Thème 2 - Des
débuts de la génétique aux enjeux actuels des
biotechnologies (10 semaines) - en cours de
modification 2006
retour accueil, plan
du cours de spécialité, thème
1 (géologie)
Mon souhait de ne pas m'impliquer dans
l'histoire des sciences qui est un domaine trop spécifique
pour qu'un petit professeur de SVT ose s'y aventurer à fait
long feu. Un enseignant est obligé de s'impliquer.
J'espère ne pas colporter trop d'idées fausses. Les
sources sont citées. Cette page met à jour les
idées présentées dans la page
d'histoire
de la génétique
(obsolète) mais les met surtout en forme dans le respect du
programme d'enseignement de terminale S, spécialité
SVT.
Sources:
http://www.mendelweb.org/
http://www.mendelweb.org/MWsapp.html
(texte intégral d'un article
de Jan Sapp: The Nine Lives of Gregor Mendel, in
Experimental Inquiries, edited by H. E. Le Grand, (Kluwer
Academic Publishers, 1990), pp. 137-166)
TOUS les ouvrages de Jan Sapp sont sans aucun doute à
recommander mais je ne les ai pas lus: Beyond the
Gene, New York: Oxford University Press, 1987; Genesis:
The Evolution of Biology. New York: Oxford University
Press, 2003 or Where The Truth Lies. Franz Moewus and the
Origins of Molecular Biology. New York: Cambridge University
Press, 1990.
Un cours de génétique accessible (avec pas mal de
parties hors programme de TS) http://www.edu.upmc.fr/sdv/masselot_05001/introduction/introduction.html
Plan de cette page:
Exercices corrigés
complémentaires
partie 1: Une
théorie de l'hérédité déjà
ancienne
Ce qui importe ce n'est pas d'essayer de
retrouver les motivations réelles de Mendel mais de
démêler ce que les successeurs de Mendel ont fait du
travail de Mendel et pourquoi il fût érigé en
père fondateur de l'hérédité du
XXème siècle.
1ère étape: Mendel, Johannsen et
De Vries: les gènes, particules
héréditaires
TD corrigé
Mendel, Johannsen et De Vries
Les travaux de frère Grégor (Johann Mendel)
publiés en 1866 s'insèrent dans une période de
l'histoire de l'Europe favorable aux sciences qui a vu le
développement de la théorie
cellulaire (énoncée par Schwann et Schleiden en
1838-1839 et précisée par Virchov en 1858; tout
être vivant est composé uniquement et au moins d'une
cellule; toute cellule est issue d'une autre cellule). Dans la
lignée du travail de nombreux sélectionneurs de
races végétales et animales, Mendel réalise un
travail que nos contemporains aiment à présenter comme
un modèle du genre scientifique
(hypothético-déductif ou
hypothético-vérificatif), probablement de façon
très peu historique (http://www.mendelweb.org/MWsapp.html):
un travail basé sur une hypothèse (la
transmission des caractères héréditaires aux
descendants par des particules présentes chez les parents),
prouvé par la réalisation et l'interprétation
d'expériences rigoureuses de croisements chez le Pois
(Pisum). C'est un des précurseurs de l'utilisation des
statistiques dans l'analyses des résultats
expérimentaux.
Les travaux de Mendel seront faussement
(http://www.mendelweb.org/MWsapp.html)
"redécouverts" et réinterprétés par une
première génération de généticiens
comme Bateson) qui a fortement contribué au mythe du
père fondateur.. Ce sont des expérimentateurs,
"discontinuistes" et tout d'abord non darwiniens, qui s'opposaient
alors aux statisticiens "continuistes" et darwiniens. Hugo De Vries
(travaille sur Oenothera lamarckiana et)
nomme mutations les changements brusques de
caractères observés dans la descendance
(page
complémentaire sur les
mutations). L'histoire n'a pas
retenu le terme de pangène donné par De Vries
aux particules héréditaires mais elle a retenu celui de
gène donné par le danois Johannsen (ainsi que
phénotype et génotype).
Quelques dizaines d'années plus tard le darwinisme est
incorporé au mendélisme, notamment par Fisher
(1936).
Au début du XXème siècle on énonce
2 lois EN HOMMAGE à Mendel:
* 1ère loi: loi de pureté des gamètes (un
hybride formé de deux traits d'un même caractère
donnera en quantités égales des cellules sexuelles
contenant soit l'un soit l'autre de traits de ce caractère)
(voir TD).
* 2ème loi: loi de ségrégation
indépendante des caractères (deux
caractères indépendants, et
qualifiés tous les deux de mendéliens, sont
séparés indépendamment dans les gamètes
et réunis indépendamment à la
fécondation; ce qui se voit lors d'expériences de
dihybridisme (voir TD).
Cette loi n'est pas du tout universelle puisque de
nombreux couples de caractères vont avoir une transmission non
indépendante ou liée, ce qui sera la base de la
théorie héréditaire reprise par
Morgan.
2ème étape: Sutton et Boveri,
Morgan et Sturtevant: les gènes, unités mutables
appartenant à un groupe de liaison (le chromosome)
Remarque:
Dans cette nouvelle partie ont peut parler de
caractères et de traits de caractères
mais le terme d'allèle peut aussi être
utilisé car il semble dater de 1902 (c'est à Bateson
que l'on doit ce mot comme ceux d'homozygote,
d'hétérozygote ou de
génétique (1905); dans ce tout début
du XXème la génétique devient une science
à la mode et des chaires universitaires de
génétique sont créées).
Il semble que ce soit Sutton (1903) et Boveri (1904)
qui proposèrent pour la première fois d'associer les
gènes aux chromosomes qui deviendraient ainsi
supports de l'hérédité
(Belin doc B4
p 125) (le comportement des chromosomes lors
de la mitose a déjà été décrit par
Flemming
en 1879 mais c'est vraiment au cours du début de 20ème
siècle que se développent les observations
cytologiques).
N.B. le terme de chromosome
désigne en cytologie une structure colorable qui
contient un centromère, zone d'attachement des fibres
kinétochoriales qui assurent son mouvement. Le
chromosome n'existe donc que sous forme compacte et lors d'une
division.
Thomas Morgan travaille sur les mutants de la mouche du vinaigre :
Drosophila melanogaster. Après s'être
opposé à la théorie chromosomique de
l'hérédité, il en deviendra le fervent
défenseur. Morgan obtient un mutant mâle aux yeux blancs
(Belin doc A3 p 126) dans une
population aux yeux rouges (caractère sauvage). Comme ce trait
de caractère n'apparaît que chez le mâle,
l'idée lui vient de l'associer à un chromosome sexuel
car il existe, chez la drosophile comme chez l'homme, un
déterminisme chromosomique du sexe: les femelles
possédant une paire (XX) d'homologues
(confirmé par N. Stevens, élève
de Morgan, en 1909) et les mâles un chromosome X et un
chromosome Y (Belin doc A2 p
126).
|
Annexe: autosomes et gonosomes
|
|
Les caractères associés aux chromosomes
peuvent en première approximation être
associés à n'importe quelle partie de chaque
chromosome (à l'exception notable du
centromère, de séquence très
particulière, qui est la zone d'attachement des
fibres kinétochoriales).
Chaque paire de chromosomes homologues (autosomes)
associe les MÊMES CARACTÈRES (donc les
mêmes gènes) qui, bien sûr, peuvent
différer selon leurs allèles
(différents s'ils sont
hétérozygotes ou identiques chez les
homozygotes).
Les chromosomes homologues sont qualifiés
d'autosomes. Les chromosomes sexuels (gonosomes) ne
sont homologues (chez la Drosophile et chez
l'homme par exemple... mais il y a beaucoup de
déterminismes génétiques
différents dans le monde vivant) que pour la
femelle (XX). Chez le mâle l'X et l'Y ne sont pas
homologues, du moins sur toute leur longueur. Il existe en
effet ce que l'on appelle une partie propre de l'X et une
partie propre de l'Y.
- Si un caractère est associé à la
partie commune de l'X et de l'Y, sa transmission sera de
mode autosomal (pas de différence avec un
caractère associé à un
autosome).
- Si un caractère est associé à la
partie propre de l'Y, il sera présent que chez les
mâles et aura une transmission de type toujours
dominant (quelque soit l'allèle présent il
est toujours seul et peut donc être
exprimé).
- Si un caractère est associé à la
partie propre de l'X, on dit que le caractère est
"lié au sexe". Il n'est présent
qu'en un seul exemplaire chez le mâle (qui ne porte
qu'un X) car il n'est pas porté par l'Y.
|
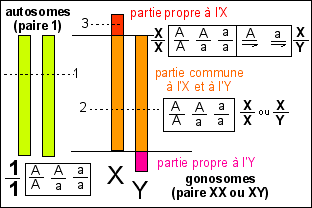
Une représentation très théorique des
gonosomes humains ou de la drosophile comparés
à une paire d'autosomes (n°1). Un
caractère présent sous deux allèles (A
ou a) pourrait présenter les génotypes
suivants selon qu'il est associé aux autosomes (1) ou
aux gonosomes en position 2 (autosomale) ou 3 (lié au
sexe).
|
|
Remarque:
Pour les curieux, je vous invite à consulter une
représentation
plus exacte des gomosomes humains
afin que vous compreniez un peu ce que le discours qui
précède a de simplificateur.
|
1er exercice.
Avec ce que vous savez du support chromosomique de
l'hérédité comment Morgan a-t-il pu obtenir
des femelles à yeux blancs
?
(Belin doc B4 et B5 p
127).
Si le caractère couleur des
yeux dont "yeux blancs" est l'allèle muté est
lié au sexe, donc associé à la partie propre
du chromosome X (voir ci-dessus), une femelle aux yeux blancs
possède donc deux X associés à
l'allèle "yeux blancs". Un X lui vient de son parent
mâle et un X de son parent femelle. Son parent mâle
était donc à yeux blancs et son parent femelle
hybride à yeux rouges. Pour obtenir une femelle hybride
à yeux rouges il faut croiser un mâle à yeux
blancs avec une femelle sauvage (voir F3 dans le cadre
ci-dessous).
Faites une analyse formelle (avec un
tableau présentant les types de gamètes et les
phénotypes des descendants... avec leurs pourcentages
respectifs attendus). Vous noterez "+" l'allèle sauvage
"yeux rouges" et "w" l'allèle muté "yeux blancs".
Les allèles portés par deux chromosomes homologues
sont placés de part et d'autre d'une barre de fraction
(simple ou double). "y" ou un "crochon" ( )
peut désigner le chromosome Y
(pour les tableaux
typographiés on utilisera une double barre oblique et le y
pour le chromosome Y).
)
peut désigner le chromosome Y
(pour les tableaux
typographiés on utilisera une double barre oblique et le y
pour le chromosome Y).
2ème exercice
Faire une analyse formelle du croisement "femelle yeux blancs par
mâle yeux rouges"
(Belin Doc B5 p
127).
|
femelle +//+ x w//y mâle
|
femelle
w//w x +//y
mâle
|
|
F1
|
toutes les femelles ont les yeux rouges (+//w) ainsi que
tous les mâles (+//y)
|
toutes les femelles sont hybrides et ont les yeux rouges
(+//w) et tous les mâles ont les yeux
blancs
|
|
F2
|
femelles +//w x +//y mâles
|
|
+
|
y
|
|
+
|
+//+
|
+//y
|
|
w
|
w//+
|
w//y
|
femelles à yeux rouges (pour moitié +//+
et pour moitié w//+)
mâles pour moitié à yeux
rouges (+//y) et pour moitié à yeux
blancs (w//y)
|
femelles +//w x w//y mâles
|
|
w
|
y
|
|
+
|
+//w
|
+//y
|
|
w
|
w//w
|
w//y
|
femelles pour moitié hybrides à yeux
rouges (+//w) et pour moitié à yeux blancs
(w//w); mâles
pour moitié à yeux rouges (+//y) et pour
moitié à yeux blancs
(w//y)
|
|
F3
|
|
|
+
|
y
|
|
+
|
+//+
|
+//y
|
|
w
|
w//+
|
w//y
|
|
|
|
w
|
y
|
|
+
|
+//w
|
+//y
|
|
w
|
w//w
|
w//y
|
|
,
|
|
les femelles à yeux blancs
(w//w) apparaissent
chez 1/16 des femelles et les mâles aux yeux blancs
(w//y) chez 1/8 des mâles, en supposant que
chaque croisement se réalise avec la même
probabilité et que la proportion de mâles et
de femelles est la même pour tous les croisements
(cela fait beaucoup d'hypothèses et l'on
comprendra que ces valeurs sont tout à fait
théoriques... il ne s'agit d'ailleurs pas ici de
probabilités au sens strict mais de proportions
théoriques).
|
Remarques:
* Ces expériences sont historiquement le point de
départ de nombreuses études qui conforteront la
théorie chromosomique de
l'hérédité.
* Une mutation est une anomalie génétique
conservée (non réparée) lors de la
reproduction. Il faut qu'elle touche donc les cellules
spécialisées (lignée germinale) ou les
premières étapes du développement. Ensuite il
faut qu'elle s'exprime (il vaudrait mieux dire qu'elle
modifie l'expression phénotypique habituelle de la cellule)
c'est-à-dire que par exemple, dans le cas des yeux blancs,
elle provoque un arrêt de la synthèse du pigment
rouge normalement synthétisé par les yeux sauvages.
On notera que si l'allèle "yeux blancs" peut
correspondre à une mutation très ponctuelle au
niveau d'un gène il ne faut pas penser que l'allèle
"yeux rouges" est dû à lui seul au
fonctionnement du gène non muté. Les termes
d'allèles n'ont pas la même signification pour un
allèle sauvage et un allèle muté. C'est
pour cela que la notation "w+" par exemple pour
désigner l'allèle sauvage par opposition à
l'allèle muté (yeux blancs = white) est à
déconseiller.
Morgan découvre avec ses collaborateurs d'autres mutations
dont ils étudient la transmission héréditaire.
Elles forment 4 groupes de liaison: c'est-à-dire
qu'elles ne se répartissent pas au hasard dans les descendants
mais qu'elles présentent une liaison (linkage, en
anglais): elles ont tendance à rester associées. Ils
émettent alors l'hypothèse que ces quatre groupes de
liaison sont assimilables aux 4 paires de chromosomes de la
drosophile. La liaison étant donc "simplement le
résultat mécanique de la localisation des gènes
dans les chromosomes" (groupes présentés en 1915 -
Belin B3 p 129).
Cette théorie s'est trouvée confirmée pour de
très nombreux caractères chez d'autres organismes
eucaryotes chez qui l'on a étudié la transmission de
caractères héréditaires (Pois, Muflier,
Maïs...).
3ème exercice:
Donnez les fréquences attendues pour un croisement entre un
mâle sauvage (corps gris et yeux rouges) et une femelle
double mutante au corps noir et aux yeux marron-sombre (F1) puis
un test-cross (F1 par le parent double récessif) si ces
deux couples d'allèles étaient indépendants.
Même question avec le sexe opposé. On notera les
allèles dominants "+" et les allèles
récessifs "eb" (corps noir = ebony) et
"cn" (oeil marron sombre = cinnabar).
Comparez avec quelques résultats obtenus par Morgan
(Belin B2 p
129). Que pouvez-vous en
déduire ? Quelles hypothèse pouvez-vous
émettre sur le degré de liaison entre les
caractères appartenant à un même groupe de
liaison ?
