sur de l'ADN total extrait coupé par une enzyme de restriction. Les fragments obtenus ont de 1 à 5 kb (kilobases = 1000 bases) et sont des zones de répétitions d'un motif unique de 15 à 100 nucléotides (ces zones VNTR sont appelées aussi ADN minisatellite). |
|
On peut aussi réaliser une empreinte d'ADN à partir de l'ADN microsatellite (zones de dinucléotides répétés en nombre variable et dispersées) . D'autres techniques (marqueurs moins universels) peuvent être employées. La technique de coloration et de migration employée est le Southern Blot (pour une présentation et une discussion de cette technique voir cours terminale spécialité). Il n'est pas rare qu'en criminologie on trouve des séquences d'ADN que l'on amplifie par PCR (voir cours terminale spécialité) afin de voir si, dans ces séquences il n'y aurait pas des VNTR afin de pouvoir faire une empreinte d'ADN. |
|
On ignore l'origine et le rôle de ces zones à répétition dans l'ADN humain même si des hypothèses sont émises (voir cours) . Remarque: Par contre il est assez bien établi que ces zones présentent une signature individuelle claire: deux individus n'ont pas les mêmes fragments de restriction, c'est-à-dire les mêmes sites de coupure, c'est-à-dire les mêmes séquences de répétitions de VNTR. On peut conseiller le site de l'université Paris 6 : http://www.edu.upmc.fr/sdv/ masselot_05001/ applications/typage.html |
|
|
|
|
|
|
||||
|
|
Chaque technique permet de fournir un argument technologique (et scientifique) de l'identité d'un reste d'ADN (et donc est présenté avec une incertitude) mais ne constitue jamais une preuve (le domaine de la preuve est un domaine juridique ou légal). |
|
Voir discussion dans le cours de terminale et aussi les illustrations d'un cours de biochimie de la fac d'Orléans, qui semble très bien fait: mais le texte manque; http://www.univ-orleans.fr/sciences/ BIOCHIMIE/L/ Illustrations%20cours/ SC-L5BO-08%20Genetique% 20Appliquee/Les%20 tests%20genetiques%20 -%20Cours%206.pdf |
||||
|
À l'heure des NCIS, Bones et autres Experts, la police scientifique n'a jamais autant occupé le devant de la scène... mais que d'énormités dans ces séries (il y a de bonnes idées de TPE là-dedans). |
|
|
|||||
| |
|
L'empreinte génétique n'est donc pas, comme certains l'ont cru, et le colportent encore, une sorte de code figé dans une molécule, mais ressemblerait davantage à l'empreinte de nos pas qu'on laisserait sur la vase et qui permet de remonter à nos caractéristiques anatomiques... Il me semble que l'on peut affirmer que l'individualité des marqueurs génétiques (essentiellement les zones répétitives d'ADN qui n'ont pas de rôle dans la fonction de synthèse protéique et ne sont donc pas des gènes) est un indice supplémentaire montrant que l'ADN conserve une mémoire de l'histoire (du développement) et du fonctionnement de l'organisme; cet ADN n'est pas à l'origine d'une fonction propre mais bien la trace des dynamiques de l'organisme. |
|||||
| |
|
|
|
De nombreux marqueurs génétiques sont connus et permettent de comparer des organismes de la même espèce, d'espèces voisines ou d'organismes très éloignés |
|||
| |
|
La plupart du temps le paradigme moléculaire fait considérer une séquence d'ADN marqueur comme un élément stable du génome, transmis héréditairement et qui a une fonction connue. |
|
Cette vision est contradictoire puisque dans certains cas les séquences marqueurs ont des rôles inconnus et ne semblent pas être transmises héréditairement mais bien résulter d'un métabolisme. La liste des marqueurs utilisés sort du cadre de ce cours. On peut regretter que les hypothèses émises pour préciser le cadre d'utilisation de chaque marqueur soient moins souvent exposées que les résultats plus ou moins médiatisés obtenus par ces marqueurs comme arguments phylogénétiques. |
|||
|
brève La Recherche, 404, janvier 2007, 31 (C. Murgia et al., 2006, Cell, 126, 477-487) |
|
Voici
un exemple
récent
pour le cas de
cellules
cancéreuses
chez le Chien
: Ce résultat peut cependant s'interpréter comme une modification du génome par la cellule cancéreuse elle-même; les similitudes génétiques reflétant les similitudes des dynamiques cellulaires impliquées. |
|||||
| |
|
|
|
l'ADN ancien et dégradé peut être utilisé par comparaison avec l'ADN des organismes vivants |
|||
|
Freshly
excavated
fossil bones
are best for
amplification
of ancient DNA
(2007).
