Je peux transmettre la vie...
retour accueil
résumé, formulation
par cycle
Avertissement !
A mon avis le travail pour cette partie est plus difficile pour les
"spécialistes" que pour ceux qui n'ont que des connaissances
anciennes. A l'école , il faut recentrer les concepts sur
l'homme et replacer l'hérédité chromosomique
à une place qui n'est pas privilégiée. Ce
travail est sans aucun doute plus difficile à réaliser
pour un étudiant qui a une licence de biologie des organismes
et qui pourrait (peut-être ?) manquer d'une vision
globale.
|
Le travail de reproduction chez
l'homme est caractérisé biologiquement
par des sexes séparés
(nécessitant la recherche du partenaire),
fécondation interne (organes d'accouplement,
fécondation en milieu liquide) et
viviparité (gestation placentaire) suivie d'un
allaitement (glandes mammaires) et d'un soin aux
jeunes maintenu pendant une assez longue
période.
Du point de vue de la finalité biologique, le
travail de reproduction est don de la vie,
transmission de la vie.
L'acquisition de la capacité à se reproduire
est qualifiée de puberté (du latin
pubes, le poil). Elle correspond à une
maturité structurale et fonctionnelle des organes de
la reproduction (organes génitaux) et s'accompagne de
signes extérieurs (caractères sexuels
secondaires) notamment l'apparition d'une pilosité
(du latin pilus, le poil, le cheveu) aux niveau de
certaines zones de la peau.
|
Cours
L'homme est classé dans l'embranchement des
Vertébrés, classe des Mammifères et ordre des
Primates. La reproduction chez les Primates est une reproduction
sexuée avec sexes séparés
(nécessitant la recherche du partenaire),
fécondation interne (organes d'accouplement) et
viviparité (gestation placentaire) suivie d'un
allaitement (glandes mammaires) et d'un soin aux jeunes
maintenu pendant une assez longue période.
Dans le chapitre précédent nous avons vu les
étapes de la gestation, de l'accouchement, de l'allaitement
et, très rapidement, du soin aux jeunes. Dans cette partie
nous verrons uniquement les différences entre les deux sexes
et la fécondation.
Nous avons présenté la vie comme un travail.
Maintenant nous la voyons sous l'aspect de la reproduction le
travail de reproduction. Ce travail conduit à un
don. Les parents donnent la vie à leur enfant. On parle
aussi de transmission de la vie dans la mesure où les
parents transmettent quelque chose qui ne leur appartient pas. Ces
notions relèvent sans aucun doute plus de l'éthique que
de la biologie et le choix des termes est important. Nous nous
intéressons à une fin (d'où la
nécessité de parler d'éthique): la fin de la
reproduction est bien de donner la vie à des enfants, de les
élever et de perpétuer ainsi l'espèce. Il s'agit
bien d'une fin biologique. L'ensemble des structures, des
organes, des régulations, concourent de façon
coordonnée à une fin biologique : la reproduction.
Donc, il est important de souligner que le travail de reproduction,
met en jeu non seulement la capacité pour la femme à
être fécondée mais aussi à mener à
bien la gestation, l'allaitement puis les soins au(x) jeune(s).... La
fonction de reproduction ne se limite pas à la
fécondation, ni même à la gestation... (si chez
de nombreux mammifères le soin aux jeunes est essentiel
à la fonction de reproduction, combien plus la famille devrait
être protégée chez l'homme...). Notre
étude n'atteindra pas le domaine éthologique alors
qu'il est évidemment essentiel pour ce travail que l'individu
ne réalise pas tout seul...
L'acquisition de la capacité à se reproduire est
qualifiée de puberté (du latin pubes, le
poil). Elle correspond à une maturité structurale et
fonctionnelle des organes de la reproduction (organes
génitaux) et s'accompagne de signes extérieurs
(caractères sexuels secondaires) notamment l'apparition d'une
pilosité (du latin pilus, le poil, le cheveu) aux
niveau de certaines zones de la peau.
Cette partie sera pour nous l'occasion d'aborder les
systèmes de régulations physiologiques que nous ne
pouvons pas détailler dans chacune des parties de notre
étude. La plupart des idées présentées
ici sont reprises du chapitre du cours de
terminale sur la physiologie humaine de la reproduction. Par
contre, il est regrettable que certains manuels, comme
malheureusement le Tavernier (chapitre 1 de la partie 2, p 134),
n'échappent pas au réductionnisme ambiant et
présente la reproduction comme un phénomène
basé avant tout sur la transmission des caractères
héréditaires placés de façon à mon
avis "archaïque" dans un très hypothétique
"programme génétique". Nous reviendrons sur cette
partie en fin de chapitre mais il me semble important de
présenter d'abord la reproduction comme un travail
coordonné de très nombreux organes et plus encore de la
coopération entre deux individus de sexe différents
à une même fin biologique.
I. le travail de reproduction chez
chacun des sexes
a. les déterminismes internes - la capacité interne
des organes au travail de reproduction
1. au niveau de l'organisme entier :
L'apparition de la puberté est sous la dépendance de
facteurs internes mais aussi de l'alimentation. On rapporte par
exemple comme un fait clairement établi que l'âge des
premières menstruations pour les filles européennes est
tombé de 15-16 ans en moyenne en 1900, à 12-13 ans dans
les années 70 ; tout en précisant qu'en 1900 les jeunes
filles de milieux aisés avaient leurs premières
règles à l'âge de 12-13 ans. Une chose est
certaine : l'âge moyen cache une grande variabilité
individuelle.
Les phénomènes de vieillissement touchent aussi bien
les ovaires (épuisement du stock d'ovocytes à partir de
38-40 ans), que l'utérus (ralentissement de la
préparation cyclique à la gestation, augmentation de la
mortalité embryonnaire et des avortements) et les
mécanismes physiologiques régulateurs (cycles ovariens
anormaux.., chute des taux hormonaux à la
ménopause).
Pour les garçons, la puberté est plus tardive.
2. au niveau des organes:
2.1 Chez la jeune femme
Les organes de la reproduction, c'est-à-dire
impliqués de façon directe et nécessaire dans le
travail de reproduction sont:
- les voies génitales : un vagin (impair),
avec les structures annexes correspondant à la partie la
plus externe du tractus génital : la vulve
(fermée partiellement par l'hymen), le
périnée, les grandes et les petites
lèvres, le clitoris): l'organe de
l'accouplement, l'utérus (impair mais terminé
par deux cornes utérines paires) : l'organe de la nidation
et de la gestation, les oviductes ou trompes de
Faloppe (pairs et terminés chacun par un pavillon) :
organes de la fécondation
- les glandes génitales : les ovaires :
organes de la gamétogénèse (production des
gamètes), les mamelles (seins) : organes de la
lactation, des glandes annexes associées aux voies
génitales : les glandes de Bartholin.
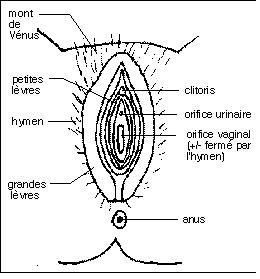
Les organes génitaux externes (vulve) de la femme.
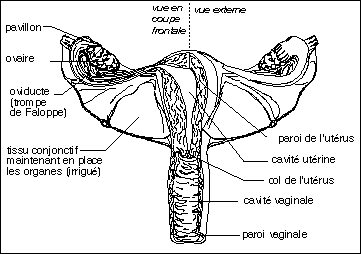
Les voies génitales féminines en coupe frontale (partie
gauche) et en vue externe (partie droite).
Les glandes de Bartholin débouchent à
l'extrémité vaginale, avant le repli des petites
lèvres, non représenté ici.
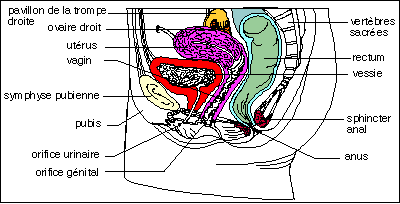
Coupe sagittale du bassin féminin très
schématique. Vous noterez la position de l'utérus, col
orienté vers l'arrière, et des ovaires latéraux,
coiffés par les pavillons dans leur partie
antérieure.