Il est à noter que les
caractères choisis sont associés à des
autosomes et ne sont donc pas liés au sexe.
|
Hypothèse 1
caractères "couleur du corps" et "couleur
des yeux" indépendants
(étant donné que les
deux caractères sont indépendants, on
notera leurs allèles de part et d'autre d'une
double barre oblique pour chaque caractère
successivement)
|
|
mâle (+//+ +//+) x (eb//eb cn//cn cn)
femelle
|
|
F1
|
toutes les descendants ont le corps gris (ils sont
hybrides +//eb) et les yeux
rouges (ils sont hybrides
+//cn)
|
|
Pour ce test-cross le sexe de l'un ou l'autre des
parents importe peu.
|
|
test-cross
|
(+//eb +//cn) x (eb//eb cn//cn)
|
|
+ +
|
+ cn
|
eb +
|
eb cn
|
|
eb cn
|
+//eb
+//cn
|
+//eb
cn//cn
|
eb//eb
+//cn
|
eb//eb
cn//cn
|
chaque type est représenté dans la
population avec une fréquence de 25% soit 1/4 des
descendants
|
|
Hypothèse 2
caractères "couleur du corps" et "couleur
des yeux" liés
(étant donné que les
deux caractères sont liés, on notera leurs
allèles de part et d'autre d'une seule double
barre oblique)
|
|
mâle (+ + // + +) x (eb cn // eb cn)
femelle
|
|
F1
|
toutes les descendants ont le corps gris et les yeux
rouges (ils sont hybrides pour chaque
caractère)
|
|
Pour ce test-cross le sexe des parents est
important car il n'y a pas de recombinaison chez le
mâle de drosophile, c'est-à-dire que les
caractères sont toujours liés de
façon statistiquement absolue
|
|
test-cross
|
femelle (+ + // eb cn) x (eb cn // eb cn)
mâle
|
,
|
parentaux (1-x)
|
recombinés
(x)
|
|
(1-x)/2
|
(1-x)/2
|
x/2
|
x/2
|
|
|
+ +
|
eb cn
|
+ cn
|
eb +
|
|
eb cn
|
+ + // eb cn
|
eb cn // eb cn
|
+ cn // eb cn
|
eb + // eb cn
|
les caractères restent liés chez
les descendants mais partiellement: les types
parentaux, c'est-à-dire qui
possèdent les couples d'allèles des
parents, sont les plus fréquents (chacun avec une
fréquence de 1/2-x/2); les couples
d'allèles dits recombinés,
car ils recombinent les allèles parentaux, sont
les moins fréquents (avec chacun une
fréquence de x/2). La fréquence de
recombinaison (x) ne doit jamais atteindre 0,5 sinon 1-x
= x et donc les caractères se "comportent"
statistiquement comme des caractères
indépendants. Ils ne sont plus liés
statistiquement, ce qui était notre
hypothèse de départ.
NB: Avez-vous bien compris
que la liaison est ici
statistique ?
La liaison est une association entre un caractère
et un chromosome
(dont on ne
connaît pas encore, à cette étape
historique, le lien physique, qui sera
l'ADN); la liaison la
plus étroite (absolue) donnerait un pourcentage de
recombinaison nul (comme chez le mâle de
drosophile); la liaison la plus lâche donnerait un
pourcentage de recombinaison (type recombiné)
égal au pourcentage de non recombinaison (type
parental), ce qui signifierait que les caractères
étudiés sont statistiquement
indépendants. Le chromosome est bien ici une
notion statistique: c'est une groupe de
liaison dont les caractères sont
liés par des taux supérieurs aux taux de
recombinaison entre caractères. On associe ensuite
ce groupe de liaison statistique à la structure
cytologique "chromosome", ce qui constitue une
théorie chromosomique de
l'hérédité.
|
mâle (+ + // eb cn) x (eb cn // eb cn)
femelle
|
,
|
parentaux
|
|
1/2
|
1/2
|
|
|
+ +
|
eb cn
|
|
eb cn
|
+ + // eb cn
|
eb cn // eb cn
|
Comme il n'y a pas de recombinés chez le
mâle de drosophile, les caractères restent
liés chez les descendants et seuls les types
parentaux, c'est-à-dire qui possèdent les
couples d'allèles des parents sont
représentés (chacun avec une
fréquence de 1/2)
|
|
Analyse des résultats de
Morgan
|
femelle (+ + // eb cn) x (eb cn // eb cn)
mâle
|
,
|
parentaux (1-x)
|
recombinés
(x)
|
|
(1-x)/2
|
(1-x)/2
|
x/2
|
x/2
|
|
|
+ +
|
eb cn
|
+ cn
|
eb +
|
|
eb cn
|
+ + // eb cn
|
eb cn // eb cn
|
+ cn // eb cn
|
eb + // eb cn
|
|
Morgan
|
46%
|
43,7%
|
5,2%
|
5,1%
|
Les caractères sont clairement liés
et le degré de liaison est évalué
par le pourcentage de recombinaison (x = 5,1% + 5,2%
=10,3%)
|
La liaison partielle obtenue pour certains gènes (ceux
étudiés ci-dessus par exemple) à conduit Morgan
à proposer l'hypothèse de la disposition
linéaire des gènes sur le chromosome.
Certains caractères, appartenant au même groupe de
liaison se séparent tout de même lors de la formation
des gamètes. Une première hypothèse cytologique,
toujours en vigueur actuellement, a été de proposer un
mécanisme d'enjambement des chromosomes ou
crossing-over à la prophase de la méiose
(Belin p 130). Le
mécanisme du crossing-over est cependant encore loin
d'être élucidé.
Quelques remarques:
1 - Un taux de 50% de recombinaison ne peut en principe pas
être atteint car cela correspond à deux gènes
indépendants statistiquement. Il ne faut pas oublier que la
notion de caractères liés est une notion
statistique. Vu de cette manière, le chromosome est un
groupe de liaison.
2 - Les crossing-over ne peuvent pas être vus, si ils
existent, lorsqu'ils affectent deux chromatides d'une même
chromosome ou lorsqu'ils affectent des gènes homozygotes
(c'est-à-dire présentant les mêmes
allèles sur la paire d'homologues d'un individu).
3 - Pourquoi le mâle de drosophile présente-t-il une
liaison absolue de ses gènes (pas de crossing-over) alors
qu'il semble que la liaison partielle soit la règle et est
la plupart du temps identique chez les deux sexes ?
4ème exercice:
Chiffrez le pourcentage de recombinaison entre les
caractères partiellement liés vus dans l'exercice
précédent. Les caractères indépendants
se séparent dans les gamètes, les liés
restent ensemble mais se séparent partiellement, on dit
qu'ils se recombinent (sous entendu avec d'autres
caractères).
La dernière hypothèse de Morgan et des ses
collaborateurs est que les crossing-over ont autant de chance de
se produire en tout point du chromosome. Ainsi le taux de
recombinaison (c'est-à-dire la fréquence des
crossing-over dans leur modèle) entre deux gènes est
indépendant de la nature de chaque gène, il ne
dépend que de leur position relative sur le chromosome, ce qui
revient à dire que le crossing-over est un
phénomène mécanique. Ainsi plus les loci (locus
= emplacement) de deux gènes sont éloignés, plus
leur distance génétique est élevée, plus
le taux de recombinaison sera grand. Morgan et ses collaborateurs
établissent ainsi des cartes factorielles qui sont la
schématisation des positions respectives des gènes
évaluées à partir des pourcentages de
recombinaisons. Morgan présente avec Alfred Sturtevant en 1913
la première carte génétique. Depuis on a
défini le centiMorgan (cM) comme unité de
recombinaison: 1 cM étant égal à 1% de
crossing-over.
5ème exercice:
(Belin p
131)
TP5 Nathan p 66-67 Activité 1 et 2; Belin n°7 p
141
|
TP5 Nathan p 66-67 Activité 2 -
Des croisements entre
variétés de tomates
|
|
gène
|
caractères
|
allèle sauvage
|
allèle muté,
récessif
|
|
1
|
uniformité de la couleur de la
feuille
|
vert uni
|
+
|
tachetée
|
t
|
|
2
|
taille du plant
|
standard
|
+
|
naine
|
n
|
|
3
|
peau du fruit
|
lisse
|
+
|
veloutée
|
v
|
|
Pour les trois gènes, l'allèle sauvage
est dominant sur l'allèle muté, parce que
en F1, tous les individus sont de phénotype
sauvage [+ + +].
Le phénotypes sont notés entre crochets
[ffffff].
|
|
On notera que les trois gènes associés
à des trois caractères sont liés
(appartiennent au même groupe de liaison ou
chromosome) parce que les individus issus du test-cross
d'un individu de la F1 avec un double récessif
pour les trois gènes considéré,
présentent des phénotypes (8
différents) qui ne sont pas en proportions
égales. C'est donc bien que ces trois gènes
sont plus ou moins liés entre eux.
On notera donc les trois allèles portés
par chaque chromosome homologue de part et d'autre d'une
simple (ou double ici) barre de fraction (ici
oblique).
|
|
+ + + // + + + x t n
v // t n v
F1
+ + + // t n v
test-cross
(parent hybride issu de F1 x
double récessif pour chacun des trois
gènes)
+ + + // t n v x t n
v // t n v
|
|
+ + +
|
t n v
|
+ n
v
|
t +
+
|
+ + v
|
t n +
|
+ n
+
|
t +
v
|
|
t n v
|
[ + + +]
|
[ t n v ]
|
[ + n v ]
|
[ t + + ]
|
[ + + v ]
|
[t n + ]
|
[+ n + ]
|
[ t + v ]
|
|
nombre de plants
sur 1000
|
417
|
425
|
55
|
59
|
16
|
20
|
3
|
5
|
|
|
La distance entre les gènes 1 et 2 est
mesurée dans notre modèle morganien par le
pourcentage de recombinaison. Les gamètes du
parent hybride (issu de la F1) associent les
allèles + avec + d'une part et t avec n d'autre
part. Les recombinaisons sont donc
réalisées par des associations
+ avec n et
t avec + (notées en bleu
dans le tableau). Il suffit donc d'additionner le
nombre de plants présentant ces recombinaisons et
de le diviser par le nombre total de plants pour obtenir
une évaluation de la distance entre les deux
gènes.
d1-2 =( 55 + 59 + 3 + 5 )/ 1000 =
12,2%
|
|
De la même manière, la distance entre les
gènes 2 et 3 est mesurée dans notre
modèle morganien par le pourcentage de
recombinaison. Les gamètes du parent hybride (issu
de la F1) associent les allèles + avec + d'une
part et n avec v d'autre part. Les recombinaisons sont
donc réalisées par des associations + avec
v et n avec +. Il suffit donc d'additionner le nombre de
plants présentant ces recombinaisons et de le
diviser par le nombre total de plants pour obtenir une
évaluation de la distance entre les deux
gènes.
d2-3 =(16 + 20 + 3 + 5 )/ 1000 =
4,4%
|
|
Pour la distance entre les gènes 1 et 3, on
procède de la même manière mais il
faut faire attention à ne pas oublier des
recombinaisons cachées. En effet, si les
associations + avec v et t avec + sont bien des
recombinaisons, il ne faut pas oublier que l'association
+ avec + et t avec v peut être parentale mais aussi
issue d'un double
crossing-over, une fois entre les gènes 1
et 2 et l'autre fois entre les gènes 2 et 3.
Normalement ces cas de faux
parentaux ne sont pas visibles, sauf, comme dans
le cas ici présent, lorsque les gènes
considérés sont séparés par
un autre gène pour lequel on a déjà
des recombinaisons. Ainsi les deux derniers
phénotypes associent bien les allèles
parentaux pour les gènes 1 et 3 mais pas pour les
gènes 1 avec 2 ni 2 avec 3; il faut donc
les compter doublement pour le
calcul de la distance entre les gènes 1 et
3. Je vous rappelle que la distance
génétique estimée ici repose sur le
pourcentage de recombinaison entre les deux gènes.
S'il y a eu deux
crossing-over entre deux gènes, cela
signifie qu'ils sont éloignés d'autant et
qu'il faut prendre en compte dans le calcul de la
distance ces deux crossing-over. On obtient alors:
d1-3 =(55 + 59
+(2x3) +
(2x5) + 16 + 20 )/
1000 = 16,6%
Ce qui permet de dessiner la "carte
génétique" de ce chromosome de tomate
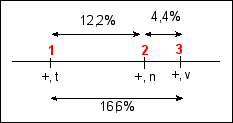
Les distances exprimées en % de recombinaison
pourraient aussi être exprimées en cM ou
unités de recombinaison.
|
|
TP5 Nathan p 66-67 Activité 1 ou Belin exercice
n°7 p 141 - Localisation de trois
gènes sur un chromosome
Cet exercice est identique au
précédent mais il comporte deux
difficultés
supplémentaires...
|
|
gène
|
caractères
|
allèle sauvage
|
allèle muté,
récessif (notations
officielles entre parenthèses, par
simplification j'utilise celle de votre
livre)
|
|
1
|
couleur de l'œil
|
rouge
|
+
|
vermillon
|
v
|
|
2
|
structure de l'aile
|
avec veine transversale
|
+
|
sans veine transversale
|
sv (cv)
|
|
3
|
forme du bout des ailes
|
lisse
|
+
|
incisé (cut)
|
i (ct)
|
|
Pour les trois gènes, l'allèle sauvage
est dominant sur l'allèle muté, parce que
en F1, tous les individus sont de phénotype
sauvage [+ + +].
Le phénotypes sont notés entre crochets
[ffffff].
|
|
On notera que les trois gènes associés
à des trois caractères sont liés
(appartiennent au même groupe de liaison ou
chromosome) parce que les individus issus du test-cross
d'un individu de la F1 avec un double récessif
pour les trois gènes considéré,
présentent des phénotypes (8
différents) qui ne sont pas en proportions
égales. C'est donc bien que ces trois gènes
sont plus ou moins liés entre eux.
On notera donc les trois allèles portés
par chaque chromosome homologue de part et d'autre d'une
simple (ou double ici) barre de fraction (ici
oblique).
|
|
1ère
difficulté: notez bien que, si les parents sont
bien de souche pure, ils ne sont pas sauvages ni
mutés en même temps pour chaque
gène.
+ sv i // + sv i x v + +
// v + +
F1
+ sv i // v + +
test-cross
(parent hybride issu de F1 x
double récessif pour chacun des trois
gènes)
+ sv i // v + + x v
sv i // v sv i
|
|
+ sv i
|
v + +
|
+ + +
|
v sv i
|
+ sv +
|
v + i
|
+ + i
|
v sv +
|
|
v sv i
|
[ + sv i ]
|
[ v + + ]
|
[+ + + ]
|
[ v sv i ]
|
[ + sv + ]
|
[ v + i ]
|
[+ + i ]
|
[v sv + ]
|
|
nombre d'individus
sur 1448
|
592
|
580
|
94
|
89
|
5
|
3
|
40
|
45
|
|
|
Sans refaire toute l'analyse précédente
on peut immédiatement noter que pour les 4
derniers phénotypes il se passe quelques chose
d'anormal par rapport à l'exercice
précédent. En effet le nombre d'individus
recombinés avec un seul crossing over entre les
gènes 2 et 3 (5+3 = 8) est bien inférieur
à celui qui recombinent à la fois les
gènes 1-2 et 2-3 soit (40+45 = 85). Ceci n'est
possible qu'en supposant que les gènes ne sont pas
dans le bon ordre et que les distances 1-3 et 2-3 sont
inférieures à 1-2.
C'est la seconde difficulté.