Mélanie
Pruvost,
Reinhard
Schwarz,
Virginia Bessa
Correia,
Sophie
Champlot,
Séverine
Braguier,
Nicolas Morel,
Yolanda
Fernandez-Jalvo,
Thierry
Grange, and
Eva-Maria
Geigl. Proc.
Natl. Acad.
Sci. USA.
Publié
online le 8
Janvier 2007 |
|
La paléogénétique a fait des progrès très importants : |
|
L'ADN conservé dans des os en cours de fossilisation se dégrade jusqu'à 50 fois plus lors du stockage dans un musée, que lorsque ces os sont enfouis dans le sol. Les ossements archéologiques et paléontologiques doivent être traités comme des échantillons biologiques pendant et après leur prélèvement. On recherche dans les séquences récupérées et amplifiées, que l'on considère comme fossiles, des marqueurs que l'on utilise en typage génétique comme nous l'avons survolé ci-dessus. |
|
Mais, cette extension des méthodes précédentes à des fragments d'ADN plus ou moins dégradés que l'on amplifie (par PCR) repose sur des hypothèses encore plus hasardeuses et souvent au niveau de l'espèce puisque l'on s'efforce d'établir des phylogénies. |
|
| |
|
|
|
Un gène code-barre, marqueur de l'espèce... une utopie ? |
|||
|
http://www.snv.jussieu.fr /vie/dossiers /barcode /barcode.htm http://www.barcoding. si.edu/ DNABarCoding.htm Document assez complet en anglais: http://www.barcoding.si.edu/ PDF/ BLIllustrated26 jan04v1-3.pdf Un article (en anglais) montrant les limites de la méthode: Kipling W. Will, Daniel Rubinoff, Myth of the molecule: DNA barcodes for species cannot replace morphology for identification and classification, Cladistics, 20, 1, 47-55, 2004 |
|
L'idée d'un code-barre moléculaire spécifique (qui permette d'identifier avec certitude une espèce) à partir d'un segment d'ADN pour tous les êtres vivants est simple mais à mon sens utopique.
|
|
Le barcoding ou plus exactement lifebarcoding (http://www. barcodeoflife .org/) a été initié à l'université de Guelph en 2003 (BOLI = BarCoding of Life Initiative). La séquence utilisée est une petite région du gène mitochondrial qui code pour la sous-unité I du complexe de la cytochrome oxydase (COI, voir par exemple sur la base pdb, l'oxydoréductase du cœur de bovin 1occ, ou 1hrc sur la base des protéines membranaires). Mais en cas d'incertitude il est prévu que l'on puisse séquencer d'autres parties du gène, voir le gène entier, puis d'autres gènes et des ARN (voir par exemple : http://www.coreocean.org/Dev2Go.web?id=255158). Indépendamment des polémiques théoriques qu'elle soulève, le barcoding nécessite une méthode standardisée d'extraction de l'ADN, de préparation des échantillons, d'amplification, de séquençage... et a un coût non négligeable.