Un peu d'embryologie pour ceux qui veulent
approfondir:
Les ovaires (gonades femelles) se mettent en
place chez le fœtus humain entre la 4ème et la
7ème semaine de vie fœtale (le fœtus mesure entre 6
et 8 mm) à partir des crêtes génitales
situées au sommet des lames latérales qui sont des
massifs de tissus embryonnaires qui font partie du mésoderme
(tissu en position intermédiaire entre le feuillet externe :
l'ectoderme et le feuillet interne : l'endoderme) au stade bourgeon
caudal. Les ébauches gonadiques sont colonisées par
des cellules germinales d'origine
variées selon les classes de Vertébrés. Les
cellules somatiques (par opposition à germinales) forment un
blastème (amas de cellules embryonnaires) d'origine
discutée selon les groupes de vertébrés. Les
premiers follicules ovariens n'apparaissent que vers la 16ème
semaine de gestation (le fœtus mesure alors une dizaine de
centimètres de long). La différenciation du
tractus génital femelle commence vers la
8ème semaine de vie fœtale ( le fœtus mesurant alors
quelques 3-4 cm), à partir des pièces
intermédiaires, issues des mêmes massifs
mésodermiques que ceux donnant les ovaires. Les canaux de
Müller embryonnaires donneront l'oviducte et l'utérus, la
vagin résulte de l'évolution de la zone de jonction
(sinus) entre les canaux de Müller et de Wolff (ces derniers
régressant dans le sexe femelle), les bourrelets
génitaux ectodermiques donneront les lèvres, et le
tubercule génital, qui donne ici le clitoris et le
pénis chez l'homme. En absence de stimulation par les hormones
mâles (testostérone produite par les cellules de Leydig
dès la 8ème semaine de vie fœtale) et hormone
antimüllerienne (AMH), possible jusqu'à la 16ème
semaine, les organes génitaux évoluent naturellement en
organes femelles. Si le rôle d'un facteur contrôlé
par le chromosome Y (TDF, Testis Determining Factor) semble avoir
été mis en évidence, il n'en reste pas moins que
le contrôle de la différenciation sexuelle reste
largement inconnu. On ne connaît pas la liaison
précise entre le sexe chromosomique
(cytogénétique) et le sexe histologique et
physiologique.
|
L'ovaire assure 4 fonctions :
- l'ovogenèse : multiplication des ovogonies
(cellules germinales) et maturation des ovocytes (chez les
Primates les réserves de l'œuf sont faibles mais
la vitellogenèse, accumulation de
réserves de type vitellus, en est une phase
essentielle)
- la folliculogenèse : multiplication des
cellules somatiques entourant l'ovocyte (les cellules
folliculaires participent elles aussi de façon
essentielle à la vitellogenèse, accumulation
des réserves dans l'œuf)
- l'ovulation : libération d'un œuf
(ovocyte mature), on parle aussi de ponte ovulaire et
d'oviposition chez les ovipares
- une fonction endocrine : synthèse d'hormones
stéroïdiennes et peptiques contrôlant le
fonctionnement ovarien mais aussi pratiquement tous les
organes intervenant dans la reproduction (organes
d'accouplement, de ponte, de gestation...).
|
Les ovaires sont irrigués par une artère qui
pénètre entre les follicules et se ramifie. Mais les
capillaires ne dépassent pas la lame basale des follicules. De
nombreuses expériences de greffe ont permis de mettre en
évidence la sécrétion par les follicules en
croissance de nombreux facteurs chimiques (VEGF, TGFalpha...) qui
stimulent la multiplication et la croissance des cellules
endothéliales et donc favorisent la revascularisation des
ovaires greffés.
L'innervation ovarienne (essentiellement sympathique) se
développe en même temps que la vascularisation et les
cordons nerveux suivent les vaisseaux sanguins (dont elles innervent
les fibres lisses) jusqu'aux cellules de la thèque externe des
follicules. De nombreux neuromédiateurs ont été
isolés de l'ovaire (NAdr, dopamine, substance P, VIP, NO...)
et interviennent soit dans l'expulsion des ovocytes, soit dans le
débit sanguin et donc, indirectement, dans la croissance des
follicules.
L'ovulation est un phénomène rapide (quelques
minutes) et libère habituellement un ovocyte secondaire de
façon alternée entre l'ovaire droit et gauche chez la
femme (le taux d'ovulation , nombre d'ovocytes libérés
en même temps est habituellement de 1 chez la vache, 2 chez la
brebis, 8 à 30 chez la truie et 6 à 17 chez la ratte).
La libération simultanée de deux ovocytes par l'un ou
les deux ovaires chez la femme peut conduire à la gestation de
(faux) jumeaux. L'ovulation est "spontanée" chez la femme dans
le sens où elle ne semble pas être habituellement
"provoquée" comme c'est le cas par exemple chez la chatte (la
lapine, la femelle du vison, de l'écureuil...) à la
suite d'une stimulation lors de l'accouplement. Mais il est connu
qu'un stress, un accident, une émotion... peuvent provoquer
l'ovulation chez la femme. Il semble que l'ovaire présente des
contractions musculaires qui assurent l'expulsion de l'ovocyte
secondaire entouré de ses cellules folliculaires formant le
cumulus oophorus et accompagné du liquide folliculaire
(liquor folliculi), lors de la rupture du follicule mûr
(follicule de De Graaf) qui fait saillie à la surface de
l'ovaire (son diamètre atteint 2 cm pour un ovaire d'un
diamètre maximal de 3 cm). La commande de ces contractions
pourrait être d'origine nerveuse mais aussi simplement
causée par la baisse de pression dans la cavité
folliculaire (voir plus bas). L'ovulation s'accompagne parfois de
légers saignements. Après l'ovulation le follicule
rompu se referme et se transforme en corps jaune par une croissance
des vaisseaux sanguins qui colonisent la granulosa dans laquelle les
cellules folliculaires se transforment en cellules lutéales
sécrétrices (luteus = jaune en latin). Un
caillot sanguin persiste au centre du corps jaune. Le corps jaune se
forme en quelques heures, fonctionne une quinzaine de jours puis
régresse rapidement en absence de fécondation (corps
jaune cyclique ou provisoire), il perd sa couleur et se transforme en
masse fibreuse, le corps blanc, qui disparaîtra, ne laissant la
place qu'à une cicatrice à la surface de l'ovaire. En
cas de fécondation le corps jaune se développe et
devient corps jaune gestatif qui persiste pendant presque toute la
durée de la grossesse. Le cycle ovarien est donc long
(5 mois minimum, l'ovulation intervenant au début de la
dernière quinzaine), alterné et chevauchant (entre les
deux ovaires, séparé par 28 jours en moyenne de
décalage, ce qui permet un fonctionnement cyclique avec une
ovulation chez une femme tous les 28 jours). On a l'habitude de
parler d'un cycle sexuel de 28 jours mais c'est celui de
l'utérus qui commande l'événement le plus
marquant : l'apparition des règles qui revient effectivement
tous les 28 jours en moyenne (cette durée varie selon les
femmes et au cours de leur vie ; on a observé que les cycles
les plus longs correspondent aux femmes les plus jeunes (entre 25 et
43 jours entre 13 et 17 ans) et les cycles les plus courts aux femmes
les plus âgées (entre 24 et 32 jours à partir de
40 ans). Ainsi la phase préovulatoire est appelée phase
folliculaire et dure pas en fait 14 jours mais plus de 4 mois et demi
pendant laquelle de nombreux follicules commencent une maturation.
Par contre la phase post-ovulatoire ou lutéale ou encore
lutéinique, dure effectivement environ 13-14 jours si l'on
considère uniquement la période de fonctionnement du
corps jaune.