C'est donc qu'il n'y a qu'un crossing-over pour
les phénotypes [+ + i ] et [v sv +
] mais deux pour les phénotypes [ + sv +
] et [ v + i ].
|
|
En utilisant le même raisonnement que dans
l'exercice précédent, on a donc:
d1-2 =( 94 + 89 + (2x5) + (2x3) + 40 +
45 )/ 1448 = 19,6%
d2-3 =(40 + 45 + 5 + 3 )/ 1448 = 6,4%
d1-3 =( 94 + 89 + 5 +3 )/ 1448 =
13,2%
|
|
Ce qui permet de dessiner la "carte
génétique" de ce chromosome de drosophile
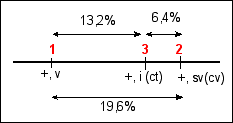
Les distances exprimées en % de recombinaison
pourraient aussi être exprimées en cM ou
unités de recombinaison.
|
Remarques:
* Les exercices scolaires pour le baccalauréat sont
TRAFIQUÉS afin d'avoir des distances qui se somment
algébriquement (ce qui n'est habituellement pas le cas, ce
que l'on explique par le présence des crossing-over doubles
indécelables, laissant invariant la liaison chromosomique
entre les deux loci étudiés: les distances
génétiques sont toujours sous-évaluées
par rapport aux distances cytologiques).
* À lire: la drosophile,
l'outil des généticiens...
Certaines mutations ont aussi un support chromosomique qui peut
être mis en évidence par les techniques de colorations
de bandes des chromosomes. On établit ainsi une carte
cytologique des gènes associés aux mutations.
Il ne faut pas confondre (Belin p
133):
* la carte factorielle (appelée aussi
génétique ou encore chromosomique), établie par
des méthodes statistiques, et
* la carte cytologique établie par hybridation de
sondes radioactives ou par observation de modification de bandes
colorées après mutation.
Globalement, la correspondance est bonne si l'on s'en tient à
la disposition linéaire des gènes. Par contre les
distances sont nettement différentes. Mais la carte
cytologique n'est pas beaucoup plus précise que la carte
factorielle. On ne connaît pas le degré de compaction de
l'ADN dans les chromosomes polytènes... En tout cas on ne
devrait employer le terme de carte chromosomique que lorsque
l'on tient compte à la fois des données statistiques et
cytologiques.
On pourra consulter avec profit les pages de
l'UMPC (Université de Paris 6): http://www.edu.upmc.fr/sdv/masselot_05001/cartographie/cartographie.html.
En conclusion
Les gènes sont des unités de
mutation et de recombinaison: un gène est une unité
mutable appartenant à un groupe de liaison (statistique) : le
chromosome.
La plupart des caractères matériels peuvent
être associés à un ou plusieurs gènes
chromosomiques. Les gènes mendéliens (pangènes)
et gènes morganiens sont des notions qui se recoupent mais ne
se superposent pas. Pour désigner la génétique
chromosomique utilisée dans ce chapitre, on peut parler d'un
modèle héréditaire chromosomique.
Exercice
d'application de type bac: énoncé,
corrigé, présentant
notamment les résultats de Calvin Bridges
(Bridges, C. B. 1914. Direct proof through
non-disjunction that the sex-linked genes of Drosophila are borne on
the X-chromosome, Science, N. S. Vol. XL: 107-109) qui
apportent une confirmation expérimentale de l'association des
gènes, particules héréditaires, avec le
chromosome, structure colorable de la cellule
(l'article est disponible traduit à l'adresse
: http://www.ac-amiens.fr/pedagogie/svt/info/hist_genet/Bridges.html).
3ème étape: Garrod, Beadle et
Tatum, Griffith, Avery.... le gène unité fonctionnelle
et la théorie de l'information génétique
Avant tout, il est indispensable de
comprendre que les apports de la biologie moléculaire du
gène se sont fait DANS LE CADRE DE LA THÉORIE
CHROMOSOMIQUE DE L'HÉRÉDITÉ qui n'est donc
pas, à ce jour (en 2006), remise en cause, du moins par la
majorité des généticiens.
Je m'inscris en faux contre l'affirmation du
programme selon laquelle la biologie moléculaire constitue une
nouvelle rupture. La preuve de cette absence de rupture est bien que
l'on continue toujours d'utiliser le formalisme morganien pour les
allèles. La notion de gène, unité fonctionnelle
se surimpose à la notion de particule
héréditaire mais n'est pas du tout en rupture avec
elle.
Les premières étapes qui on conduit à la
notion de gène, unité
fonctionnelle puis de la découverte de l'ADN et de
la liaison entre l'ADN, l'ARN et les protéines,
ne peuvent faire l'objet de questions à
l'examen (elles ont été rapidement
étudiées (même si ce n'est pas dans leur aspect
historique) en seconde puis en 1èreS);
je laisse donc chacun libre de se cultiver... Elles
sont résumées dans le Nathan sur une double page (p
74-75) et plus détaillées dans le Belin (pp
144-151). Elles sont aussi plus développées dans
la page sur la
génétique de ce site (devenue
obsolète en 2006, un nouveau cours est en
préparation).
La naissance de ce qui est devenu le
paradigme* de
l'information génétique est plus difficile
à situer historiquement. Voir à ce sujet la page
sur la génétique. Toute l'information
nécessaire pour construire le vivant serait contenue dans ses
gènes qui contiendrait en quelque sorte le programme du
vivant. En transmettant des gènes les parents
transmettent à la fois les caractères de
l'espèce et ceux qui déterminent l'individu.
Ce n'est pas une théorie héréditaire, c'est
une théorie partielle du vivant qui fusionne indûment
la notion d'information et de cause
(voir
ci-dessous). La causalité
étant supposée évidente au regard d'une
matière toute puissante. Elle est indissociable d'un
matérialisme (voir par exemple l'article
"forme" de Jean Petitot dans l'Encyclopédie Universalis ou la
page de ce site sur les modèles).
Ce paradigme est actuellement en cours d'abandon. Le cours de ce site
apporte de nombreux éléments qui vont dans le sens
d'une vue plus ouverte du vivant. L'information
génétique n'est qu'une des composantes du vivant. En
utilisant un vocabulaire classique on peut dire qu'il faut lui
associer une information cytoplasmique et une information
environnementale. De plus l'information ne décrit pas seule le
vivant, il faut lui associer la matière et l'énergie.
Dans ces pages le concept unificateur est celui de travail. Un
travail de nutrition, de relation et de reproduction. Voir pour des
généralités le cours de
seconde et pour un approfondissement sur l'information
génétique, le cours le
1èreS. En voici un résumé sous forme de deux
schémas: La vie est un triple
travail de nutrition, de reproduction et de relation; c'est un
mouvement d'énergie, de matière et d'information
(cours de 2nde).
L'information génétique au
cœur de la vie (cours de 1èreS).
Remarque (le 1/05/2005):
Un article de S. Lolle et R. Pruitt paru dans la revue scientifique
Nature (Nature 434, 505 - 509 (24 March 2005)) met en lumière
le fait considéré comme de plus en plus courant qu'il
existe des réversions de certains gènes
associés à des protéines. Une réversion
apparemment dirigée consiste chez un individu
présentant un gène muté identifié,
à récupérer (par un procédé
inconnu mais les auteurs proposent l'hypothèse d'une matrice
d'ARN transmise par une hérédité non
mendélienne...), au bout de quelques
générations, une ou deux copies du gène
ancestral non muté. La fréquence des réversions
peut atteindre quelques dizaines de % des descendants. Ici, ces
chercheurs ont travaillé sur le modèle
génétique usuel qui est une petite plante:
Arabidopsis thaliana, à partir d'un gène HOTHEAD
(HTH) associé à une protéine ACE (ADHESION OF
CALYX EDGES) de 594 aa intervenant dans la signalisation cellulaire
et située pour une bonne part vers l'extérieur de la
cellule; cette protéine possède une activité
enzymatique (de type mandelonitrile lyase qui semble être une
propriété très générale de
nombreuses protéines extracellulaires... voir La Recherche,
386, mai 2005, pp14-15). Le gène HTH associé est
exprimé dans les fruits, les racines, l'inflorescence, les
fleurs, la feuille et la tige; on, connaît une impressionnante
liste de 22 mutants dont ceux étudiés par cette
équipe. La quasi intégralité (il manque au moins
quelques figures...) de l'article en anglais peut être
consultée à l'adresse: http://www.euchromatin.com/Pruitt01.htm
mais il ne peut s'adresser qu'à des enseignants de SVT ayant
un bon niveau de génétique et je ne pense pas qu'il
apporte quelque lumière supplémentaire. Il peut
être complété par les données du site
http://www.arabidopsis.org/
pour les gènes clonés et leurs produits.
Ajout février 2006
À la suite de mon effort de compréhension de la vision
thomienne de la vie (voir page
sur les modèles de René
Thom), je pense qu'un nouveau paradigme pourrait voir le jour. Le
matériel génétique ne serait que la trace stable
des dynamiques de l'organisme vivant. Le chromosome et secondairement
l'ADN serait en quelque sorte l'œil du cyclone
métabolique, le point stable des catastrophes au sens de
Thom. Ce n'est pas la matière qui importe c'est ce dont elle
est la trace. C'est ainsi que lorsque l'on croît voir dans
l'ADN un code de l'identité de l'individu, de ses
capacités et des ses maladies, cela ne serait pas une
véritable erreur. Mais il faudrait alors considérer
l'ADN comme le résultat des dynamiques de l'organisme.
Une matière qui garderait ainsi la trace de l'état
physiologique et serait aussi un miroir des événements
passés. Non pas comme un élément qui CAUSE mais
comme un éléments qui INDIQUE les
phénomènes dynamiques, présents et
passés, complexes et intriqués.... Le gène
moléculaire ne serait plus un code, ni une matière
manipulée mais le
résultat signifiant
de la dynamique de l'être
vivant.
Un ADN transféré seul aurait alors une autre
signification que celle d'un ordre. Pourquoi pas un signal. Les
organismes sélectionnés après transfert
par vecteur seraient ceux qui se sont adaptés et ont
modifié leurs réseau métabolique de telle
façon que tel ou tel caractère apparaisse, dont la
trace serait précisément l'apparition de cet ADN. Pour
les bactéries l'ADN serait peut-être lié de
façon beaucoup plus étroite au réseau dynamique,
ce qui permettrait peut-être d'agir directement sur le
réseau un introduisant un ADN étranger ou en utilisant
le fait que les cellules bactériennes réagissent en
s'adaptant à la présence de tout ADN étranger
dans le milieu. L'ADN étant vu comme un moyen habituel de
communication entre organismes (ou l'ARN, sans oublier le rôle
des virus). Tout organisme s'adaptant aux organismes voisins en
fonction de leur métabolisme, peut être notamment
à l'aide du signal
ADN. Dans ce cas l'ADN aurait aussi le
sens de signal DÉCLENCHEUR mais uniquement pour de l'ADN
étranger qui pénétrerait dans un organisme. Ce
rôle de signal serait certainement très important in
vitro pour des cellules eucaryotes en culture mais perdrait toute
signification globale chez un pluricellulaire. Mais par contre les
virus et autres particules génétiques pourraient garder
ce rôle de signal qui déclencherait un réponse de
l'organisme. Une sorte de moyen de communication au sein de la
population. On verrait ainsi une grippe générée
par un individu affaibli comme un signal transmis à toute une
population comme signe de souffrance. Si l'on compare avec une
substance nutritive qui serait libérée dans le milieu,
le rôle de l'ADN n'est pas très différent.
Plongé dans un milieu auquel on fournit sans cesse du glucose
l'organisme adapte son métabolisme. Plongé dans un
milieu où circulent des fragments d'ADN, l'organisme adapte
ses dynamiques à ces traces des dynamiques voisines qu'il
considère comme issues de cadavres (de cellules ou
d'organismes).
partie 2: Des
biotechnologies génétiques
surévaluées
Les techniques de ce qu'il convient d'appeler le génie
génétique permettent de manipuler les gènes
et dans une certaine mesure le vivant. Les succès obtenus sur
les Procaryotes et sur quelques organismes-modèles eucaryotes
ont pu faire penser à certains qu'ils détenaient les
clés du vivant. On est loin du compte.
1. Les outils des biotechnologies
génétiques
On désigne par "technologie de l'ADN recombinant"
les techniques de manipulation de l'ADN mises au point à
partir des années 1970 et l'on réserve habituellement
le terme de génie génétique aux
manipulations génomiques appliquées. (On notera que le
terme "génie génétique" est nettement moins
péjoratif que "manipulation génétique" qui
fût le premier terme employé). Le génie
génétique est un ensemble de techniques permettant de
former de nouvelles combinaisons d'ADN susceptibles d'être
insérées dans une cellule vivante et y être
exprimées.
Maintenant on parle souvent de génomique, qui
désigne la science des gènes et des techniques de la
génomique pour désigner la technologie.
Voici un aperçu très simplifié des
principales techniques regroupées sous le terme de technologie
de l'ADN recombinant :
|
Technologie de l'ADN recombinant... quelques
techniques
|
|
couper l'ADN
|
les endonucléases ou endonucléases
de restriction ou enzymes de restriction peuvent in
vitro couper spécifiquement une séquence d'ADN
(double brin)
|
Les enzymes de restriction ou
endonucléases de restriction
:
|
|
on nomme ainsi toute molécule capable de
dégrader de façon spécifique
tout ADN étranger. Elles ont
été isolées chez les
bactéries qui dégradaient rapidement
les ADN des virus (bactériophages) qui leur
injectaient leur ADN : ces bactéries
résistantes aux virus, restreignant la
croissance de ceux-ci, furent isolées et
cultivées. On pu alors extraire de leur
cytoplasme des enzymes dites de restriction (qui
provoquaient une restriction de croissance du
virus). Il semblerait que les cellules
bactériennes se protègent contre leur
propres enzymes de restriction en modifiant
chimiquement leur ADN par méthylation.
Actuellement les biologistes moléculaires
pensent que la méthylation est un moyen pour
la cellule procaryote de reconnaître son
propre ADN. Chez les eucaryotes la
méthylation est interprétée
comme un marqueur de l'activité des
gènes... voir à ce sujet par
exemple les Ch 415 et 27 de Gènes de B.
Lewin).
|
nom
|
organisme
|
* = sites de coupure
|
commentaires
|
|
AluI
|
Arthrobacter
luteus
|
5'-----A
G*C
T-----
--------T
C*G
A-----5'
|
F
|
C - donnent deux "bouts collants"
(extrémités
cohésives) puisqu'elle ne coupe pas
les deux brins de la chaîne d'ADN au
même endroit (c'est
un avantage car en traitant avec la
même nucléase l'ADN à
insérer et le vecteur on
prépare leur recombinaison, mais
c'est aussi un inconvénient car des
fragments peuvent se
réassocier...)
F - coupure "franche" sans "bouts
collants"
5' désigne
l'extrémité dite 5' du brin
d'ADN; ce sont les carbones du
désoxyribose qui sont ainsi
numérotés; à
l'extrémité 5' du brin d'ADN
il y a un groupement phosphate dont une
des liaisons est disponible. Le phosphate
du nucléotide de
l'extrémité 3' est
engagé dans une liaison avec le
nucléotide suivant.
|
|
BamHI
|
Bacillus amyloliquefaciens
H
|
5'-----G*GATC
C-----
---------C
CTAG*G---5'
|
C
|
|
BgIII
|
Bacillus globiggi
|
5'-----A*GATC
T----
---------T
CTAG*A---5'
|
C
|
|
EcoRI
|
Escherichia coli
|
5'-----G*AATT
C-----
---------C
TTAA*G---5'
|
C
|
|
EcoRII
|
5'-----*CCTGG
----
---------
GGACC*---5'
|
C
|
|
EcoRIII
|
5'-----G*CCTG
C-----
---------C
GACC*G---5'
|
C
|
|
HaeIII
|
Hæmophilus
ægyptus
|
5'-----G
G*C
C-----
--------C
C*G
G-----5'
|
F
|
|
HindIII
|
Hæmophilus
influenzæ b
|
5'-----A*AGCT
T-----
---------T
TCGA*A---5'
|
C
|
|
HpaI
|
Hæmophilus
parainfluenzæ
|
5'-----GT
T
*A
AC-----
--------CA
A*T
TG-----5'
|
F
|
|
PstI
|
Providencia
stuartii
|
5'-----C*TGCA
G----
---------G
ACGT*C---5'
|
C
|
|
SalI
|
Streptomyces albus
|
5'-----G*TCGA
C----
---------C
AGCT*G---5'
|
C
|
On remarquera que les séquences reconnues
sont généralement de 6 paires de
bases.
|
Remarque: On connaît des
endonucléases qui coupent les ARNm.
|
|
coller l'ADN
(recombiner)
|
les ligases sont des enzymes faisant intervenir
l'énergie de l'ATP (molécule + ATP
<==> ADP
+ molécule - P) ou d'un autre
nucléoside triphosphate (GTP, UTP...).