|
|
Depuis 2003, la méthode, initialement basée sur une seule séquence à été progressivement étendue à d'autres séquences, puis intégrée de façon plus ou mois forcée (du fait de l'hégémonie de la biologie moléculaire dans les sciences de la vie) à une démarche d'inventaire patrimonial en faisant fi des questions de signification des résultats obtenus. Un aveuglement inquiétant: « La valeur des codes barres fait l'objet de nombreux débats dans la communauté des systématiciens ; fait intéressant, ceci ne se reflète pas dans la communauté des utilisateurs potentiels qui y voit surtout les bénéfices qu'elle peut en tirer. La tension est principalement liée à la façon dont les codes barres seront utilisés.» Dr Richard LANE, Directeur scientifique du Natural History Museum, Londres (GB) lors du Symposium Buffon en octobre 2007 au MNHN De plus, aux USA le barcoding est clairement intégré à une vision écologique industrielle et commerciale (voir le blog http:// phe.rockefeller. edu/ barcode/). Des utilisations ponctuelles sont incontestablement intéressantes pour certains groupes d'organismes et pour certaines missions ponctuelles. Il existe en effet une forte demande de détermination des espèces "absolue" (scientifique, répétitive et standardisée, avec une incertitude minimale.... bref, un rêve positiviste, si l'on se réfère à la notion d'espèce, on est au cœur du sujet de la signification de l'identité moléculaire...) de la part des systématiciens travaillant par exemple sur des collections de spécimens, plus ou moins anciens, dans les muséums notamment ou encore dans des recensements sur le terrain. |
|
| |
|
1.3 Les gènes héréditaires, une théorie ancienne sur les caractéristiques héréditaires des individus au sein d'une parenté |
|||||
|
|
Les caractères héréditaires et leur support matériel (les gènes, dans la théorie héréditaire mendélo-morganienne) sont considérés comme caractéristiques de l'individu ou de la fratrie et ils s'échangent au sein de la parenté. |
|
Ce ne sont pas des caractères de l'espèce mais des individus. C'est leur héritabilité qui intéresse le théoricien dans la droite ligne du travail des sélectionneurs du XIXème siècle. |
|
Cette
théorie
est
déjà
un peu
ancienne mais
est toujours
enseignée. Des confusions graves ont été faites par les enseignants à cause des formulations des programmes entre gène héréditaire et moléculaire et il n'est pas rare que des élèves, voire des collègues, confondent ces notions et croient que l'emplacement d'un gène moléculaire sur une carte chromosomique à haute résolution (par FFLP et SSLP principalement) permet d'en déduire un mécanisme de transfert héréditaire, ou, pire encore, que ce gène moléculaire correspond forcément à un gène héréditaire, support d'un caractère phénotypique. |
||
|
2. les empreintes digitales, les iris et autres marqueurs morphologiques |
|
en travaux |
|||||
|
E.U.
empreintes
digitales |
|
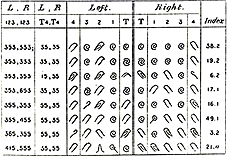
Personne ne remet en cause l'individualité de marqueurs morphologiques, facilement accessibles et qui sont utilisés depuis fort longtemps dans l'identification judiciaire. Chaque empreinte possède un dessein particulier mais ce sont les caractéristiques des desseins situés entre les crêtes qui permettent une identification sûre : |
|
« L'élément
fondamental
n'est
cependant pas
le dessin des
crêtes,
mais le nombre
de triradii ou
deltas :
il s'agit de
disposition en
étoiles,
qui
s'inscrivent
entre les
branches des
dessins. Deux
deltas
correspondent
à un
tourbillon ;
s'il n'y a
qu'un delta,
il s'agit
d'une
boucle ;
l'absence de
deltas produit
un arc, avec
des
crêtes
sinueuses et
parallèles
d'un bord
à
l'autre. Il y
a une
corrélation
stricte entre
nature du
dessin et
nombre de
deltas. |
|
« Les
figures
dessinées
par les
crêtes
dermo-épidermiques
de la face
palmaire de la
main et des
doigts et de
la face
plantaire du
pied et des
orteils, sont
appelées
dermatoglyphes
(du grec derma, -tos,
peau, et gluphê,
gravure). Ils
n'existent
nulle part
ailleurs. Aux
doigts, ils
sont synonymes
d'empreintes
digitales. |
|
| |
|
|
|||||
|
Quelle est l'origine des formes des crêtes ? |
|
Des modèles externes (voir ancienne page sur les modèles pour la différence entre modèle interne et externe) ont été élaborés (voir ci-dessous) mais ce sont les modèles internes, qui s'intéressent à la genèse des formes (littéralement morpho-genèse) qui intéressent davantage le biologiste. |
|
|
|||
|
modèle externe
une
page
très
documentée
qui propose
des
modèles
externes, |
|
modèle interne Si l'on accepte de passer sur quelques paragraphes agaçants, je recommande la prise de vue que fait Vincent Fleury sur la morphogenèse des empreintes digitales dans son ouvrage : Des pieds et des mains (2003, pp 25-31). Car l'apparition des crêtes qui sont des PLIS à la surface de la peau des doigts (qui est présentée judicieusement comme un matériau FIBRÉ - voir cours sur les protéines: exemple de la peau) est un problème de morphogenèse et la physique est très bien outillée pour modéliser ces phénomènes.