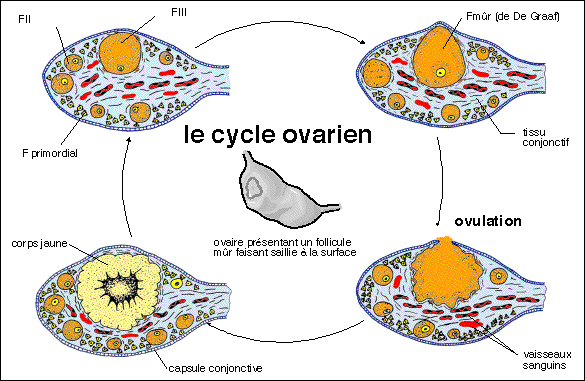
Le cycle ovarien pour un ovaire chevauche celui d'un autre ovaire et
correspond non pas à l'évolution d'un seul follicule
comme on peut le supposer à partir de ce schéma mais de
plusieurs centaines de follicules primordiaux (600 ?) qui donneront
au plus 1 unique follicule mûr ovulant (sauf dans certains cas
d'obtention de jumeaux) puis un corps jaune qui finit par devenir
follicule blanc cicatriciel.
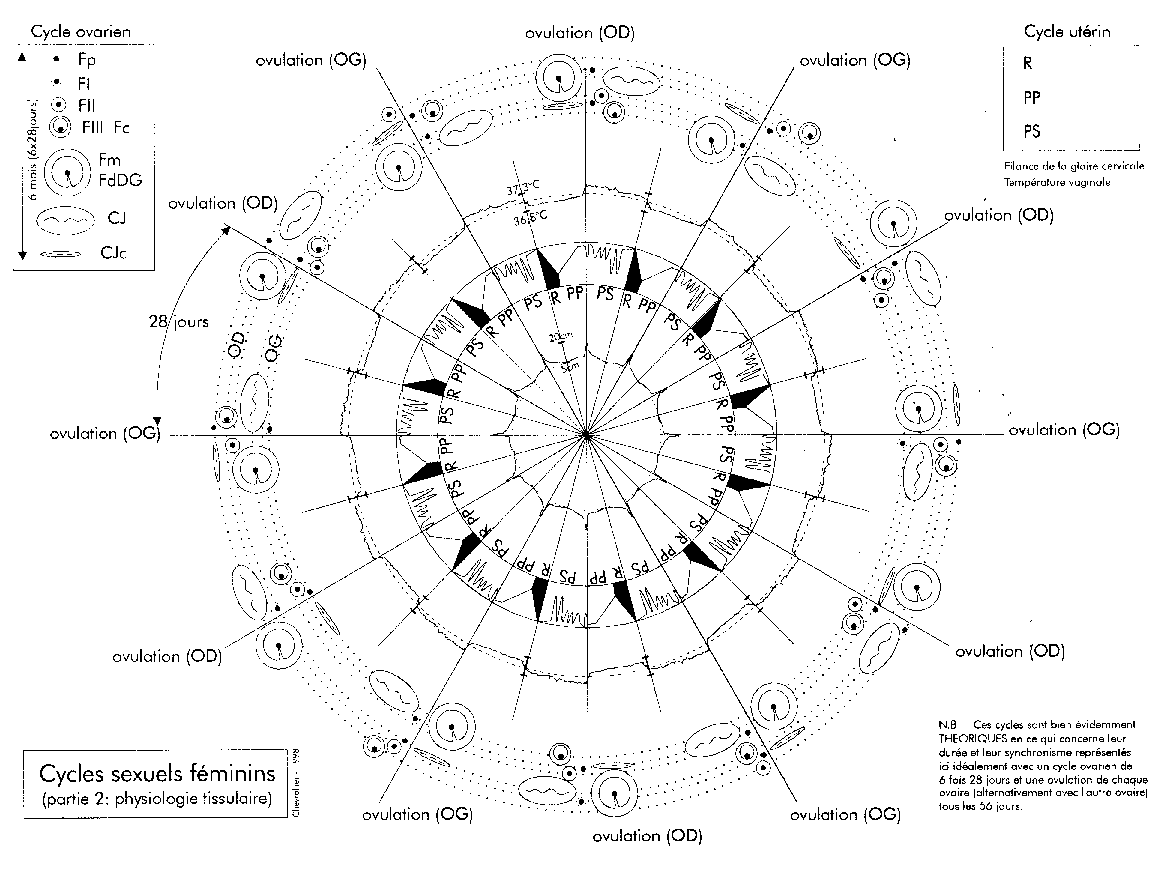
Une représentation des cycles des deux ovaires (droit OD et
gauche OG).
Pour le cycle ovarien : Fp = follicule primordial, FI = follicule
primaire, FII = follicule secondaire, FIII = follicule tertiaire (ou
Fc = follicule cavitaire), Fm = FdFG = follicule mûr ou de De
Graaf, CJ = corps jaune, CJc = corps blanc ou corps jaune
cicatriciel.
Pour le cycle utérin : R = règles, PP = phase
proliférative de la muqueuse utérine, PS = phase
sécrétoire de la muqueuse utérine. Ces cycles
théoriques sont présentées pour un
fonctionnement exactement alternatif des deux ovaires et des cycles
utérins réguliers de 28 jours exactement, ainsi que
pour un temps de passage Fp à Fm (ovulation) de 5 fois 28
jours.
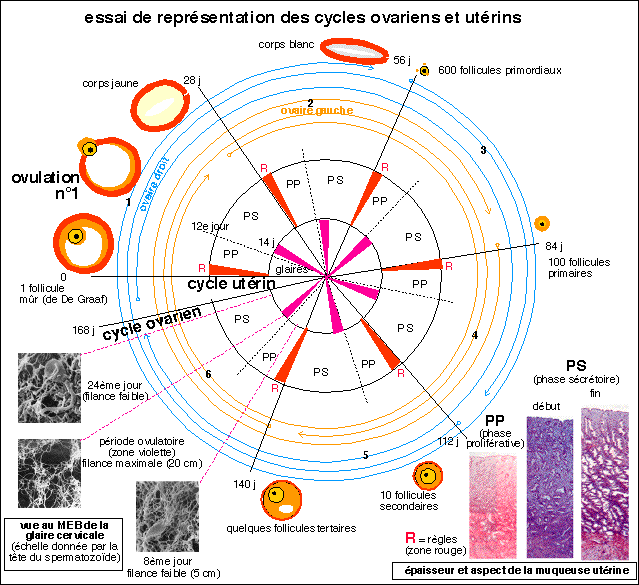
5 mois et demi (168 jours) = UN "cycle" ovarien complet
ou SIX "cycles" ovariens entrecroisés entre l'ovaire droit et
l'ovaire gauche
pour SIX cycles utérins complets
Un schéma simplifié qui rompt avec la
représentation habituelle source d'erreurs.... (photos
d'après Bordas, Tavernier, 1ère AB, 1988)... mais qui
est incontestablement encore trop compliqué....Le cycle
ovarien dure près de 6 mois pour un groupe de follicules
primordiaux mais plusieurs cycles se déroulent en même
temps dans chaque ovaire (décalés de 56 jours) et entre
les deux ovaires (décalés de 28 jours)...alors que le
cycle utérin est bien de 28 jours, de façon
théorique.
On fait habituellement commencer le cycle utérin
à l'apparition des règles ou menstruations ou
encore phase catamériale (chez la femme et seulement quelques
espèces de singes) qui correspond à la
dégradation de la muqueuse utérine
(endomètre) du cycle précédent
(4/5ème de la superficie environ) du notamment à la
nécrose des artères spiralées qui se
développent surtout en fin de phase sécrétoire.