Certaines ligases sont capables in vitro de lier deux
molécules d'ADN par liaisons covalentes (une sur
chaque brin). Si leurs extrémités sont
cohésives et complémentaires on peut ainsi
former une molécule d'ADN hybride avec deux ADN
provenant d'organismes différents (ADN
chimère). C'est notamment le cas de l'insertion
d'un ADN étranger, isolé par une enzyme de
restriction ER, dans un plasmide coupé par cette
même enzyme de restriction ER.
Dans le cas où les extrémités ne sont
pas cohésives, on peut les rendre cohésives
à l'aide d'une enzyme (transférase terminale)
capable d'ajouter (en 3') une chaîne
monospécifique de nucléotides (poly A ou poly
T par exemple). Il existe aussi une ADNligase
réalisant des ligatures entre molécules d'ADN
à bouts francs (extraite du phage T4).
|
|
dupliquer (multiplier, cloner)
l'ADN
|
* multiplication in vivo
par l'ADN polymérase
la multiplication d'une séquence d'ADN était
auparavant conditionnée par son insertion dans un
vecteur d'insertion (virus ou plasmide) que
l'on introduisait à son tour dans un vecteur de
multiplication (bactérie ou cellule eucaryote
comme la levure) qui, en se multipliant, reproduisait la
séquence d'ADN insérée (génie
génétique). Cette technique est connue sous le
nom de clonage d'un gène (isolement,
insertion, multiplication).
* multiplication in vitro par amplification
enzymatique par l'ADN polymérase extraite d'une
bactérie thermophile. C'est notamment la technique de
l'ACP (amplification en
chaîne par polymérase: ce terme doit remplacer
en français la PCR (polymérisation en
chaîne de l'ADN): http://www.education.gouv.fr/bo/2007/1/CTNX0609645K.htm)
qui permet de produire de grandes quantités d'un
fragment d'ADN sans le cloner.
|
|
synthétiser de
l'ADN
|
la synthèse se différencie de la
multiplication par le fait que l'on procède à
partir de nucléotides et non d'ADN. De plus cette
synthèse d'ADN artificiel peut être
contrôlée de façon à obtenir de
l'ADN de séquence déterminée. Cette
technique n'est au point que pour des
oligonucléotides (quelques dizaines de
nucléotides).
Les oligonucléotides peuvent servir d'amorce
pour des synthèses dirigées d'ADN. Ainsi, on
peut réaliser une synthèse dirigée
d'ADN in vivo à la suite de l'insertion d'un
oligonucléotide portant par exemple une modification
que l'on souhaite conserver dans un gène. Par exemple
dans le cadre de mutations dirigées
(mutagenèse dirigée) on hybride un
oligonucléotide spécifique (amorce) avec un
ADN monocaténaire comportant le gène que l'on
désire modifier. Par l'ADN polymérase on
synthétise in vitro un ADN bicatéanaire
comportant la mutation souhaitée (sur un seul des
brins).
|
|
dénaturer l'ADN
|
la dénaturation de l'ADN est la
séparation des deux brins (par rupture des liaisons
hydrogène) ; elle s'obtient par simple chauffage
modéré (vers 60°C) et est alors
réversible. A plus haute température il
peut y avoir rupture des liaisons covalentes entre et
à l'intérieur des nucléotides et les
brins sont altérés; la dénaturation est
alors irréversible.
|
|
renaturer l'ADN
|
la renaturation de deux brins d'ADN
séparés par chauffage modéré
(vers 60°C) est spontanée dès retour
à une température plus basse.
la renaturation ou l'hybridation (voir ci-dessous) sont
utilisées dans l'association d'une sonde avec
une molécule d'acide nucléique
|
|
hybrider l'ADN
(apparier des séquences d'ADN
ou d'ADN et d'ARN complémentaires)
|
On peut hybrider (c'est-à-dire associer
artificiellement) des brins d'ADN (qu'il faut donc avoir
séparé auparavant par dénaturation)
venant de molécules différentes mais aussi des
molécules d'ARN avec des brins d'ADN ou encore des
brins d'ARN. C'est encore l'hybridation mais cette fois
c'est son caractère spécifique qui est
employé pour l'hybridation d'une sonde (courte
séquence marquée radioactivement (sonde
chaude) ou non (sonde froide, par exemple sonde
fluorescente, voir FISH dans
l'ancien
cours de terminale)...) avec une séquence
d'ADN ou d'ARN complémentaire au sein d'un grand
nombre de molécules d'acides nucléiques.
Les techniques de Northern et Southern blot
appliquées respectivement aux molécules
d'ARN et d'ADN simple brin permettent de visualiser
rapidement l'hybridation entre une sonde radioactive et des
acides nucléiques simple brin séparés
par électrophorèse sur gel et
transférés par effet buvard sur une feuille de
nitrocellulose. On qualifie de "Western blot" une technique
assez similaire à partir de protéines.
|
|
transcrire l'ADN en ARN
|
la transcription est réalisée in
vivo par de gros complexes enzymatiques contenant une
ARN polymérase (voir cours
de 1èreS); in
vitro elle peut être obtenue avec un rendement
plus faible. L'ensemble des ARN transcrits
dans une cellule est appelé transcriptome par
analogie avec le génome désignant
l'ensemble des gènes.
|
|
"retro-transcrire" l'ARN en ADN =
"transcription inverse"
|
des enzymes de type transcriptase inverse (ou
reverse) ou retrotranscriptase (capables de catalyser
la synthèse d'un ADN monobrin (dit ADNc ou
ADN complémentaire à partir d'un ARN,
voir cours de
1èreS) ont été
découvertes par Tenin, Baltimore chez les virus
(1970) et par Beljanski chez Escherichia coli (1972)
puis chez d'autres cellules procaryotes (Agrobacterium
tumefaciens) et eucaryotes (voir
histoire
de la
génétique).
|
|
insérer l'ADN dans une
cellule
(transformer un oragnisme par
transgenèse)
|
l'ADN peut être inséré
* chez les procaryotes et quelques cellules eucaryotes comme
la levure de bière, par l'intermédiaire d'un
vecteur d'insertion de type phage, virus, plasmide, cosmide
(plasmide artificiel d'E. coli contenant le
gène cos du phage lambda),
* chez les eucaryotes par mico-injection (à l'aide
d'une micropipette et d'un micromanipulateur),
électroporation (exposition brève à un
courant électrique de très haut voltage de
cellules de mammifères ou de plantes en
présence d'ADN à insérer), bombardement
(fusil à gènes avec des microprojectiles
couverts d'ADN, utilisé notamment pour obtenir des
maïs transgéniques), agitation ou par
l'intermédiaire d'un vecteur d'insertion
bactérien (cas d'Agrobacterium
tumefaciens...).
|
|
exprimer un gène
|
l'insertion du vecteur dans la cellule transformée
qui se multiplie n'implique pas que le gène
inséré soit exprimé. On utilise des
vecteurs d'expression qui sont souvent des
dérivés du plasmide pBR322 d'E. coli
(voir exercice)
qui contient les signaux nécessaires à la
transcription et à la traduction.
|
|
séparer-séquencer
|
La méthode diffère totalement selon la
longueur de la molécule d'ADN que l'on veut
séquencer.
- la cartographie de restriction : l'action
d'une (ou de plusieurs) enzyme(s) de restriction sur des
molécules d'ADN données fournit un ensemble
de fragments de tailles variées qui dépend
de la séquence de l'ADN coupé. Les
fragments (dits "fragments de restriction"
d'une taille de quelques milliers à quelques
dizaines de milliers de paires de nucléotides)
peuvent être séparés et leur taille
évaluée. Ce qui permet, par comparaison de
l'action de plusieurs enzymes de restriction,
l'établissement de ce que l'on appelle une
carte de restriction. On peut donc penser que deux
cellules possédant la même information
génétique auront exactement la même
carte de restriction. De la même manière,
deux cellules de fonction identique mais prise chez deux
individus différents, sont supposées avoir
des cartes de restriction différentes mais
voisines.
- l'électrophorèse sur gel : le
gel de polyacrylamide permet de séparer des
molécules d'ADN de moins de 500 nucléotides
qui ne différent que par un seul
nucléotide. On utilise des gels d'agarose pour des
molécules de plus grande taille et enfin la
méthode d'électrophorèse sur gel
d'agarose en champ pulsé pour des molécules
d'ADN de très grande taille (millions de paires de
nucléotides). La coloration des fragments d'ADN
est réalisée soit au bromure
d'éthidium, fluorescent en lumière
ultra-violette lorsqu'il est lié à l'ADN,
ou le 32P radioactif (de type béta) que
l'on doit auparavant incorporer aux groupements phosphate
de la molécule d'ADN ce qui implique sa
réplication...
- la méthode chimique et la méthode
enzymatique de séquencage rapide de l'ADN ;
la méthode chimique combine un marquage par
le 32P radioactif, une destruction
modérée de l'une des 4 bases, et une
séparation par électrophorèse sur
gel. La méthode enzymatique repose sur
l'utilisation de l'ADN à séquencer comme
matrice pour réaliser une copie marquée
(radioactivement ou chimiquement) de la séquence
recherchée, suivie par un ajout
contrôlé de nucléotides un par un et
une séparation sur gel de chaque fragment en cours
d'allongement. C'est cette dernière méthode
qui est surtout utilisée dans les automates (voir
par exemple Biologie moléculaire de la cellule,
pp297-298).
|
|
génie
génétique
|
- isolement "du" gène, en fait d'une
séquence d'ADN (essentiellement
représentée par l'utilisation des enzymes
de restriction... voir ci-dessus); cette étape
peut nécessiter l'utilisation de transcriptase
inverse où de cloner le gène à
partir du génome entier (ce qui est la
méthode habituelle et qui permet d'obtenir une
banque génomique qui contient des vecteurs
recombinants (voir ci-dessous) de tous les gènes
d'un organisme) ou partiel...
- insertion dans un vecteur d'insertion (virus ou
plasmide bactérien) qui est aussi appelé
"chimère" ou plutôt recombiné (ADN
recombinant, vecteur recombinant) nécessitant
l'utilisation des enzymes de type ligases...
- transformation (ou transfection ou encore
transduction...) d'un hôte en vecteur de
multiplication par insertion du vecteur modifié
(phase d'amplification) associée aux techniques de
criblage (isolement) des transformants...
- expression de la séquence
insérée et amplifiée
|
|
Exercices d'application: construire
une carte de restriction
simplifiée
Belin n°8 p 164 (le texte du
sujet est erroné, voir ci-dessous les questions
corrigées); Nathan TP1, 2, p 76 et exercice
n°1 p 85
|
|
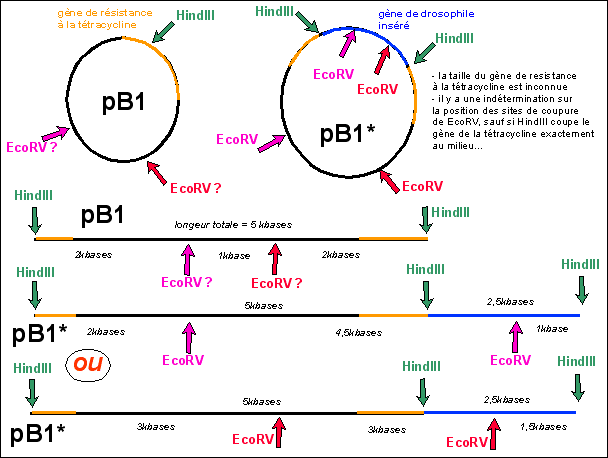
Un plasmide d'une souche bactérienne (B1)
circulaire (que l'on nommera pB1) contient un seul site de
restriction de l'enzyme HindIII. Ce site est situé
dans le gène de résistance à la
tétracycline (un antibiotique) que porte de
plasmide.
On coupe le génome de la drosophile à l'aide
de la même enzyme de restriction HindIII. On obtient
des fragments qui sont insérés (voir
génie génétique) dans des vecteurs
d'insertion qui sont justement les plasmides pB1 de la
même souche bactérienne (B1). Par des
techniques appropriés on crible les transformants
ayant intégré un fragment d'ADN de la
drosophile. On obtient ainsi un ensemble de souches de
bactéries B1* transformées dont leur plasmide
pB1* a intégré tel ou tel gène
drosophilien. C'est ce que l'on appelle une banque d'ADN. En
fait il s'agit d'une banque de clones de souches
bactériennes B1* (recombinées) dont le
plasmide recombiné (pB1*) contient tel ou tel
gène de drosophile.
Le clone n°15 contient par exemple un gène de
drosophile inséré. On analyse l'ADN de ce
plasmide transformé (pB1*) par digestion enzymatique
en présence des enzymes de restriction Hind III et/ou
EcoRV. Les résultats sont présentés
ci-dessous, le témoin étant le plasmide pB1
non recombiné.
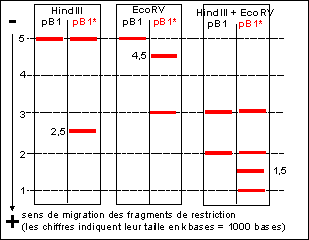
|
Questions:
1) Définir un clone et une banque
génomique.
2) Expliquer l'importance de l'unicité
et de la localisation du site de restriction d'Hind
III
3) Construire la carte de restriction de pB1
et de pB1* du clone 15 pour les deux enzymes de
restriction utilisées ici.
|
|
|
Corrigé
1) Un clone est un ensemble de cellules identiques
génétiquement (voir page sur les
manipulations cellulaires). Ici le terme désigne
plus particulièrement une population
bactérienne issue d'une cellule unique,
génétiquement transformée. Un clone est
donc, dans cet exemple, une souche bactérienne
transformée (B1*) présentant un gène de
drosophile intégré à son plasmide pB1*.
Chaque clone possède un gène identifié
de drosophile. L'ensemble des clones constitue une
banque génomique plasmidique de la drosophile.
La banque n'est complète que si l'on a
l'intégralité du génome de l'organisme
étudié inséré, gène par
gène, dans un très grand nombre de clones. Ce
qui suppose de connaître tous les gènes de
l'organisme... ce qui est pour le moins rare.
D'une façon plus générale, une banque
d'ADN pourrait être définie comme le lieu
où l'on conserve des organismes ou des parties
d'organismes génétiquement identifiés
et transformés.
2) Un plasmide qui présente plusieurs sites de
coupures par une enzyme de restriction (plusieurs sites de
restriction pour une même enzyme) donnera plusieurs
petits fragments d'ADN sans que l'on puisse insérer
un gène en récupérant un plasmide
complet et circulaire. La localisation du site de
restriction au sein d'un gène marqueur (ici un
gène de resistance à un antibiotique) permet
de sélectionner les souches ayant
inséré le gène de drosophile (c'est le
criblage des transformants). En effet, un plasmide
recombiné pB1* (avec un fragment d'ADN de drosophile
qui s'est inséré dans son gène de
résistance à la tétracycline), a un
gène de résistance à la
tétracycline altéré (non fonctionnel).