Remarque: |
|
«
Il y a donc un
lien entre la
forme des
empreintes et
le
détail
de la forme du
doigt. Mais
quel est ce
lien ? Il est
fort simple
à
comprendre. Il
tient au
caractère
tissé,
fibré
de ces
empreintes
digitales. Des
pieds et des
mains
(Genèse
des formes de
la nature),
Vincent
Fleury,
Flammarion,
col; Champs,
2003), |
|||
| |
|
L'identification humaine par les iris, le réseau veineux des mains ou tout autre caractéristique anatomique mise en place précocement, stable et présentant une forte variabilité individuelle, repose sur les mêmes principes morphogenétiques. Si l'on pouvait observer aisément l'arbre bronchique ou la vascularisation d'un organe, on arriverait à la même variabilité individuelle. C'est une des questions essentielles de la morphogenèse que de comprendre comment peuvent se mettre en place des structures de même forme générale, même ordre de croissance et surtout même fonction, par des chemins différents. Ce que l'on peut exprimer par la finalité ou la contrainte. Actuellement la réponse la plus courante fait une large part à l'émergence et au hasard. Je préfère parler de causalité (ce qui ne signifie pas déterminisme), philosophiquement plus riche. |
|||||
|
3. les groupes sanguins et tissulaires et autres marqueurs immuno-biochimiques |
|
en travaux |
|||||
|
E.U. hématologie |
|
Depuis la découverte en 1901 par Karl Landsteiner des groupes sanguins, maintenant nommés A, B, AB et o, on a trouvé de très nombreuses molécules membranaires de types glucides, lipides ou protides qui assurent une reconnaissance entre cellules ou entre des molécules circulantes (comme les anticorps ou immunoglobulines) et les cellules. |
|
La découverte par Jean Dausset d'antigènes des leucocytes (HLA : human leucocytes antigens) postulés dès 1952, va élargir encore le champ des possibilités d'originalité des combinaisons de marqueurs cellulaires. On va progressivement considérer que ces marqueurs définissent le "soi" immunitaire (s'opposant au "non soi") qui permet à l'organisme de déterminer si une cellule lui appartient ou est étrangère. Mais le système de reconnaissance supposé va rapidement être limité au système immunitaire, de nombreuses cellules ne possédant pas ces marqueurs. De plus les gènes impliqués dans leur synthèse (CMH : complexe majeur d'histocompatibilité) se révèlent très complexes tant au niveau de leur organisation que de leur expression. Actuellement ces marqueurs sont des éléments essentiels du système immunitaire mais on ne les voit plus comme un système général et universel de reconnaissance cellulaire. |
|
La génétique et l'étude de la transmission héréditaire de ces marqueurs cellulaires reste un des domaines de fécond travail en commun entre médecins hématologistes, immunologistes, biologistes moléculaires, généticiens... et, comme dans tout domaine où sont principalement mises en avant les applications pratiques (et ici, les applications médicales), la théorie reste assez stable et assez nettement ancienne. |
|
| |
|
|
|
|
|||