Le sang est rendu incoagulable par sécrétion d'un
facteur fibrinolytique (la fibrine est une protéine
essentielle de la coagulation) local. L'épaisseur de
l'endomètre est alors minimale (environ 1 mm). Les saignements
des règles s'accompagnent souvent de contractions de la
musculeuse utérine (myomètre) plus ou moins
douloureuses. Les règles peuvent durer environ 5 jours (3
à 6 jours) puisque la destruction de l'endomètre
n'intervient pas d'un seul coup mais par lambeaux ; puis
l'endomètre commence à s'épaissir par
prolifération des cellules muqueuses (phase
proliférative), pour atteindre 5 mm d'épaisseur
maximale à ce stade. Pendant toute la durée de la phase
proliférative des contractions utérines plus ou moins
fortes interviennent. A partir du 13ème jour du cycle sexuel,
se fait un "silence utérin" caractérisé par
l'arrêt des contractions utérines, la muqueuse se creuse
de glandes en doigt de gant qui sécrètent un abondant
mucus (glaires) et du glycogène : c'est le début de la
phase sécrétoire. L'épaisseur maximale de
l'endomètre, atteinte pendant cette phase (phase
lutéinique ovarienne), est de 8 mm chez la femme. On notera
l'aspect "déchiqueté" de l'endomètre (on parle
de dentelle utérine CHEZ LA LAPINE), favorable à la
nidation, c'est-à-dire à l'installation, dans
une de ces cryptes, de l'embryon issu de la fécondation, qui a
lieu au niveau d'une des trompes. Au niveau du col de
l'utérus, la muqueuse est réduite et
sécrète un mucus clair plus ou moins abondant et filant
(glaires cervicales de cervix = le col en latin) qui
permettent de déterminer l'étape du cycle sexuel (on
mesure la filance, c'est à dire l'étirement sans
rupture qui est maximal autour de l'ovulation : plus de 20 cm).
Chez la femme le cycle vaginal reste discret alors qu'il est
accompagné de profondes modifications chez de nombreux
mammifères. Le frottis vaginal chez la femme permet
essentiellement de dépister un éventuel cancer de
l'utérus plutôt que de déterminer les
étapes du cycle sexuel, même si la période
post-ovulatoire est marquée par une augmentation de la
kératinisation des cellules et donc du nombre de cellules
mortes (mesuré par l'index caryopycnotique).
2.2 Chez le jeune homme
Le système reproducteur masculin comprend :
- des voies génitales, communes
(urètre) dans la partie terminale pénienne
(externe) avec les voies urinaires, au sein d'un organe copulateur
impair, le pénis. Les voies génitales
internes se séparent d'avec les voies urinaires au niveau
de la prostate et se continuent par deux spermiductes (canaux
déférents) aboutissant à
l'épididyme coiffant chaque testicule.
L'épididyme est le lieu d'acquisition de la
fécondance des spermatozoïdes et de leur
aptitude à la mobilité (ils ne seront mobiles
qu'après l'éjaculation). Le temps de transit dans
l'épididyme est de l'ordre de 10 jours.
Les canaux déférents sont essentiellement un
réservoir (d'une contenance supérieure à
plusieurs éjaculats) mais ils peuvent éliminer les
spermatozoïdes non utilisés par phagocytose ou par
élimination dans les urines. Les spermatozoïdes y
conservent leur pouvoir fécondant pendant au moins trois
semaines. Les canaux déférents sont innervés
au niveau de leurs muscles lisses.
- les glandes de le gamétogenèse sont les
testicules (gonades masculines pairs) situés
à l'extérieur de l'organisme dans les bourses. Les
glandes annexes sécrétant la majeure partie du
liquide séminal (sperme) sont la prostate (deux
demi-glandes regroupées en un organe impair), les
vésicules séminales, les glandes
bulbo-urétrales, et éventuellement
l'épididyme qui a à la fois un rôle de
stockage et de maturation des spermatozoïdes.
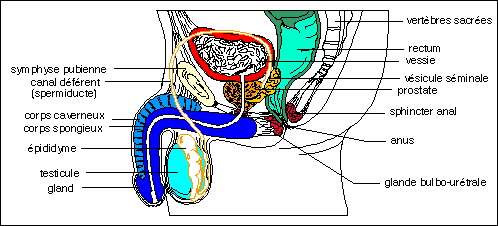
Coupe sagittale de l'appareil génital masculin.
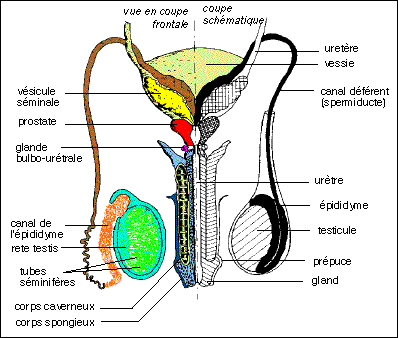
Un schéma inhabituel de l'appareil génital masculin en
vue frontale
(d'après Vander in Précis de Physiologie, Doin, 1998)
mettant bien en évidence les organes ou conduits pairs ou
impairs.
Chez l'homme comme chez de nombreux mammifères, les
testicules migrent avant la naissance, depuis une position
embryonnaire voisine du rein, vers une position à
l'extérieur de l'abdomen, dans des poches cutanées (les
sacs scrotaux) qui, avec les fibres élastiques et musculaires
lisses du derme (muscle dartos) et les muscles lisses
étirés provenant de la cavité abdominale (muscle
crémaster du grec kremastêr = suspenseur),
forment les bourses. Le maintien du testicule et de
l'épididyme à une température inférieure
de 4 à 6°C à celle du corps est indispensable chez
l'homme pour le bon déroulement de la méiose et la
conservation des spermatozoïdes. La cryptorchidie (maintien des
testicules en position haute, intra-abdominale) entraîne une
stérilité définitive et doit donc être
opérée rapidement chez le jeune enfant. Le maintien de
la température basse au niveau du testicule résulte de
l'arrangement vasculaire (on parle d'échangeur thermique) au
niveau du testicule (la contraction du dartos réduit la
surface du testicule), de la position du testicule par rapport
à l'abdomen (la contraction des cremaster vient placer les
testicules contre la paroi abdominale), et d'une régulation
circulatoire (si la température scrotale est
élevée artificiellement le rythme respiratoire
s'accélère).
|
Les testicules présentent principalement
deux fonctions :
* la spermatogenèse : production de
spermatozoïdes par multiplication des cellules
souches, méiose et
spermiogenèse (transformation des spermatides
en spermatozoïdes)
* une fonction endocrine : sécrétion
d'androgènes (stéroïdes) et de
peptides indispensables au développement et à
l'activité des voies génitales et des glandes
annexes masculines, sous la dépendance de
médiateurs endocrine dits gonadotropes.
|
3. au niveau des cellules
3.1 Chez la jeune femme
Du point de vue des cellules sexuelles, les cellules primordiales
germinales d'origine endodermique, appelées gonocytes
se divisent activement par mitose entre le 3ème et le
5ème mois après la fécondation et donnent
naissance aux ovogonies (groupées par paquets et
reliées par des ponts cytoplasmiques). Commence alors une
phase d'accroissement pendant laquelle les ovogonies se
séparent, s'entourent de quelques cellules folliculaires
aplaties, se chargent de réserves, augmentent de
diamètre jusqu'à atteindre environ 50 µm, et
présentent un certain nombre de modifications au niveau du
noyau car les chromosomes deviennent visibles (condensation de
l'ADN). Certaines ovogonies sont clairement au stade leptotène
de la prophase méiotique, on dit qu'ils sont
entrés en méiose. On observe de tels stades chez la
femme à partir de la 7ème semaine de vie fœtale
jusqu'à la naissance, soit environ 20 jours après le
début de différenciation sexuelle de l'ovaire en gonade
féminine. Puis les chromosomes (qui sont arrivés au
stade diplotène) se décondensent. On note que cette
maturation se fait toujours avec un pic d'oestradiol chez tous les
Mammifères étudiés, mais aussi chez des Reptiles
et des Oiseaux. Ces gonocytes sont alors devenus des ovocytes
I (il faut veiller à conserver le terme d'ovocyte
primaire, souvent masqué par le chiffre I que l'on
énonce "un" parfois un peu rapidement; le terme d'ovocyte
secondaire faisant référence à un ovocyte
qui est issu d'une division de l'ovocyte primaire et qui est
présent chez l'homme mais pas chez tous les animaux, bien
évidemment) dont le stock est donc déterminé
à la naissance. On estime à près de 7 millions
le nombre de gonocytes qui donneront environ 1 à 2 millions
d'ovocytes I à la naissance, et on pense qu'il ne reste plus
qu'environ 300.000 ovocytes I dans les deux ovaires à
l'âge de 7 ans (au moment de la puberté), tous les
autres ovocytes ayant dégénéré. On pense
qu'après les quelques 300 ovulations d'une femme au cours de
sa vie sexuelle (entre la puberté et la ménopause), il
y a épuisement du stock d'ovocytes, ce qui fait quelques 600
à 1000 ovocytes I au total impliqués dans chaque
phénomène de maturation conduisant à
l'ovulation. On considère que l'arrêt du fonctionnement
ovarien coïncide avec l'épuisement du stock d'ovocytes
(la femme a un épuisement très précoce par
rapport à d'autres Mammifères pour lesquels celui-ci
arrive en fin de vie). Du point de vue du stade méiotique, on
dit souvent que l'ovocyte I semble a commencé la
première division de méiose, mais en fait l'ovocyte
I n'est pas bloqué en prophase de 1ère division mais
à un stade de décondensation qui fait suite à
une maturation sexuelle et qui est similaire au stade G2
(interphase, après duplication de l'ADN) mais avec un
début de condensation de l'ADN (source : Biologie
moléculaire de la cellule, 3ème éd., p 1020).