La souche transformée (B1*) devient donc sensible
à la tétracycline. Il suffit donc, SUR UNE
RÉPLIQUE (bien sûr car sinon on détruit
la souche que l'on recherche), de faire agir la
tétracycline: la souche sensible (détruite ou
dont la croissance est réduite) est la souche
transformée qu'il suffit alors de
récupérer au niveau de la culture ayant permis
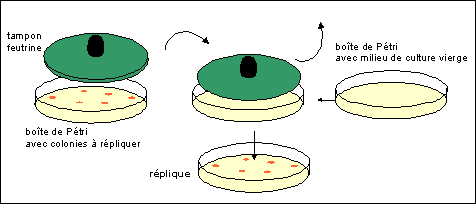
de faire la réplique (voir illustration
ci-dessous).
3)
|
Il me semble bien difficile de
présenter vraiment ces méthodes de façon
accessible à un élève de terminale sans faire
appel à des notions assez complexes sur la structure du
génome et surtout sur la régulation de son expression.
Il me paraît dérisoire de vouloir expliquer une
technique d'hybridation de sonde radioactive alors qu'un
élève de TS ne sait même pas que les gènes
eucaryotes sont morcelés en introns et exons. Certains manuels
scolaires ont d'ailleurs franchi le pas, ce que je ne fais pas
étant donné que la complexité du génome
(et les interactions entre gènes) ne me semble pas changer
fondamentalement le paradigme mendélien-morganien.
Depuis près d'un
demi-siécle de nombreuses voix de scientifiques, de
philosophes...de renom se sont levées pour dénoncer
l'inflation des résultats expérimentaux sans enjeu
théorique. A quoi sert-il d'accumuler des données si
l'on a pas fait l'effort de théoriser la recherche ? La
recherche en biologie moléculaire travaille sur le même
paradigme depuis plus de 50 ans. Il est temps que cela change.
Pour ce qui est des motifs du lobby de la biologie
moléculaire, en France particulièrement, il est clair
que les enjeux sont politiques, et économiques. S'y ajoutent
probablement des enjeux philosophiques et éthiques.
2. Des enjeux économiques, politiques,
philosophiques et éthiques
2.1 de la prudence dans l'utilisation du paradigme de
l'information génétique en justice dans la technique
des empreintes génétiques
Considérations générales:
La technique qui permet de réaliser des "empreintes
génétiques" est une technique comprenant une
dénaturation, une digestion par des endonucléases puis
l'hybridation par une sonde radioactive multilocus (très peu
spécifique) suivie d'une électrophorèse sur gel.
Elle est onéreuse. Elle fournit un cliché qui est
utilisé comme une sorte de carte d'identité
génétique. On peut essayer de mettre en évidence
ce que l'utilisation de cette technique présente comme
hypothèses :
- chaque personne possède une empreinte
génétique (en fait une carte de
restriction-hybridation par la sonde multilocus) qui lui est
propre et qui est la même pour chacune des ses cellules et
qui n'évolue pas dans le temps. Cette affirmation n'est
qu'un postulat, même si il est hautement probable dans la
plupart des cas étudiés ;
- ce qui suppose aussi pour la technique elle-même que
l'action des enzymes de restriction est toujours la même,
qu'elle coupe SANS ERREUR, toujours au même endroit si le
génome est identique. De la même manière
l'hybridation doit se faire de façon TOUJOURS
IDENTIQUE....
On voit aisément que ce qui peut être
présenté avec une INCERTITUDE raisonnable pour une
détermination qui n'est pas aussi grave, n'a pas la même
signification lorsqu'il s'agit de l'utiliser comme PREUVE pour
condamner un prévenu ou faire un recherche de paternité
ou maternité biologique. Comme toute mesure il faut absolument
présenter le résultat avec une incertitude (qui est
souvent absolue car indéterminée statistiquement), aux
jurés d'apprécier ensuite l'utilisation qui doit en
être faite.
Il est certain que l'évaluation de cette incertitude n'est pas
chose aisée mais il faut s'y essayer. La présentation
des incertitudes évaluées le plus honnêtement
possible pour toutes les mesures expérimentales est en quelque
sorte le garant de la crédibilité des scientifiques. Il
n'est pas rare que l'on parle de certitude alors qu'il s'agit de
convictions puisqu'il existe toujours une incertitude
expérimentale.
Exercice du Nathan n°2 p103
Recherche du coupable d'un viol.
Ce problème a fait l'objet d'une partie un peu
détaillée sur une page de ce site sur les
statistiques.
Sur un échantillon de 1.000 tests d'ADN (et pas moins car
sinon l'échantillon n'est pas représentatif) il y a 3
faux positifs (ce chiffre est celui que l'on trouve
dans toute la littérature mais je ne sais pas comment il est
obtenu). Si l'on fait 1.000 tests on doit donc obtenir,
statistiquement 4 profils concordants en supposant qu'il y a un
coupable parmi les testés. Sur les 4 profils identiques au
profil de référence, on a donc un seul coupable, soit
25% de chance de connaître ainsi le coupable. Mais le
problème est que l'on a jamais 1.000 tests mais que l'on
raisonne tout au plus sur quelques échantillons (sauf dans
certains cas très particuliers où la recherche a
été étendue à toute une population...).
Dans ce cas il est clair que l'outil probabiliste est INUTILISABLE.
On peut bien tomber comme on peut mal tomber et surtout on NE PEUT
PAS CHIFFRER L'ERREUR. Un test d'ADN peut indiquer le coupable, comme
il peut être un faux positif, sans aucune certitude. Seul,
c'est un test auquel on ne peut pas accorder de poids. Avec un
faisceau d'autres preuves, il prend du poids, mais pas
statistiquement.
2.2 de l'émergence d'une nouvelle médecine ni
curative, ni préventive mais informative: la médecine
prédictive
La médecine prédictive est un mot nouveau
(prédictif) forgé pour désigner la
médecine de la prévision qui s'ajouterait à
curatif et préventif qui définissent classiquement les
deux aspects de la médecine qui soigne. Cette
"médecine" ne soigne pas, elle informe. Elle est
essentiellement probabiliste et repose sur un paradigme très
fort de déterminisme génétique des maladies.
Elle fourni une indication d'un risque
génétique.
On notera qu'à partir du moment où l'on fait le choix
de tenir compte de ses résultats et d'agir, on sort du domaine
prédictif pour passer dans le domaine préventif,
à partir des éléments obtenus de façon
prédictive.
Le dépistage (étymologiquement "remonter la
piste") est à double sens: il s'agit de repérer, le
plus tôt possible, même sans signe apparent, une maladie
ou une malformation déjà existante mais aussi
repérer des individus présentant un risque
génétique élevé. Si repérer des
individus atteints, même asymptomatiques fait partie de la
médecine préventive, le dépistage du risque
génétique est bien uniquement prédictif.
Le terme de diagnostic enfin repose sur l'identification de
quelque chose de connu (du grec dia = "à travers" et gnosis =
"la connaissance"). On ne peut diagnostiquer qu'une maladie connue.
On ne peut pas diagnostiquer un risque. On peut par contre
diagnostiquer un allèle (ce que l'on nomme "diagnostic
génétique").
La plupart des conclusions en médecine
héréditaire font appel à la théorie
morganienne de l'hérédité à laquelle on a
ajouté la théorie de l'information
génétique. Les techniques modernes de la
génomique ont été mises au service de cette
même théorie sans la remettre en cause. On recherche des
allèles dans des cellules embryonnaires, fœtales ou
adultes et on suppose une causalité directe entre
allèle et phénotype.
Pour ce qui est de l'analyse des arbres généalogiques
(donc humains) on se contente d'utiliser le modèle morganien
de l'hérédité établi à partir de
drosophiles et que l'on extrapole à l'homme puisque des
analyses statistiques de descendance (comme celles
réalisées chez la drosophile ou le pois) y sont
impossibles.
|
Remarques sur les maladies
génétiques
(d'après
Génétique humaine, Jean-Louis Serre et
Josué Feingold, Dossier INSERM Nathan,
1993)
|
|
Une maladie génétique est une
maladie dans laquelle le facteur héréditaire
est principal. Au sujet de la cause des
maladies, voir par exemple une page
sur la santé dans l'ancien
cours de TS. Étant donné que l'on est
dans le cadre d'une hérédité
morganienne étendue à la biologie
moléculaire du gène, tout facteur
héréditaire devient un facteur lié
à l'ADN (nucléaire ou mitochondrial). On
distingue alors:
- les maladies monofactorielles ou
à caractère monogénique
ou encore à caractère
mendélien: tous les symptômes de la
maladie sont reliés à un seul gène qui
se présente sous forme d'un allèle
morbide (à l'origine de la maladie) qui peut
être dominant ou récessif. Pour sortir de la
dichotomie dominance/récessivité on parle de
pénétrance de l'allèle
dominant qui peut plus ou moins s'exprimer chez un
hétérozygote, un allèle récessif
ne s'exprimant pas, sauf chez un homozygote. De nombreuses
maladies considérées comme autosomales
dominantes présentent une pénétrance
incomplète et souvent variable avec le sexe
(camptodactylie, brachydactylie, certaines formes de
cataracte congénitale, de microphtalmie ou
d'anophtalmie...). Parmi les maladies gonosomales on
distingue celles qui sont liées à des
gènes portés par la partie propre de l'Y,
celles qui sont liées à la partie propre de
l'X (maladies "liées au sexe") et celles qui sont
liées à des gènes situées dans
la partie commune à l'X et à l'Y et qui sont
qualifiées de pseudo-autosomales. Il ne faut pas
oublier qu'un même symptôme (par exemple
l'opacification du cristallin) peut être du à
de nombreuses maladies avec différents agents.
Ensuite, dans le cas d'une maladie génétique
reconnue, plusieurs formes génétiques peuvent
coexister (on connaît des cataractes
congénitales qui se transmettent sous les modes
dominant autosomique, récessif autosomique ou
récessif lié à l'X). Enfin
différentes formes, dues à des gènes
différents, peuvent se transmettre sous le même
mode héréditaire.
Pour voir une illustration des
caractères monogéniques vous pouvez par
exemple visiter la page www.ac-creteil.fr/svt/Doc/Doc1S/lego/lego.htm
où un collègue modélise les chromosomes
puis les gènes avec des briques de LEGO©...Les
gènes sont bien des particules localisées sur
un chromosome, chaque gène étant
associé à un caractère. Si les
gènes étaient des unités de fonction,
ils pourraient comporter plusieurs sous-unités sur
différents chromosomes...et leurs lois de
transmission n'auraient plus grand chose à voir avec
une répartition statistique plus ou moins
équiprobable dans les gamètes. Ces
caractères, toujours associés à des
gènes dans le cadre de la théorie
chromosomique de l'hérédité, mais
associés à plusieurs gènes en
interaction sont dits justement "polygéniques" mais
aussi à tort "non mendéliens".
- les maladies multifactorielles ou
à caractère polygénique
ou à caractère "non mendélien"
: plusieurs gènes sont associés aux
symptômes. Pour éviter de ranger dans ce groupe
toutes maladies héréditaires pour lesquelles
aucun gène n'a été (encore)
isolé, il faut bien comprendre que ce qui est en
cause ici c'est la ségrégation
indépendante des caractères lors de la
formation des gamètes. Pour ces maladies on ne remet
pas en cause l'hérédité morganienne, on
propose que le caractère soit associé à
plusieurs gènes (polygénisme) ou
bien qu'un même gène puisse conditionner
plusieurs caractères
(pléiotropie) ou enfin qu'il y ait des
interactions non allèliques (et donc du milieu) entre
gènes (épistasie).
- les maladies mitochondriales à
hérédité maternelle puisque l'on pense
que toutes les mitochondries sont transmises par
l'ovocyte.
- les maladies chromosomiques qui touchent des
portions de chromosomes suffisamment grandes pour être
détectées par des techniques de coloration
(caryotype). Il peut y avoir des monosomies, des trisomies
ou des délétions, des translocations...; ces
différentes anomalies pouvant se combiner.
|
1er exemple : le diagnostic d'une
anomalie chromosomique: la trisomie 21 (voir
page annexe) (travail
personnel)
2ème exemple: dépistage et diagnostic de la
phénylcétonurie.
(corrigé en cours)
(Exercices Belin p 207 ; vous vous aiderez d'une
page annexe sur la phénylcétonurie
(TD de 1èreS) et du cours de
1èreS).
|
Éléments de correction:
1. On considère que cette maladie est une
maladie génétique monofactorielle
présente sous deux allèles (+,
sain) et (ph, morbide). D'après les deux arbres
généalogiques on peut déduire que,
dans cette hypothèse, l'allèle morbide est
récessif par rapport à l'allèle sain
(des enfants (AII3, BII1 et BII3) sont nés malades
de parents sains; leurs parents portaient donc chacun
l'allèle morbide à l'état
caché; l'allèle morbide est donc dit
récessif).
La prévalence de la maladie
étant de 1 naissance sur 10.000 dans la population
européenne (nombre de nouveaux cas sur une
période donnée), on peut estimer que le
risque de porter l'allèle (fréquence
allèlique dans la population
européenne) pour un individu de cette
population est 1/100 (puisque deux individus porteurs de
sexe opposés ont un enfant malade avec un risque
de 1/100 x 1/100 = 1/10.000).
La liaison chromosomique du gène
peut être étudiée toujours dans le
même cadre simpliste d'une maladie
génétique monofactorielle. Le gène
n(est pas lié à la partie propre de l'Y car
un garçon malade n'a pas son père et ses
frères malades. De même, le gène
n'est pas lié à la partie propre de l'X car
la fille BII1 est malade d'un père sain (dans ce
cas, le père ne pourrait que lui avoir
donné un X porteur de l'allèle ph, et donc
être malade, ce qui n'est pas le cas). Donc le
gène étudié est associé
à une partie autosomale des chromosomes (autosomes
ou partie pseudo-autosomale des gonosomes). (Cependant
cette étude n'était pas nécessaire
puisque l'on vous disait dans le sujet que le gène
était situé sur le chromosome 12).
Comme AII2 n'est pas malade il n'a que 2/3 de risque
d'avoir l'allèle ph (être porteur) et donc
1/2 x 2/3 = 1/3 de risque de donner l'allèle ph
dans un gamète. Sa femme AII1 est issue de la
population européenne sans plus de
précision et peut donc donner l'allèle ph
avec un risque de 1/100. Ce qui fait un risque pour
l'enfant à naître d'être malade de 1/3
x 1/100 = 1/300.
Comme les deux parents BI sont
hétérozygotes et porteurs de
l'allèle ph ils peuvent le donner dans un
gamète avec une probabilité de 1/2 chacun.
ce qui fait 1/2 x 1/2 = 1/4 de risque pour l'enfant
à naître d'être malade. la
présence de frères et sœurs malades
déjà nés ne change pas ce
risque.
2. La technique des ASO (Allele Specific Oligoprobe)
permet de détecter des mutations ponctuelles (par
substitution d'une seule base de l'ADN). Elle est
utilisée ici pour le codon 280 où l'on
connaît une mutation ponctuelle (G changé en
A dans l'ADN au niveau du brin codant) conduisant
à un changement d'aa (Glu changé en Lys)
dans la protéine PAH dont nous étudions ici
le gène. On voit donc que les parents sont tous
deux hétérozygotes (un exemplaire de chaque
allèle), ce qui est une surprise pour la
mère (AII1) qui n'avait qu'un risque de 1/100 et
dans une moindre mesure pour le père, qui avait un
risque de 2/3. L'enfant AIII3 est homozygote
récessif avec une double mutation 280Lys//280Lys,
ce qui est aussi surprenant. Il a toutes les chances
d'être malade à la suite de cette mutation
double (voir les phénotypes sur le site de l'Inrp:
http:/www.inrp.fr/Acces/biotic/gpe/dossiers/phenylcetonurie/html/mutations.htm
) associé à une
phénylalaninémie très forte (voir
site INRP).