Dans ce cas la méiose de commencerait que quelques jours ou
quelques dizaines d'heures avant l'ovulation (actuellement on parle
de déblocage de la méiose ou reprise de la
méiose, 12 h avant l'ovulation...). Les
phénomènes cellulaires qui interviennent lors des
étapes qui se déroulent pendant la longue phase de
repos et de dégénérescence (on parle
d'atrésie (du grec a : privatif et
trêsis : perforation) folliculaire : disparition des
cellules folliculaires et de l'ovocyte sans ovulation) ont des
déterminismes encore mal connus (1 ovocyte I sur 20 en
"réchappe"... et 1 ovocyte présent à la
naissance sera ovulé sur 7.000 ). Les mécanismes
cellulaires de l'atrésie restent très discutés.
Ils sont rapprochés de l'apoptose (du grec apo : loin
et piptein : tomber), mécanisme de dégradation
enzymatique de l'ADN, car des cellules de follicules
atrésiques présentent une transformation du noyau en
une masse compacte de chromatine (cellule dite pycnotique, du grec
puknos : compact). De nombreux facteurs chimiques ont
été invoqués ainsi que la vascularisation du
follicule.
Un prolongement : ne pas oublier que le concept de
méïose n'est .....qu'un concept
:
Si l'on part de considérations cytologiques et
si l'on se réfère aux mécanisme connus sans
s'intéresser à la structure des chromosomes, on peut
proposer une interprétation des mécanismes
reproducteurs avec un seul type de division cellulaire: la
mitose.
Je m'explique:
Que se passe-t-il donc de différent dans les cellules
sexuelles ? D'une part il y a une période de maturation
sexuelle qui est plus ou moins longue mais qui
précède la "mitose sexuelle". Classiquement on place
ces phases dans la prophase de la première division de
méiose. On distingue 5 stades inégaux ou non selon
l'aspect des chromosomes qui sont alors visibles (leptotène,
zygotène, pachytène, diplotène et
diacinèse). Cette période de maturation est toujours
longue par rapport aux divisions qui lui font suite (plus de 11 jours
sur 12 en comptant les deux divisions aboutissant à la
formation des spermatozoïdes chez la souris, 6 jours sur 7 pour
la formation des grains de pollen chez le lis, plus de 10 ans si l'on
considère l'ovocyte féminin, et plus de 20 jours sur 21
pour les spermatozoïdes humains).
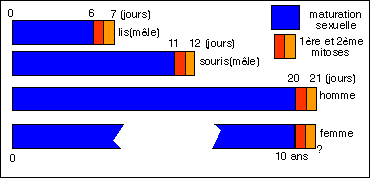
Comparaison entre les temps de maturation de la cellule sexuelle et
la durée des deux divisions (appelées mitoses ici)
qui lui succèdent chez quelques organismes.
C'est pendant cette longue période que l'on
place l'hypothétique crossing-over (voir à ce sujet
le cours de
terminale). Si l'on se
réfère à ce qui se passe lors de la division qui
suit cette période de maturation sexuelle, il y a toujours
séparation des chromosomes homologues et non des
chromatides... pourquoi ne pas alors invoquer les fameux complexes
synaptonémaux et les chiasma qui en résultent (?) pour
justifier que dans la mitose qui lui fait suite, les chromatides
de chaque chromosome restent aisément accolées alors
que les chromosomes homologues sont tirés chacun d'un
côté de la cellule ou de l'autre. La mitose qui fait
suite à une maturation sexuelle, que l'on peut qualifier de
mitose sexuelle sépare donc les homologues et non les
chromatides de chaque chromosome. Si les cellules filles en restent
là et ne se divisent plus on a des cellules à n
chromosomes mais avec une quantité d'ADN identique à
celle de la cellule de départ (classiquement notée Q).
Un argument important dans cette interprétation est que lors
de cette division il n'y a pas de division des centrioles, comme pour
toutes les mitoses des plantes. Ce phénomène est aussi
retrouvé dans les activations
parthénogénétiques des ovocytes de
mammifères qui se divisent sans centriole, même si les
centrioles semblent se former de novo, après plusieurs
divisions de segmentation (source: La reproduction des
vertébrés, Masson, 1998, p 190).
La division suivante peut donc se faire directement sans passer par
une interphase avec notamment une phase S de synthèse. Une des
clés de la division cellulaire est sans aucun doute dans la
formation et la duplication des centrosomes (voir paragraphe
précédent). On sait provoquer chez l'oursin des
bourgeonnements de la cellule qui contiennent des centrosomes mais
pas de noyau (Biologie moléculaire de la cellule, 3ème
éd., p 914). La mitose intervenant à la suite de la
mitose sexuelle et non précédée par une
interphase pourrait donc séparer les chromatides de chaque
chromosomes pour obtenir ainsi des cellules haploïdes avec une
quantité d'ADN de Q/2.
Cette interprétation de la maturation sexuelle des cellules
dites "germinales" et des éventuelles mitoses qui suivent
remet aussi en cause la notion de lignée germinale et
somatique qu'il serait tout aussi intéressant de bousculer
un peu.
Lors de la folliculogénèse, on pourra noter
que les cellules de la granulosa forment une population
homogène, reliées par des jonctions perméables
(gap-junctions). La granulosa est séparée de la
thèque interne par une lame basale (composée
essentiellement de fibronectine) que ne pénètrent ni
les capillaires sanguins, ni les fibres nerveuses.
(Cette disposition est à rapprocher de la
structure du testicule avec les cellules de Sertoli et germinales
d'un côté et les cellules de Leydig de l'autre de la
lame basale..). A maturité, un follicule féminin
mesure 15 à 20 mm et contient près de 5 millions de
cellules folliculaires.
On en arrive en effet au problème central en biologie de la
formation de l'œuf (car c'est bien lui qui va déterminer
le développement embryonnaire).
Durant l'ovogenèse, la phase de maturation de l'œuf
qui comprend la maturation sexuelle au sens employé ci-dessus,
ne présuppose pas des éventuelles divisions qui lui
font suite. On observe en effet que l'œuf (mature donc
libéré par la gonade) peut être, selon les
groupes animaux, à différents stades de division et
donc, vis-à-vis de son matériel
génétique, peut être diploïde (la
quantité d'ADN est de 2Q par rapport à la
quantité d'ADN d'une ovogonie et le nombre de chromosomes est
de 2n (l'étoile de mer ou l'Ascaris par exemple), l'œuf
peut aussi être bloqué en métaphase chromosomique
d'une première division (Insectes ou Prochordés)) ou
haploïde (à la suite d'une déroulement complet
d'une division, la plupart du temps très inégale
donnant un globule polaire et un ovocyte secondaire qui
possède n chromosomes avec une quantité d'ADN Q) avec
début d'une nouvelle division (et blocage en métaphase
"chromosomique" comme les Vertébrés) ou
déroulement complet comme l'oursin (n chromosomes avec Q/2
d'ADN).
Le deuxième point important concerne les
caractéristiques cytologiques de la cellule œuf
(désormais au sens de cellule sexuelle femelle mature). Sa
taille (en fait son volume)
tout d'abord et ses réserves qui sont
liées à ce paramètre. Un cellule embryonnaire
typique humaine a un diamètre d'une dizaine de
micromètres alors que la cellule œuf en mesure
typiquement 120 (le rapport entre le volume de ces deux cellules
supposées sphériques est de 1/1000 car je n'ose
rappeler que le volume augmente avec le cube du rayon de la
sphère). On a un rapport assez équivalent pour
l'œuf d'oursin (Echinodermes) ou de Cnidaire. Une cellule
embryonnaire d'amphibien ne semble pas être fondamentalement de
taille différente à celle d'un embryon humain et donc
voisine d'une à quelques dizaines de micromètres alors
que la cellule œuf d'amphibien atteint de un à quelques
millimètres (ce qui fait un rapport de volume de l'ordre de
1/8000). On a un rapport voisin pour les Annélides, les
Mollusques Lamellibranches, les Agnathes.... Les œufs de
très grande taille (oiseaux, reptiles, Céphalopodes,
Sélaciens, Monotrèmes...) atteignent quelques
centimètres pour une cellule embryonnaire de quelques
centaines de micromètres tout au plus, ce qui fait un rapport
volumique de plus de 1/ 1.000.000 (quoique l'on s'éloigne de
plus en plus du modèle sphérique). Pour les arthropodes
(dont les insectes), l'œuf, franchement allongé et
atteignant quelques millimètres, semble présenter un
rapport voisin de celui des amphibiens. Cet accroissement volumique
peut être relié directement à la quantité
de réserves. Les catégories cités plus haut
correspondent aux classiques œufs alécithes,
oligolécithes, hétérolécithes,
télolécithes et centrolécithes qui font aussi
référence à la position du
noyau de l'œuf dans la cellule géante. Ces
catégories, très perfectibles, sont essentielles pour
comprendre les premières étapes du développement
embryonnaire. On citera par exemple les cocons des Plathelminthes qui
contiennent une œuf entouré de cellules vitellines
complètes... et qui ne rentrent pas dans ce cadre trop
étroit.
On en arrive donc aux phénomènes
chromosomiques qui se déroulent lors de
l'ovogenèse. Si l'on considère l'augmentation de volume
de la cellule, il semble indubitable qu'il doit y avoir une
expression extrêmement performante de l'information
génétique (partiellement bien sûr) au cours de
cette période, ne serait-ce que pour synthétiser les
innombrables réserves. On doit bien sûr prendre en
compte l'apport extracellulaire (la synthèse du vitellus est
classiquement localisée au foie chez les oiseaux par exemple,
puis celui-ci passe dans le sang pour gagner les ovaires). Il ne faut
pas non plus négliger le rôle des cellules folliculaires
ou d'autres cellules qui peuvent avoir un rôle nourricier pour
l'œuf en cours de croissance. Mais on explique aisément
par cet énorme besoin de synthèse les fréquentes
polyploïdies observées. Il est aussi
connu (Biologie moléculaire de la cellule, 3ème
éd., p 1024) que le nombres des gènes codant pour des
ARN ribosomiaux augmente fortement (jusqu'à plusieurs millions
de copies de ces gènes pour les amphibiens par exemple alors
qu'une cellule embryonnaire typique n'en contient que quelques
centaines). On notera aussi l'interprétation que l'on fait des
chromosomes en écouvillon de nombreux ovocytes en cours
de maturation, les boucles d'ADN déroulé correspondant
peut-être à des sites en cours de transcription.
En résumé, chez la femme le gamète femelle
ou cellule œuf, est un ovocyte secondaire (ovocyte II)
(bloqué en métaphase de deuxième division de
méiose, accolé à un globule polaire (GP1),
entouré d'une membrane pellucide et de nombreuses cellules
folliculaires (formant le cumulus oophorus).
3.2 Chez le jeune homme
Les testicules humains peuvent être décrits comme
comprenant deux compartiments séparés par une membrane
basale (voir schéma dans la partie suivante) et
entourés par une tunique fibreuse (l'albuginée)
formée de cellules conjonctives et de cellules musculaires
lisses, l'ensemble étant richement irrigué:
- un compartiment interne ou germinal : avec deux types
de cellules : les cellules de Sertoli (cellules
somatiques, c'est-à-dire non transmises lors de la
reproduction : elles appartiennent au soma) et les
cellules germinales (on dit aussi cellules de la
lignée germinale : spermatogonies, spermatocytes,
spermatides et spermatozoïdes) qui subissent notamment la
méiose mais aussi de nombreuses mitoses et qui sont
entourées par les cellules de Sertoli.
A partir de la puberté les cellules de Sertoli ne se
multiplient plus, elles se fixent à la membrane basale et
établissent des liaisons entre elles en formant une
barrière (barrière hémato-testiculaire)
imperméable aux hormones peptidiques, aux
lipoprotéines et aux grosses molécules en
général.
Les cellules germinales primordiales se multiplient dans les
crêtes génitales chez l'embryon (voir embryologie
plus haut) puis dans le compartiment germinal entre les cellules
de Sertoli jusqu'à la puberté en formant des
spermatogonies souches qui se diviseront
régulièrement à partir de la puberté
pour donner naissance aux lignées
spermatogénétiques successives. Contrairement
à l'ovaire il n'y a pas de début de méiose
avant la puberté (cette quiescence semble du à un
facteur inhibiteur sécrété par les cellules
de Sertoli MPF(meiotic preventing factor) : en effet des
spermatogonies souche déplacées hors du
contrôle des cellules de Sertoli, entrent en méiose).
Les spermatogonies qui se divisent restent souvent reliées
par des ponts cytoplasmiques et forment ainsi des clones dont les
mitoses sont synchronisées. Comme pour l'ovaire on observe
(sans explication) un grand nombre de
dégénérescences par apoptose des
spermatogonies souches. Le terme cycle
spermatogénétique désignant les
transformations d'une spermatogonie à un ensemble de
spermatozoïdes (multiplication, méiose,
spermiogenèse), est certes mal choisi car il n'y a pas de
retour au point de départ (le terme d'hélice
spermatogenétique serait bien plus approprié :
ce sont en effet des hélices où les cellules de la
lignée germinale présentent une série de
transformations successives qui s'imbriquent le long du tube
séminifère) mais est employé. Le nombre
théorique de spermatozoïdes obtenus par division
mitotiques puis méiose d'une spermatogonie souche (souvent
3 mitoses goniales suivies d'une méiose et donc
(1x2=>2x2=>4x2=>8x4=>) 32 spermatozoïdes) est...
théorique. Le nombre de divisions goniales est
régulé par d'innombrables facteurs endocrines et
paracrines (des cellules de Sertoli notamment amis aussi des
cellules germinales en cours de spermatogenèse qui exercent
un rétrocontrôle sur les mitoses goniales...). De
nombreuses dégénérescences par apoptose
interviennent aussi. La phase de spermiogenèse,
élaboration d'une vésicule contenant de nombreuses
enzymes (acrosome) et perte de la majeure partie du cytoplasme
dans lequel il reste un appareil mitochondrial important
placé en hélice autour du centriole essentiel pour
la fécondation et prolongé par le flagelle,
structure locomotrice, compaction de l'ADN avec élongation
et compaction nucléaire, production et mise en place de
protéines membranaires et cytoplasmiques nécessaires
à la fécondation (notamment d'un facteur
d'activation de l'ovocyte). Ces modifications nécessitent
la transcription et la traduction de l'information
génétique. Ces mécanismes sont relativement
synchronisés entre les spermatides issus d'une même
gonie, du fait des ponts cytoplasmiques et de la continuité
de la membrane cellulaire du moins dans les premières
étapes de la spermiogenèse. La durée de
l'hélice spermatogénétique est d'environ
74 jours pour l'homme (ce temps correspond
approximativement aux 24h de durée d'une mitose goniale
suivie de la durée mesurée très
précisément à l'aide de marqueurs radioactifs
et strictement constante pour chaque espèce entre le
début de formation des spermatocytes I (stade
leptotène) et l'obtention des spermatozoïdes, la
méiose durant elle aussi environ 24h).