Pour la famille B les trois individus testés sont
homozygotes sains et donc de génotype
280Glu//280Glu pour ce site de mutation.
Avec la méthode du Southern Blot appliquée
sur des petits fragments de restriction (RFLP... que je
considère hors programme en tant que telle), le
sujet nous explique (mal) que la mutation
localisée au codon 408 (région c) et
caractérisée par un mutation ponctuelle
dans l'ADN par substitution de base (C en T pour le brin
codant) et dans la chaîne d'aa de la PAH un
changement d'aa (Arg en Trp), dégage un site de
restriction nouveau au voisinage de ce codon 408 pour
l'enzyme MspL. Au lieu donc d'obtenir un seul fragment
long de restriction de 23 kbases, on obtient dans le cas
d'un allèle muté un fragment de 13,5 et un
autre fragment de 9,5. Mais il faut savoir que ce petit
fragment n'est pas repéré sur
l'électrophorèse présentée
dans le sujet.
D'après le document 3 on peut donc affirmer que
les parents BI sont tous les deux
hétérozygotes, ce qui était connu et
que l'enfant à naître est homozygote
récessif, ce qui n'était probable qu'avec
un risque de 1/4.
Il est à noter que, contrairement à ce
qu'affirme la question du sujet, cette méthode
n'apporte pas beaucoup de certitudes. D'abord comme tout
geste technique, il est entaché d'incertitude,
bien difficile à chiffrer (erreurs de
manipulation, d'action des enzymes, des sondes... de
migration...). Ensuite, le fait de se focaliser sur trois
allèles peut se justifier par le fait que, dans
une famille atteinte d'une maladie
génétique, on considère qu'une
mutation dans le gène incriminé à
plus de chance de se transmettre dans la famille qu'une
autre mutation dans le même gène. Cependant,
ce n'est pas une certitude et il est possible que
d'autres allèles puissent aussi être
mutés (regardez à nouveau le tableau
des mutations sur le site de l'Inrp). Enfin, et
surtout, s'il est légitime, dans un exercice
scolaire de terminale, de raisonner à partir de
l'hypothèse simpliste d'une maladie
génétique monofactorielle, il n'est plus
acceptable que cela soit le cas dans des analyses
génétiques médicales
proposées à des patients. Pour une
ouverture sur les déterminismes de la
phénylcétonurie, voir page
annexe du cours de 1èreS.
|
Remarques:
Pour un début d'approche de génétique des
populations voir la loi d'HardyWeinberg dans l'ancien
cours de spécialité. Pour
les notions de épidémiologiques de prévalence
et de morbidité, voir une page
annexe sur les statistiques.
3ème exemple: la mucoviscidose, un maladie à
forte composante génétique: de la recherche d'une
causalité au diagnostic puis à l'évaluation du
risque. (travail personnel)
Belin p 191-193, et ancien cours
de TS spécialité partie 2.3
Remarque: ne pas oublier l'utilisation des antibiotiques
"suppresseurs de codons stop", qui ont été
utilisés avec succès chez certains malades atteints
de mucoviscidose (Pascale Fanen: «Un traitement
pharmacologique restaure le gène
déficient», La Recherche, 370,
décembre 2003, 26-27 d'après M. Wischanski et al.,
NEJM, 349, 1433, 2003 dont le résumé en anglais a
été ajouté dans l'ancien
cours de spécialité);
2.3 de la prudence et des limites des techniques
de transgénèse sur les êtres vivants (OGM)
* une transgénèse
réussie: la production d'insuline humaine par
des bactéries Escherichia coli génétiquement
modifiées. Exercice d'après Didier p 80-81.
Exercice et corrigé.
Cette transgénèse est à la fois le premier et
le plus probant de exemples d'application de la technologie de
l'ADN recombinant (génie génétique). Vous
noterez combien cette technique est complexe et le temps
nécessaire pour mettre au point la souche
génétiquement modifiée utilisée
actuellement.
(Un deuxième exemple, pourrait être celui de la
somatostatine, une hormone hypothalamique de 14 aa intervenant
dans la croissance chez l'homme).
* de nombreuses transgénèses ont
été réalisées d'abord chez des
procaryotes, puis chez des eucaryotes notamment sur les
végétaux (riz, maïs...), pour
l'amélioration du rendement des cultures mais aussi pour la
recherche. On est actuellement dans une phase expérimentale
d'utilisation des OGM à très grande
échelle.
Les transgénèses animales sont plus difficiles que
celles réalisées sur des plantes mais fort
explorées.
|
Nathan p 88-89 (Belin p 166-183):
1 - Comparer la transgénèse et les OGM; que
recouvrent ces notions ?
2 - Pourquoi parle-t-on surtout d'OGM du règne des
plantes ? Quelles en sont les spécificités
?
3 - Quel est le rôle précis
d'Agrobacterium tumefaciens ? (Belin p
166-169)
4 - Belin QCM n°2 p 184
|
2.4 des limites éthiques à la transgenèse
humaine (HGM)
Les transgénèses humaines sont
habituellement à but thérapeutique et sont donc
qualifiées de thérapie génique. Elles en sont
à leur balbutiements.
|
Nathan p 96-97 (Belin p 194-197), bioéthique
dans l'ancien cours de spécialité avec
notamment la déclaration
universelle sur le génome humain et les droits de
l'homme (UNESCO, 1997):
1 - En quoi le terme d'HGM est-il faux ? Donner une autre
définition de la thérapie génique en
utilisant le terme de greffe.
2 - En quoi la transgénèse humaine est-elle
complémentaire des recherches sur les cellules
souches ?
3 - Rechercher sur internet la loi de 1994 autorisant les
recherches sur les cellules somatiques
4 - Qu'est-ce que l'éthique ? Que veut dire le
terme de limite scientifique ? Que pensez-vous de titres
de presse du style "l'homme transgresse les lois du
vivant" ? Que veut dire le terme de limite éthique
?
|
|
Exercices
corrigés:
|
- Nathan n°3 p 104
(Exercice typique de baccalauréat de type
spécialité - partie II b)
|
Sujet
Un couple ayant un enfant atteint de
drépanocytose voudrait savoir si leur
futur bébé sera également
atteint de cette maladie. Le
drépanocytose est une
maladie qui touche le
sang, elle est due à une anomalie
de la ß globine. Le gène de la
ß globine possède un allèle
sain ßA et un
allèle délétère
ßS. L'ADN des parents, de leur
enfant et de leur fœtus est
prélevé et coupé par
l'enzyme MstII. Une électrophorèse
est réalisée pour séparer
les différents fragments obtenus qui sont
identifiés par une sonde
spécifique marquée.
Question:
En utilisant l'ensemble de ces données,
expliquez
si le futur
bébé sera atteint de
drépanocytose.
Document1: "gène"
(séquence
très partielle) de la ß
globine
Document 2 : carte de restriction des
sites MstII du chromosome
11 au voisinage du gène de la
ß globine: position
d'appariement de la sonde utilisée dans
le document 3
NB: la sonde
repère aussi bien les fragments 0,2kb que
1,2 kb que 1,4 kb.
Document 3: arbre
généalogique de cette famille
(voir texte) et résultats de
l'électrophorèse (id)
les résultats
étant présentés à la
verticale en dessous du figuré de chaque
membre de la famille.
Remarque:
Le texte du sujet est critiquable. Si la
drépanocytose est certainement le
meilleur exemple de liaison entre le
génotype et le phénotype on
connaît de très nombreux
allèles pour la chaîne ß de
l'hémoglobine (voir ancien
cours de
spécialité)
donnant lieu à des hémoglobinoses
et thalassémies aux facteurs
génétiques non
déterminants. La question n'est pas
rédigée en français correct
et laisse pantois. Ce que l'on demande
(malheureusement) en terminale est une analyse
probabiliste simpliste issue de l'arbre
généalogique qui peut être
améliorée par une analyse
génétique précise
(compliquée et coûteuse) mais qui
ne sort pas du paradigme posé d'une
maladie monofactorielle associée à
un gène unique sous la forme d'un
allèle morbide
récessif.
|
|
Corrigé
On pose une hypothèse de départ:
nous connaissons une maladie
génétique monofactorielle (la
drépanocytose) qui est due à une
mutation ponctuelle (document 1, base
n°17, A changé en T sur le brin 5')
d'un unique gène associé à
la chaîne ß de l'hémoglobine.
Comme ce n'est pas dit dans le texte, c'est le
début du gène de la ß
globine qui est représenté dans le
document 1 et la mutation ponctuelle conduit
à une substitution d'un acide
aminé en position 6 : la valine (Val)
à la place de l'acide glutamique (Glu).
Cette mutation conduit à un
allèle récessif (ßS)
par rapport à l'allèle non
muté (ßA) (la
récessivité doit être
établie à partir du document 3
puisqu'une fille malade est née de deux
parents sains, porteurs de l'allèle
à l'atat caché). Tous les
symptômes de la maladie semblent pouvoir
être rapportés à la
présence de cette hémoglobine
anormale qui fixe moins bien le dioxygène
aux basses pressions et se polymérise en
donnant des "baguettes" assez rigides qui
déforment les hématies en
faucilles qui se bloquent dans les capillaires.
Les malades sont ceux qui possèdent deux
allèles ßS.
L'analyse de l'arbre
généalogique (document 3) en
utilisant le formalisme habituel (on porte de
part et d'autre d'une double barre de fraction
les allèles portés par chaque
paire homologue) et les statistiques donnant la
proportion théorique des
gamètes (que l'on peut rapporter
à la première loi attribuée
à Mendel: loi de pureté des
gamètes) permet de dire que l'enfant
attendu présente un risque de 1/4
d'être atteint (ßS//ßS), un
risque de 1/4 d'être sain
(ßA//ßA) et un risque d'1/2
d'être porteur mais sans symptôme ou
porteur sain (ßA//ßS). En effet,
chaque parent, de génotype
(ßA//ßS) donne avec une égale
probabilité (1/2) un gamète
ßS ou ßA, ce qui fait 1/4 pour
chaque fécondation.
L'analyse génétique
utilisant l'enzyme de restriction MstII est un
peu originale dans le sens où le site de
restriction situé en position 14 depuis
le début du gène de la ß
globine (en partant de l'extrémité
5'; en 17 sur le brin 3'->5') est
supprimé lors de la mutation
(allèle ßS), ce qui fait qu'un
allèle sain (ßA) est
repéré par deux fragments de
restriction (1,2 et 0,2 kb) alors que
l'allèle muté est
repéré par un seul fragment de
restriction (1,4 kb). Ainsi les génotypes
proposés pour les parents et les
sœur de l'enfant attendu sont donc
confirmés. Le génotype de
l'enfant étant ßA//ßA
puisqu'il possède uniquement des
fragments de restriction à 1,2 et 0,2 kb.
Il est donc sain et non porteur de
l'allèle ßS.
Ce diagnostic est certain, aux erreurs
de manipulation et d'expérience
près (en effet la sonde peut mal se
fixer, l'enzyme de restriction couper de
façon erronée ou encore les
fragments mal migrer...). De plus il est
essentiel de noter que l'on reste, avec cette
méthode dans le paradigme dominant avec
l'hypothèse de départ
(justifiée) de la maladie
génétique monofactorielle. Ceci
n'est bien qu'un exercice scolaire. Il ne faut
pas lui demander de répondre à des
questions biologiques que tout
élève de terminale ne manquera pas
de se poser par ailleurs.
|
- Nathan n°3 p 86
avec le document 3 p 79 pour aider à la
compréhension (attention, ceci n'est pas un
exercice de type baccalauréat mais juste un
exercice de compréhension - exercice de
formation)
|
Sujet
Dépistage moléculaire d'une
mutation
Les propriétés de
renaturation de l'ADN sont mises à profit
dans de nombreuses méthodes de
dépistage moléculaire de mutations
ponctuelles d'un gène. L'une de ces
méthodes est l'ARMs (Amplification
Refractory Mutation System). Elle consiste
à amplifier, par ACP *),
un court fragment d'un gène, au sein
duquel peut se trouver l'une des mutations
connues de celui-ci, dont on se propose de
dépister la présence
éventuelle. D'un côté, on
utilise une amorce "commune" et, de l'autre,
deux amorces (document 1), présentant en
3' une base complémentaire, soit de la
base présente sur la séquence
standard du gène (amorce standard), soit
de la base présente sur la
séquence mutée (amorce
mutée). Par ailleurs, ces deux amorces
sont allongées par une séquence
additionnelle polyN ou polyN' de longueurs
différentes, de sorte que, selon que la
mutation est présente ou absente,
l'ACP* donnera des fragments
de longueur différente.
On teste, par cette méthode, la
présence ou l'absence d'une mutation,
notée a, d'un gène, responsable
d'une maladie récessive (le gène
fonctionnel sera noté A). Les amorces
utilisées ont une longueur de 25
nucléotides, les queues polyN et polyN'
ont des longueurs respectives de 10 et 30
nucléotides. les résultats sont
indiqués dans le document 2.
Document 1
(amélioré)
|
- une amorce commune amplifie le
brin + (s'hybride au brin -).
- l'amorce standard amplifie le brin
- s'il n'est pas muté et
l'amorce mutée amplifie le brin
- qu'il est muté; chacune de ces
deux amorces est pourvue d'une queue
polyN ou polyN' de taille
différente.
- les fragments d'ADN
amplifiés par ACP*
seront plus longs si l'ADN testé
est muté et plus courts s'il ne
l'est pas, la taille et la
différence de taille
étant visualisables sur un gel
d'éléctrophorèse.
|
|
Taille des fragments
de ACP*
(en paires de
bases: pb)
|
A
(est
atteint)
|
B
(est atteint)
|
C
(est atteint)
|
D
(non atteint,
est parent d'un
atteint)
|
E
(non atteint,
est parent d'un
atteint)
|
F
(non
atteint)
|
G
(non
atteint)
|
|
130 pb
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
110 pb
|
|
|
|
-
|
-
|
|
-
|
Document 2. Résultats du gel de
contrôle des fragments
d'ACP* obtenus pour
chaque individu.
|
Questions:
a. Indiquez la taille du fragment du gène
amplifié par PCR (ACP*).
b. Compte tenu de ces résultats et des
informations suivantes, précisez autant
que possible le génotype de chaque
individu pour la mutation testée et pour
le gène étudié.
remarques:
- la molécule d'ADN comportent deux brins
orientés du fait de l'accrochage des
nucléotides entre le sucre d'un
nucléotide et le groupement phosphate du
nucléotide précédent. Il y
a donc deux brins antiparallèles
qualifiés de brins + et - ou 3'->5' et
5'->3'
- les n°s 3' et 5' désignent les
carbones du désoxyribose (ceux de la base
de chaque nucléotides sont
numérotés sans les '); le
groupement phosphate de chaque nucléotide
est accroché en C5' et la base en
C1'.
-l'ADN polymérase est un complexe
enzymatique qui travaille toujours (par ajout de
nucléotides) dans le sens
5'->3'
|
|
Corrigé
a) le gène fait 100 paires de bases (en
comptant depuis le début de chaque
amorce).