Des cellules myoïdes (musculaires lisses mais peu
différenciées) doublent la lame basale sur sa partie
externe. Elles ne semblent pas innervées et se contractent
sous l'action de médiateurs circulants. Elles participent
à l'expulsion mécanique des spermatozoïdes du
testicule.
- un compartiment externe ou interstitiel composé
uniquement de cellules somatiques, cellules conjonctives,
cellules des systèmes sanguins et lymphatiques et
cellules à fonction endocrine (cellules de Leydig ou
encore appelées de façon trop imprécise les
cellules interstitielles). Ce compartiment est
innervé. Les cellules de Leydig proviennent du
même amas de cellules embryonnaires que les cellules de
Sertoli. Elles ne se multiplient que pendant la période
embryonnaire et commencent immédiatement à
sécréter des stéroïdes intervenant dans
la différenciation sexuelle des voies et des organes
génitaux masculins. Puis elles régressent
jusqu'à la puberté à partir de laquelle elle
présentent chez l'homme une activité endocrine
cyclique quasi continue jusqu'à la mort.
4. au niveau des molécules : le
complexe hypophyse-gonades
Chez la jeune femme
Il semblerait que chez la jeune fille impubère, les
concentrations en hormones hypophysaires de type gonadostimulines (LH
et FSH) soient très faibles et constantes, les concentrations
en hormones ovariennes (oestradiol, oestrone...) extrêmement
faibles, tout comme la sécrétions endocrines
hypothalamiques de gonadolibérines (GnRH). La mise en place de
la fonction endocrine sexuelle hypothalamique (et donc du
système nerveux) semble être le facteur
déclenchant de la mise en place des cycles sexuels
féminins.
Le fonctionnement cyclique de l'ovaire est la première
donnée. Conjointement aux modifications histologiques et
cytologiques que nous avons vu précédemment, la
fonction endocrine de l'ovaire est elle aussi cyclique. La phase
folliculaire s'accompagne d'une sécrétion croissante
d'œstrogènes (par les cellules de la thèque
interne et les cellules folliculaires) et la phase lutéinique
d'une sécrétion de progestérone (par les
cellules lutéales), alors que la sécrétion
d'œstrogènes est maintenue de façon plus ou moins
importante. En cas de fécondation, le développement
très important du corps jaune gestatif est à l'origine
d'une sécrétion de progestérone beaucoup plus
importante. Les sécrétions sont ensuite relayées
par le placenta, une fois celui-ci en place.
Le contrôle du fonctionnement cyclique est
réalisé par l'antehypophyse. Si le rôle de la
FSH, hormone antéhypophysaire, semble dominer pendant
la phase folliculaire, elle est aussi nécessaire en phase
lutéinique. De même la LH, présente aussi
en phase folliculaire, semble jouer un rôle fondamental dans
l'ovulation (pic ovulant) et dans la lutéinisation (formation
du corps jaune et sécrétion de progestérone par
celui-ci). Une liaison directe nerveuse pour l'ovulation existe aussi
certainement. Etant donné les faibles taux sanguins de ces
hormones hypophysaires on est loin de pouvoir suivre aisément
les variations de leur sécrétion en fonction de tous
les paramètres supposés efficaces. On pense
actuellement que ces deux hormones sont
sécrétées alternativement par les mêmes
cellules...
Chez le jeune homme
A partir d'environ onze ans (puberté), des cellules de Leydig
de nouveau en activité sont décelables dans le tissu
interstitiel. Leur sécrétion d'androgènes
(testostérone) est sous la dépendance des
hormones hypophysaires LH (nommée aussi ICSH
chez l'homme : intersticial cells stimulating hormone) et FSH
, elles-mêmes commandées par l'unique hormone
hypothalamique la GnRH, comme chez la femme. La
sécrétion de GnRH chez l'homme adulte est
rythmique (un pic toutes les 90 minutes). L'administration
continue de GnRH provoque une désensibilisation de ses
récepteurs et on observe alors une castration chimique. La
testostérone agit en retour (feed-back négatif)
sur la sécrétion des hormones hypophysaires,
indirectement en agissant sur l'hypothalamus (la testostérone
diminue la fréquence des pics de GnRH) et directement sur
l'hypophyse (la testostérone inhibe la sécrétion
de LH). La testostérone agit sur les cellules de Sertoli
notamment par l'intermédiaire d'une protéine de
liaison des androgènes elle-même
sécrétée par les cellules de Sertoli sous
l'action de la FSH. Les cellules de Sertoli sécrètent
aussi l'inhibine, une hormone inhibant la
sécrétion de FSH par l'hypophyse. La
testostérone, hormone masculinisante intervient sur
l'ensemble de l'appareil génital et est responsable du
maintien des caractères sexuels secondaires (pilosité,
croissance, répartition des masses graisseuses et musculaires,
timbre de la voix, sécrétions des glandes
sébacées, développement des muscles
squelettiques...).
Des précisions (issus de
Reproduction des vertébrés, Masson, 1998)
un essai de présentation globale:
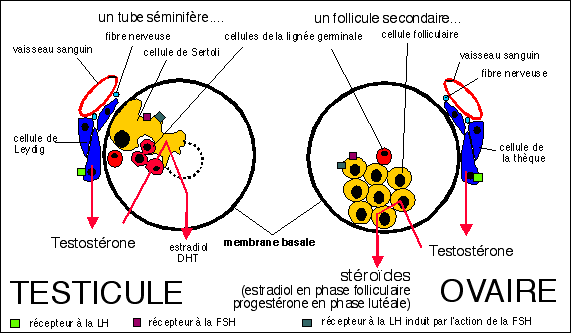
Une comparaison vraiment intéressante...
à gauche un tube séminifère, à
droite un follicule secondaire TRÈS
SCHÉMATISES...
(voir le texte ci-dessous pour des explications plus
détaillées)
Le développement ovarien et testiculaire, du moins à
partir d'un certain stade de croissance, est sous le contrôle
absolu des gonadotropines (FSH et LH).
Les cellules de la thèque interne ont des récepteurs
à la LH (dès la formation de l'antrum) mais pas de
récepteurs à la FSH.
Par contre les cellules de la granulosa expriment très
tôt des récepteurs à la FSH, cette
dernière induisant l'expression de récepteurs à
la LH.
De la même façon, les cellules de Leydig ont des
récepteurs à la LH mais pas les cellules de Sertoli,
uniquement sensibles à la FSH.
L'oestradiol (orthographié aussi
estradiol) est produit essentiellement par les cellules de la
granulosa qui transforment (activité aromatase) la
testostérone en estradiol, sous l'action stimulante de
la FSH. C'est la thèque interne, sous l'action de la LH, qui
sécrète de la
testostérone.
La FSH intervient aussi en stimulant la
sécrétion d'inhibine par les cellules de la granulosa ;
l'inhibine, à son tour stimule la sécrétion de
testostérone par les cellules de la thèque).
Chez l'homme, ce sont aussi les cellules de Sertoli qui
sécrètent l'inhibine sous l'action de la
testostérone (potentialisée par les protéines
liant les androgènes sécrétées par les
cellules de Sertoli sous l'action de la FSH).
Il y a donc clairement une analogie entre
- les cellules externes: cellules Leydig (sensibles à la
LH) sécrétant la testostérone et cellules des
thèques internes (sensibles à la LH)
sécrétant aussi la testostérone
- les cellules internes : cellules de Sertoli (sensibles
à la FSH) sécrétant l'inhibine et
contrôlant la spermatogenèse essentiellement sous
l'action de la testostérone, et les cellules de la
granulosa (sensibles d'abord à la FSH) contrôlant
l'ovogenèse et sécrétant des
stéroïdes en transformant la testostérone.