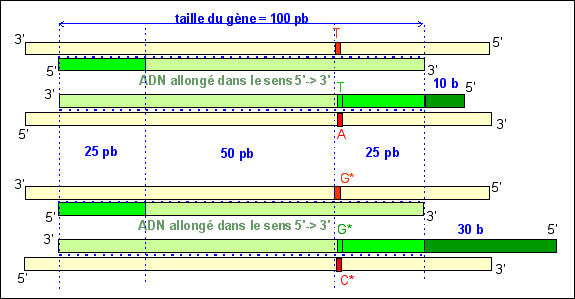
b) Il suffit de lire le génotype: un
fragment de 130 pb signifie une queue poly N' de
30 bases et donc un allèle muté a;
un fragment d'ADN de 110 pb signifie une queue
poly N de 10 bases et donc un allèle non
muté A;
A, B et C sont a//a, D, E et G sont a//A et F
est A//A.
|
- Nathan n°6 p 72
(exercice pouvant être posé en partie IIb
(de spécialité) mais dont
l'énoncé est inhabituel et
déroutant; son intérêt réside
dans le fait qu'il présente à la fois une
technique de transgenèse et une analyse
morganienne de
l'hérédité)
|
Sujet
Concevoir des croisements
La génétique mendélienne
est avant tout une science expérimentale,
fondée sur l'observation des
résultats de croisements. Le choix
pertinent de ces croisements est donc
décisif.
Document 1: garniture chromosomique de la
drosophile mâle (XY et 3 paires
d'autosomes dont une paire ponctiforme) et
femelle (XX et 3 paires d'autosomes dont une
paire ponctiforme).
Document 2: "carte génétique": le
chromosome II porte le gène
associé à la forme de l'aile
(vestigiale); le chromosome X porte le
gène associé à la couleur
de l'œil (blanc) et un transgène
(associé à la synthèse
d'une protéine fluorescente); ces deux
gènes sont distants de 3,7 unités
de recombinaison.
Document 3: souches pures de drosophiles (pour
les mutations) disponibles pour les
croisements:
On note que le transgène n'est
inséré qu'à un seul
exemplaire dans les souches mutées et que
le transgène n'est exprimé que
chez les femelles...
|
nom de la souche
|
sauvage
|
A
|
B
|
C
|
D
|
E
|
F
|
G
|
|
mutation ou transgenèse
|
aucune
|
abdomen fluorescent chez les
femelles
|
œil blanc
|
œil blanc et abdomen des
femelles fluorescent
|
ailes vestigiales
|
ailes vestigiales et abdomen
des femelles fluorescent
|
œil blanc et ailes
vestigiales
|
abdomen des femelles
fluorescent, œil blanc et ailes
vestigiales
|
Questions:
À partir de la mise en relation des
informations apportées par les documents,
déterminez, en justifiant votre
réponse, les croisements à
effectuer pour confirmer la localisation
chromosomique relative des gènes
étudiés.
Les résultats qualitatifs et quantitatifs
de chaque croisement sont attendus, en admettant
un effectif théorique de 2000 individus
à chaque
génération.
|
|
Corrigé
On a donc comme hypothèse 3
gènes:
- un gène lié au chromosome 2
(autosomal) et associé au
caractère "forme de l'aile"
présent sous deux formes: un
allèle récessif "vestigial"
(noté vg) par rapport à un
allèle sauvage (ailes longues) dominant
(noté +)
- un gène lié au chromosome 1 ou X
(gonosomal) et associé au
caractère "couleur de l'œil"
présent sous deux formes: un
allèle récessif "œil blanc"
(noté w pour white) un allèle
sauvage (œil rouge) dominant (noté
+)
- un transgène que l'on place sur le
chromosome X (1) (noté
*) et qui ne
s'exprime (sous la forme d'une protéine
fluorescente synthétisée par
certaines cellules) que chez les femelles (par
un abdomen fluorescent). Le sujet précise
que l'on insère le gène en un seul
exemplaire. On peut imaginer que l'on a donc
uniquement des femelles avec le transgène
ou que l'on possède à la fois des
mâles et des femelles. Si l'on veut que le
gène reste toujours à un seul
exemplaire on ne peut donc faire des croisements
qu'avec des femelles transgèniques.
Pour justifier par des croisements judicieux
de la position des gènes proposés,
on doit suivre les étapes suivantes:
1 - montrer que le gène associé
à la "couleur de l'œil blanc" est
situé sur le chromosome X
2 - montrer que le gène associé
à "la forme des ailes de type vestigiale"
n'est pas situé sur le chromosome X et
est donc indépendant du premier
gène
3 - montrer que le transgène et le
gène associé à "la couleur
de l'œil blanc" sont distants de 3,7
unités de recombinaison.
Etape1
Un gène "lié au sexe",
c'est-à-dire "porté" par la partie
propre de l'X, présente lors de la
première génération (F1)
une différence entre le croisement d'un
mâle de phénotype muté
(correspondant à l'allèle
récessif) avec une femelle de
phénotype sauvage et le croisement
réciproque d'un mâle sauvage avec
une femelle de phénotype muté.
|
(souche B) femelle w//w
x +//y mâle (souche
sauvage)
toutes les femelles à
yeux
rouges et tous les
mâles ont les yeux
blancs
|
(souche sauvage) femelle
+//+ x w//y mâle (souche
B)
tous les descendants ont les yeux
rouges
|
SINON
si le gène associé au
caractère "couleur des yeux
blancs" n'était pas lié
au sexe (à la partie propre de
l'X) il y aurait 100% de descendants
à yeux rouges pour les deux
croisements
|
|
sur 2000 descendants en supposant un
sex ratio de 1/1, il y aura 1000
femelles à yeux rouges et 1000
mâles à yeux blancs
|
sur 2000 descendants il y aura 2000
descendants à yeux rouges
(100%)
|
Étape 2
Le même raisonnement que
précédemment est fait pour le
gène associé au caractère
"forme des ailes vestigiales".
|
(souche D) vg//vg x +//+
(souche sauvage)
tous les descendants ont les ailes
longues (non vestigiales)
(100%)
|
SINON
si le gène associé au
caractère "forme des ailes
vestigiales" était lié au
sexe on aurait deux types de
croisements avec des résultats
différents:
|
|
sur 2000 descendants d'un individu
de la souche sauvage avec un individu
de la souche D, quelque soit les sexes,
il y aura 2000 descendants aux ailes
longues (100%)
|
femelle vg//vg x +//y
mâle
toutes les femelles ont les ailes
longues et tous les mâles ont les
ailes vestigiales
|
femelle +//+ x vg//y mâle
tous les descendants ont les ailes
longues
|
|
sur 2000 descendants en supposant un
sex ratio de 1/1, il y aura 1000
femelles à ailes longues et 1000
mâles à ailes
vestigiales
|
sur 2000 descendants il y aura 2000
descendants à ailes longues
(100%)
|
Comme le gène associé au
caractère "forme des ailes vestigiales"
n'est pas lié au sexe il est donc
lié à un autosome (mais on
ne peut pas connaître le n° du
chromosome à moins d'avoir un autre
gène que l'on saurait être
lié à ce chromosome) et il est
donc indépendant du gène
associé à "la couleur de
l'œil blanc" qui est lié à
l'X.
Étape 3
Pour estimer la liaison entre le
transgène et le gène
associé à "la couleur de
l'œil blanc" il est habituel d'utiliser un
test-cross.
(souche C) femelle w *
//w x +//y mâle (souche
sauvage)
|
|
+
|
y
|
|
w *
|
w
*//+
|
w *//y
|
|
w
|
w//+
|
w//y
|
on utilise ensuite les femelles
à yeux rouges et à
abdomen fluorescent hybrides
obtenues
(hybride de F1) femelle w * //+
x w//y mâle (souche B)
|
|
w*
|
+
|
w
|
* +
|
|
w
|
w*//w
|
+//w
|
w//w
|
+*//w
|
|
y
|
w*//y
|
+//y
|
w//y
|
*+//y
|
|
,
|
48,2%
963
|
48,2%
963
|
1,9%
37
|
1,9%
37
|
96,3%
1926
|
3,7%
74
|
|
SINON
toute autre proportion indiquerait
une distance erronée.
|
|
sur 2000 descendants en supposant un
sex ratio de 1/1, il y aura
- 1926 descendants de type parental
(non recombinés), avec un sex
ratio de 1/1, cela fait 963
mâles (dont la moitié de
phénotype à yeux blancs
et autant de phénotype à
yeux rouges), et 963 femelles dont la
moitié de phénotype
à yeux
blancs et abdomen
fluorescent et autant de
phénotype à yeux
rouges.
-74 descendants de
type
recombiné se
répartissant en 37 mâles
(dont la moitié à yeux
blancs et l'autre moitié
à yeux rouges) et 37 femelles
(dont la moitié sont à
yeux blancs et l'autre moitié
à yeux
rouges et abdomen
fluorescent.
Si l'on somme les phénotypes
on obtient:
- 499 (481+18) mâles à
yeux blancs
- 499 (481+18) mâles à
yeux rouges
- 481 femelles à yeux blancs et
abdomen fluorescent
- 18 femelles à yeux blancs
- 481 femelles à yeux rouges
- 18 femelles à yeux rouges et
abdomen fluorescent
|
Je ne suis pas
sûr d'avoir correctement exploré
toutes les possibilités... vu l'urgence
je vous propose cette interprétation non
définitive.
|
- Belin n°4 p
208-204 (Attention cette exercice n'a pas la
forme d'une question de type spécialité -
partie II b - mais il constitue un exercice
d'entraînement)
|
Sujet
Diagnostic prénatal
On dispose de 3 enzymes de restriction (doc. 1).
Ces enzymes découpent l'ADN en des
endroits précis, appelés sites de
restriction, ce qui permet à la
bactérie qui les produit dans son
cytoplasme de digérer l'ADN de
bactériophages qui tentent de
l'infecter.
On étudie deux allèles du
même gène dont les séquences
sont décrites ci-dessous (doc. 2).
L'allèle "s" correspond à
l'allèle "sain"(présent chez les
individus sains), et l'allèle "m"
à l'allèle "malade" (morbide,
présent chez les individus malades).
L'arbre généalogique
présenté (doc.3) est celui d'une
famille dont les deux parents sains ont
donné naissance à un enfant
atteint d'une maladie génétique
récessive et à un enfant sain.
Lors d'une troisième grossesse, tous les
sujets sont testés par la méthode
de Southern (digestion par enzyme de restriction
+ électrophorèse + transfert sur
nitrocellulose + mise en contact avec une sonde
radioactive + autoradiographie).
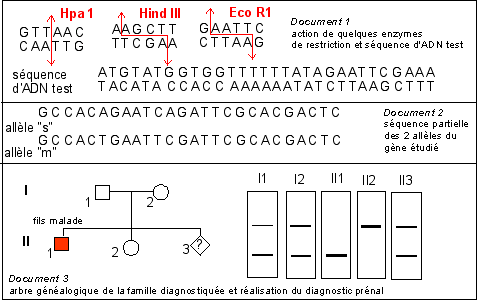
Questions:
a - Déterminer quelle(s) enzyme(s) de
restriction peuvent agir sur la séquence
proposée. Préciser sur quel(s)
site(s) ces enzymes peuvent agir.
b - À l'aide du code
génétique,
déterminer pourquoi cette séquence
correspond à un allèle malade.
c - Décrire les effets de la ou des
enzymes déterminées à la
première question sur l'allèle "s"
et sur l'allèle "m". Évaluer la
taille (en nucléotides) des fragments
obtenus.
d - Schématiser le résultat de
l'électrophorèse des fragments
obtenus après digestion par cette (ces)
enzyme(s).
e- Établir le diagnostic prénatal
de l'enfant II3. Justifier ce résultat
à partir des
autoradiographies.
|
|
Corrigé
a) seule EcoRI peut couper l'ADN de la
séquence du seul allèle m (mais
pas s)
b) en supposant que la séquence des
bases est celle du brin codant (non transcrit)
on a donc un ARNm
de séquence identique base à base
à celle de l'ADN avec U à la place
de T ; grâce au code
génétique on associe alors les aa
à chaque triplet (codon) de l'ARNm:
|
|
Thr
|
|
Ser
|
|
|
|
|
|
aa
"s"
|
|
|
ACA
|
|
UCA
|
|
|
|
|
|
ARNm
"s"
|
|
GCC
|
ACA
|
GAA
|
TCA
|
GAT
|
TCC
|
GCA
|
CGA
|
CTC
|
ADN allèle sain
"s"
|
|
GCC
|
ACT
|
G*AA
|
TTC
|
GAT
|
TCC
|
GCA
|
CGA
|
CTC
|
ADN allèle
morbide"m"
|
|
|
ACU
|
|
UUC
|
|
|
|
|
|
ARNm
"m"
|
|
|
Thr
|
|
Phe
|
|
|
|
|
|
aa
"m"
|
|
1 2
3
|
4 5
6
|
7
*
8 9
|
10 11
12
|
13 14
15
|
16 17
18
|
19 20 21
|
22 23
24
|
25 26
27
|
n° des bases
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
n° des codons
|
sur fond bleu la
séquence reconnue par l'enzyme de
restriction EcoRI
l'étoile (*)
indiquant le site de coupure
On observe donc trois substitutions de bases
entre les deux séquences:
* en position 2 (2ème codon) une
adénine (A) est changée en thymine
(T) mais comme le code génétique
est redondant les codons ACA et ACU codent pour
le même acide aminé : la
thréonine (Thr); cette mutation est donc
silencieuse (neutre).
* en position 4 (4ème codon) une cytosine
(C) et une adénine (A) sont
remplacées par une thymine (T) et une
cytosine (C) respectivement. Le codon UCA
associé à la sérine (Ser)
est changé en UUC associé à
la phénylalanine (Phe); il y a donc
changement de base et donc un peptide
différent qui est peut-être
à l'origine des symptômes de la
maladie dans les cellules exprimant le
gène étudié.
c) L'allèle "s" présente donc
un "gène" de 27 paires de bases et
l'allèle "m" coupé par l'enzyme de
restriction EcoRI deux fragments de restriction
de 7 et de 20 paires de bases.(on parle de
paires de bases car la molécule d'ADN est
double brin, même si un seul brin a
été représenté
ici).
d) L'ADN étant chargé
négativement à pH basique (on
travaille à environ pH = 9) les fragments
d'ADN déposés au pôle "-"
migrent vers le pôle "+" d'autant plus
loin et plus vite que les fragments de
restriction sont plus petits et plus
chargés électriquement. Les
fragments isolés dans le Southern Blot
sont des ADN monobrins et donc notés en
bases et non en paires de bases.
|
position (ou taille) en bases
|
s
|
m
|
s//s
|
s//m
|
m//m
|
|
pôle -
|
.
|
.
|
.
|
.
|
.
|
|
27 bases
|
-
|
|
-
|
-
|
|
|
20 bases
|
|
-
|
|
-
|
-
|
|
.
|
,
|
,
|
,
|
,
|
,
|
|
7 bases
|
|
-
|
|
-
|
-
|
|
pôle +
|
.
|
.
|
.
|
.
|
.
|
Les seuls représentés dans la
représentation du Southern blot sont les
fragments 20 et 27. On peut émettre
l'hypothèse que la sonde de marquage des
fragments ne peut pas se fixer au fragment de 7
bases.
e) D'après l'arbre
généalogique l'enfant II3 attendu
par deux parents hétérozygotes
(puisque l'allèle m est récessif
par rapport à l'allèle s puisque
un enfant (II1) est né malade de deux
parents apparemment sain qui portaient donc
chacun l'allèle à l'état
caché) a un risque de 1/4 d'être
m//m ou s//s et un risque 1/2 d'être m/s.
D'après le diagnostic prénatal
réalisé par test
génétique l'enfant II3 est
hétérozygote (m//s) puisqu'il a
notamment le même profil de restriction
que ses parents. Ce résultat apporte une
certitude aux erreurs de manipulation et
d'activité des produits utilisés
près. De plus ce résultat ne peut
être pris en compte que dans le cadre de
la théorie présentée au
début: l'hypothèse d'un unique
gène et de deux allèles connus,
séquencés....
|
|
Remarques:
Dans le cadre de cette année de
terminale, où vous découvrez la philosophie, il est
plus que temps de se poser la question du sens du mot information
génétique et de la raison de son emploi. Voici l'avis
documenté d'un grand mathématicien;
à ce sujet on peut aussi lire l'article
"forme" de Jean Petitot dans l'Encyclopedia Universalis ou quelques
mots sur la page des modèles
de ce site.
|
extrait de Un protée* de la
sémantique : l'information ,
René Thom, 1973, 6.
Écrit à la demande de J. d'Ormesson pour un
colloque de l'Unesco, Venise 28-31 mai 1973.)
Mots soulignés et colorés
par moi, note sur Protée
ajoutée.
|
|
1.6. Emplois du terme information en Biologie.
Il s'agit ici, essentiellement, de l'information «
génétique » d'une
espèce vivante, telle qu'on la trouve dans le
« dogme central » de Watson-Crick : «
l'information génétique d'un être vivant
est codée dans la composition en nucléotides
de son ADN ». Deux interprétations peuvent
ici être proposées. La première regarde
l'œuf comme un message émis par le parent
donneur (Y) ; le récepteur est alors l'embryon issu
de l'œuf lui-même ; on doit donc
considérer que le message, ici, devient son propre
récepteur ; c'est là une façon de voir
qu'il est difficile de soutenir. La deuxième
interprétation a été défendue
par quelques théoriciens de l'information en
Biologie, comme Dancoff et Quastler (note
1) : elle regarde l'organisme vivant comme le donneur
(X) de l'information, le destinataire (Y) est l'observateur
lui-même. Plus exactement, on considérera toute
morphologie naturelle comme un message émis par une
source fictive (Y) à l'adresse du savant (X), qui est
le demandeur. On retrouve ainsi une très vieille
idée : Dieu nous parle à travers les
apparences du monde, et il nous appartient de
déchiffrer son langage. Bien entendu, cette
manière de voir soulève de graves
problèmes ontologiques. Si on veut y voir plus qu'une
poétique métaphore, on est amené
à se demander si on peut encore faire une place
spéciale à la Sémiologie : si toute
forme physique ou naturelle est un message venu de Dieu,
pourquoi vouloir réserver une place spéciale
aux messages humains? De plus, dans tous ces exemples, la
finalité du procès n'apparaît pas
clairement, sauf à postuler l'intérêt
qu'il y a pour l'homme à comprendre ce qu'il voit.
Dans l'exemple (cité par M. Carreras dans son
intervention) « L'information fournie par les rayons
X après traversée du corps du patient
», il n'y a ambiguïté ni sur le donneur (le
corps du patient), ni sur le demandeur-récepteur
(l'observateur), ni sur la finalité globale du
processus (interprétation diagnostique). La seule
ambiguïté concerne le contenu signifié du
mot information, que pour ma part je réduirais
à la forme du message reçu sur la plaque
sensible, et interprété par l'observateur.
Dans les emplois scientifiques, en Biologie notamment,
où il y a effacement soit du demandeur, soit du
donneur-source, la malhonnêteté, si
malhonnêteté il y a, n'est plus que d'ordre
intellectuel. L'emploi du mot information, dans le cas de
« l'information génétique »,
témoigne de la situation psychologique suivante. Dans
le déroulement immensément complexe des
processus morphogénétiques de l'embryologie,
la Biologie Moléculaire a mis en évidence un
mécanisme important, la synthèse des
protéines. La tendance naturelle du
spécialiste est alors de dire que cette étape
est l'étape essentielle, les autres étapes
devant en être de simples conséquences. Le
mot information, en pareil cas, sert manifestement à
dissimuler l'ignorance quasi totale où nous sommes de
préciser ces autres mécanismes
prétendument subordonnés, tout en apportant
par la connotation d'intentionnalité du mot
information une caution implicite au finalisme qui sous-tend
toute pensée biologique. En ce
sens, l'information, c'est la forme
obscure de la causalité.
note(1)«
Autrement dit, la
théorie de l'information sous sa forme classique de
théorie de l'information transmise peut être
appliquée aux systèmes biologiques en
considérant qu'il existe une transmission
d'information entre l'organisme et l'observateur suivant la
méthode Dancoff et Quastler ainsi que nous l'avons
indiqué au chapitre 5 ; mais dans cette transmission
nous devons considérer chaque élément
ordonné ou organisé (macromolécules,
organelles, cellules, organismes) comme la sortie d'une voie
de communication, dont l'entrée est une source que
nous n'avons pas besoin de
spécifier. »
H. Atlan, L'Organisation biologique et la Théorie de
l'information, Hermann, Paris, p. 255.
* Protée: vieillard de la
mer dans la mythologie grecque qui avait le pouvoir de se
métamorphoser à volonté et qui, bien
qu'il connût le passé, le présent et
l'avenir, refusait de les révéler à
moins que celui qui l'avait capturé dans son sommeil
ne tienne bon et ne l'oblige enfin, une fois Protée
revenu à sa forme première, à
révéler la vérité.
|
retour texte
Pour se rendre compte des difficultés de la
notion d'information génétique, de programme
génétique ou encore de code
génétique, il suffit de voir comment les
média se sont emparés de ces termes et les ont
mélangés. Les difficultés sont patentes pour
tout collègue qui souhaite enseigner cette culture aux
élèves de 1èreES par exemple dans le cadre de
leur chapitre "du génotype au phénotype". Le programme
à ce sujet et on ne peut moins clair: "Cette partie de
programme s'appuie sur l'universalité de structure
et de fonction de la molécule d'ADN
étudiée en classe de seconde. Elle
précise, dans un premier temps, les mécanismes
biologiques assurant l'expression de l'information
génétique. Par la suite, à partir de quelques
exemples, elle appréhende la notion de complexité des
relations entre génotype et phénotype.(...) La
séquence des nucléotides dans l'ADN gouverne la
séquence des acides aminés dans la protéine
selon un système de correspondance, le code
génétique. "
Le code génétique est bien un code, une correspondance
entre un triplet de riibonucléotides de l'ARN et des aa
transportés par des ARNt spécifiques (voir
cours de
1èreS). Le code ne vient pas de
l'ADN mais des ARNt. Son universalité, c'est-à-dire
l'universalité des mécanismes de traduction,
dépend donc des ARNt.
Lorsque l'on insère un fragment d'ADN issu d'une cellule
étrangère (autre lignée cellulaire, autre
individu de la même espèce, ou d'une autre
espèce...) il existe une très grande
variété de causes qui font que cet ADN n'a pas la
même fonction que dans la cellule d'où il est issu. Il
peut d'abord être détruit, ce qui est le cas
général (c'est la raison de la présence des
enzymes de restriction dans les bactéries). Grâce
à des artifices (vecteur viral d'insertion) ou naturellement,
dans des cas rares, il s'insère dans le génome
hôte. Enfin, dans des cas extrêmement rares il peut
être exprimé, c'est-à-dire conduire à un
ARN ou à une protéine. Mais à quoi peut servir
cette protéine dans cette cellule hôte ? Affirmer que le
gène a la même fonction, consiste à rester au
niveau de l'expression moléculaire mais certainement pas
s'intéresser au phénotype. Certes, on arrive à
faire produire de l'insuline à une bactérie ou des
toxines bactériennes à une cellule du parenchyme de
réserve caulinaire d'un plant de pomme de terre, mais la
fonction de ce gène est mesurée à
l'utilité pour l'homme et non pour l'organisme qui, au mieux,
tolère ce parasitisme génique. Dire qu'il y a
universalité de la fonction de la molécule d'ADN
signifie uniquement affirmer que partout les protéines sont
synthétisées à partir de l'ADN puis de l'ARN. Ce
qui est déjà remis en cause actuellement. Par exemple
on connaît des antibiotiques qui sont de petits peptides (avec
des acides aminés L et D et des structures cycliques...) et
qui sont synthétisés par une voie différente de
la synthèse ADN-ARN-ribosomes et qui pourraient bien
constituer une voie alternative pour certains peptides. Pour la
gramicidine S bactérienne ce sont de gigantesques enzymes
(plus de 15.000 aa; la plus longue chaîne polypeptidique
connue...) - les peptide synthétases non ribosomiques ou NRPS
(Non Ribosomial Peptide Synthetase) - qui s'en chargent. Les NRPS ont
été étudiés (discrètement !)
depuis les années soixante. Les progrès de la biologie
moléculaire ont maintenant permis de couper ces énormes
molécules en modules actifs et, en insérant leurs
gènes dans des cellules procaryotes hôtes, leur faire
synthétiser des peptides antibiotiques "à façon"
(Demain, des antibiotiques à façon ?, Mohamed
Marahiel, Nadine Kessler et Uwe Linne, 2003, La Recherche,
370, décembre 2003, p 54-58).
Pour ce qui est des notions de programme
génétique j'ai déjà essayé
ailleurs d'en cerner les limites (voir commentaires
du programme de seconde). Voici juste une
petite citation de René Thom à ce sujet :
|
extrait de "La notion de
programme en biologie" (René Thom,
1984f5)
I Introduction : La théorie en biologie
Depuis qu'à partir du 16e siècle, on a
commencé à ouvrir les cadavres afin d'en
scruter les organes internes, l'explication biologique a
pris un tour résolument techniciste. On assimile
l'organe à un outil, lequel, construit par l'homme en
vue d'une fin connue, ne présente aucun obstacle
à l'intelligibilité. Dès Harvey, ce
point de vue a obtenu d'indiscutables succès. Il est
indéniable que le coeur est une pompe qui refoule le
sang dans ces canalisations que sont nos vaisseaux. De
même, les poumons se comportent (sur le plan
mécanique) comme un soufflet (mais ici
l'interprétation mécaniste laisse de
côté la fonction physiologiquement essentielle
d'échange gazeux entre l'air et le sang). Le
squelette (os + articulations) permet une
interprétation mécanique immédiate.
Toutes ces descriptions ont abouti à la
théorie cartésienne de l'Homme animal-machine,
et ce n'est guère qu'avec le plus noble de nos
organes (le siège de l'âme), à savoir le
cerveau, que l'imagerie techniciste est quelque peu
restée courte. Mais, avec la naissance
quasi-simultanée, vers 1950, des ordinateurs et de la
Biologie Moléculaire, cette imagerie a connu son plus
récent et peut-être son plus glorieux avatar.
On a fait de l'ADN des chromosomes, du génome,
l'analogue du programme qui régit le fonctionnement
d'un ordinateur. Cette dernière interprétation
offrait, sur le plan conceptuel, un nouvel, énorme
avantage. En effet, l'imagerie techniciste des organes
soulève, de manière inévitable, le
problème de la finalité. Comment tous ces
mécanismes d'une haute efficience, d'une parfaite
efficacité, comment toute cette horlogerie
pouvait-elle se construire apparemment sans horloger ?
À partir du moment où l'on pouvait supposer
« codée » dans l'ADN du génome toute
la structure organique, le mystère s'effaçait,
puisqu'il suffisait d'imaginer que l'ADN, promu au rang de
démiurge, dirigeait toute l'épigenèse
de l'être vivant, à la manière d'un
ingénieur dictant ses ordres à ses
subordonnés. Moyennant quoi, on a pu rendre
admissible la finalité, rebaptisée pour la
circonstance téléonomie. Et cela d'autant plus
que la traduction ADN ? Protéines, via l'ARN messager
était un code au sens étroit, technique, du
terme, puisque cette opération associe à tout
triplet de nucléotides un acide aminé bien
défini. Or, dans le premier cas la situation est
toute différente : il s'agit de comprendre comment
« l'information » supposée incluse dans
l'ADN génique, peut spécifier la structure
tridimensionnelle de l'organisation embryonnaire, puis
adulte. (Je parle ici des Pluricellulaires, je reviendrai
sur le cas des Unicellulaires en fin d'exposé.)
Pratiquement toute la théorie biologique moderne
s'est trouvée enfermée dans l'homonymie
abusive de ces deux emplois du mot code, et cet abus de
langage l'a condamnée à une
stérilité conceptuelle dont elle n'est pas
encore sortie...
Comment sortir de cette impasse ? Il ne fait guère de
doute que seule une certaine audace théorique peut
permettre de faire avancer la question. Il faut songer
à réintroduire en Biologie l'imaginaire, cet
imaginaire sans lequel il n'est pas de théorie.
Déjà, dans l'interprétation techniciste
des organes, ne peut-on songer comme autrefois le
suggéra Bergson à renverser l'ordre des termes
? Plutôt que d'expliquer l'organe par l'outil, ne
faudrait-il pas plutôt expliquer celui-ci par
celui-là ? Autrement dit, l'imagination intuitive qui
a permis à l'Homo Faber de construire, bien avant
toute science constituée, des outils doués
souvent d'une remarquable efficience, cette intuition ne
trouvait- elle pas son origine dans un « inconscient
biologique » légué par l'évolution
phylogénétique de l'espèce ? Et ceci
d'autant plus que la plupart des outils ne sont en fait que
le prolongement d'actions, motrices ou physiologiques, et
que l'action dans son organisation spatio-temporelle, nous
est organiquement léguée. Il faut reprendre
ici l'axiome lamarckien : la fonction crée l'organe
non pas au sens banal qui voudrait qu'une structure
organique se crée ou se développe à la
suite de l'usage qui en est fait, mais de manière
plus abstraite, plus « platonicienne ». Toute la
régulation épigénétique et
comportementale d'une espèce repose sur une structure
formelle de caractère géométrique
(topologique) qui se réalise dans l'espace des
activités métaboliques de l'organisme ; une
« fonction » apparaît alors comme le
dispositif régulatoire partiel afférent
à l'homéostasie d'un paramètre
physiologique essentiel (teneur en réserves
énergétiques, en oxygène, en
déchets organiques... par exemple), et la
réalisation organique d'une telle fonction peut
comporter les agents physico-chimiques les plus
variés, comme les organes les plus divers. (Par
exemple, la communication entre animaux d'un même
groupe social peut employer des signaux olfactifs, sonores,
visuels... souvent dans un même but). Toutes ces
« fonctions » concourent ainsi à la
canalisation de la dynamique dans l'espace des
activités métaboliques, formant ainsi un
« attracteur » global (que j'ai proposé
d'appeler la figure de régulation de l'espèce
considérée). »
Je précise que l'article de Thom
représente 25 pages de théorie
détaillée sur cette idée.
|
* un
paradigme est, au sens de Thomas Kuhn (Structure des
révolutions scientifiques, Paris, 1972) une théorie
scientifique qui, à un moment donné, est accepté
par la majeure partie des scientifiques. Le passage d'un paradigme
à l'autre, au cours de l'histoire constitue une
révolution scientifique.
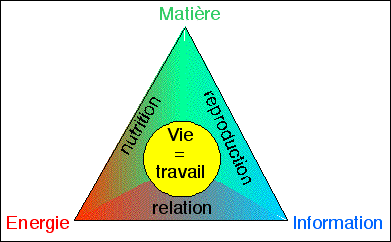{kind=link}
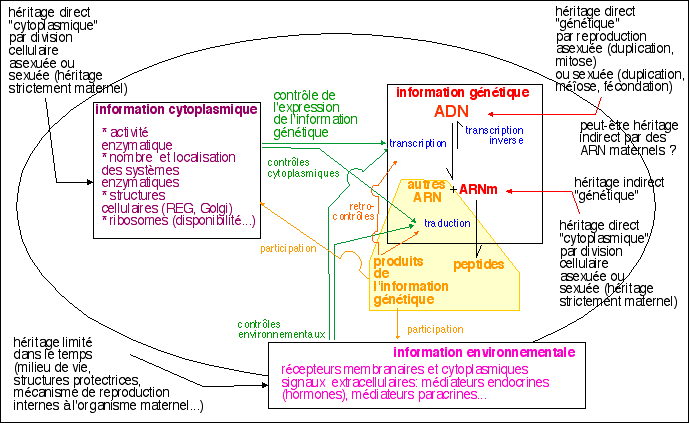{kind=link}