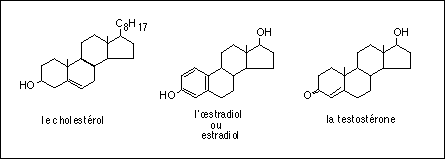
Remarque : les hormones stéroïdes sont
dérivées du cholestérol et sont
synthétisées par les cellules qualifiées de
stéroïdogènes notamment grâce à leur
équipement en récepteur au cholestérol circulant
(sous forme de lipoprotéines) et d'enzymes assurant la
synthèse mitochondriale des stéroïdes (les
chaînes de synthèse fonctionnent notamment grâce
à des cytochromes spécifiques). Les gonadotropines
stimulent à la fois la synthèse des
stéroïdes et des cytochromes. Progestérone,
testostérone et estradiol ont des récepteurs
nucléaires spécifiques (activant directement certains
gènes...) dans de nombreux neurones, cellules hypophysaires,
des voies génitales et du foie. Mais les stéroïdes
agissent aussi à des niveaux non génomiques (estradiol
par exemple sur les récepteurs du GABA ou les
récepteurs des opiacés...).D'une façon
très générale les stéroïdes agissent
sur la multiplication des cellules, leur différenciation
fonctionnelle et leurs activités de synthèse.
Sous l'action des gonadotropines (LH et FSH) les follicules
sécrètent de nombreuses protéines dont certaines
ont des actions autocrines (stimulation de la cellule
sécrétrice elle-même), d'autres paracrines
(stimulation des cellules voisines sans transport par le sang),
d'autres enfin endocrines (sécrétion
d'hormones), notamment ceux agissant sur la sécrétion
des gonadotropines par l'antehypophyse. On peut citer les facteurs de
croissance de la famille des TGF-ß (activines-inhibines,
follistatine), les facteurs de la famille des IGFs
(Insuline-like-Growth Factor), les facteurs de la famille des EGFs
(Epidermal Growth Factor).
Le blocage de la méiose persiste tant que l'ovocyte
n'a pas accumulé de réserves (80% de sa taille finale
chez les Mammifères). Alors, si l'ovocyte est cultivé
isolément, il reprend spontanément sa méiose. On
a donc recherche un facteur d'inhibition, qui reste inconnu à
ce jour. Certains auteurs pensent donc qu'il n'existe pas. La
séparation de l'ovocyte d'avec ses cellules folliculaires (qui
sont très étroitement liées à l'ovocyte
car des "pieds " pénètrent la partie
périphérique du cytoplasme, isolée bien
sûr par la membrane plasmique), fait intervenir dans tous les
cas de profonds remaniements cytologiques. La reprise de
méiose est toujours marquée par un pic de Ca2+,
comme la fécondation.
On a mis aussi en évidence un facteur de nature chimique
encore inconnu qui semble être indispensable à la
décondensation du noyau du spermatozoïde qui a
fécondé l'ovocyte. Ce facteur (MPGF = male pronucleus
growth factor) est sécrété par les cellules de
la granulosa, est diffusible et est accumulé dans le
cytoplasme de l'ovocyte....
L'ovulation semble être d'abord sous contrôle
d'une élévation du niveau plasmatique d'estradiol (pour
les Mammifères) qui provoque la décharge de GnRH
responsable de la décharge des gonadotropines
hypophysaires.
Il semble que ce soit la quantité de gonadotropines
disponibles par rapport au besoin des follicules en croissance qui
détermine pour une espèce son taux d'ovulation.
Des techniques de perfusion d'ovaires mûrs de Mammifères
in vitro ont permis de préciser les déterminismes de
l'ovulation : la première étape est la dissociation
des cellules folliculaires essentiellement sous l'action de la
FSH qui provoque une sécrétion d'acide hyaluronique par
les cellules de la granulosa, libérant ainsi l'ovocyte dans
l'antrum. Le gonflement du follicule est rendu possible par la
dissociation des fibres de collagène des membranes externes de
l'ovaire (albuginée) et de la thèque externe. La
rupture du follicule semble résulter non d'une
augmentation de la pression dans la cavité folliculaire mais
d'un amincissement et d'une dissociation des cellules de la paroi du
follicule au niveau de l'apex, zone protubérante du follicule
mûr. Quelques heures avant l'ovulation on observe une
vasoconstriction générale de l'ovaire qui
provoque la mort des cellules épithéliales au niveau de
l'apex. C'est la dégradation des cellules
épithéliales (enzymes lytiques
libérées...) qui provoquerait la rupture de
l'albuginée et des thèques sous-jacentes. On observe
alors une fuite de liquide folliculaire qui fait baisser la pression
hydrostatique du follicule et c'est cette dernière qui
provoquerait l'expulsion complète de l'ovocyte et des cellules
périovocytaires (formant la corona radiata) par une
contraction de l'ovaire. L'ovulation est actuellement
considérée comme un mécanisme relevant de la
réaction inflammatoire localisée. En effet, on y
retrouve tous les facteurs chimiques (histamine, prostaglandines,
bradykinine...) entraînant une augmentation du flux sanguin (le
volume de sang de l'ovaire est multiplié par sept après
la décharge ovulante chez la ratte), une augmentation de la
perméabilité vasculaire, arrivée de nombreux
phagocytes.... Cette compréhension est évidemment
à la base des connaissances sur les mécanismes de
régulation artificielle par l'homme des ovulations...
On peut qualifier le corps jaune de glande endocrine
éphémère, plus ou moins structurée
(l'expression est tirée du Reproduction des
vertébrés, p 120, références
citées en début de page). Chez les Primates le
corps jaune a pour fonction essentielle de synthétiser des
hormones stéroïdes, essentiellement la
progestérone, mais aussi de nombreuses hormones ou facteurs de
croissance peptidiques (ocytocine, relaxine, IGFs, inhibines,
prostaglandines...). Le corps jaune inhibe aussi la
folliculogénèse. Le développement du corps jaune
n'est pas du à une multiplication cellulaire mais à
l'hypertrophie des cellules de la granulosa dont le contenu en
ADN augmente et qui deviennent polyploïdes. Les cellules
de la thèque interne (chez la femme et le singe rhésus)
restent groupées en îlots ou en travées
associées à du tissu conjonctif et forment des septa
entre les amas de cellules lutéales. C'est la LH qui semble
contrôler le maintien du corps jaune chez la femme. La
régression intervient en absence de fécondation au bout
d'environ 2 semaines. On a isolé un facteur de
régression du corps jaune : facteur de lutéolyse (du
groupe des prostaglandines) chez la brebis. Toutes les
expériences tentant de démontrer le rôle des
hormones lutéotropes (LH) sur la lutéolyse se sont
soldées par des échecs et l'on pense maintenant que
celle-ci est bien sous la dépendance d'un facteur
sécrété par l'utérus, sauf chez la femme
et les Primates, chez lesquels l'hystérectomie (ablation de
l'utérus) est sans effet. La question est donc encore ouverte.
Des facteurs embryonnaires bloquant la lutéolyse en cas de
gestation ont été isolés chez des Primates : le
plus connu est l'hCG (human chorionic gonadotropin),
sécrété par le trophoblaste (partie la plus
externe de l'embryon en cours de développement qui participera
notamment à la formation du placenta). Il est
sécrété en grande quantité pendant les
deux premiers mois de vie embryonnaire, il inhibe la
lutéolyse, stimule les sécrétions
stéroïdiennes (progestérone surtout) du corps
jaune gestatif et du placenta et stimule l'hypertrophie et la
relaxation des cellules musculaires du myomètre utérin
et des vaisseaux. Le corps jaune persiste chez les Primates pendant
toute la durée de la gestation.
En résumé :
Le point de vue fondamental : chez tous les
vertébrés la régulation du fonctionnement des
gonades (ovaires et testicules) par le système nerveux
central se fait par l'intermédiaire d'une glande
endocrine interposée entre la gonade et le SNC :
l'hypophyse. Cette régulation se fait bien
sûr en synergie avec la fonction endocrine propre des gonades
qui intervient sur leur propre développement et sur les autres
organes de la reproduction. Ce qui peut être illustré
par ce petit schéma :
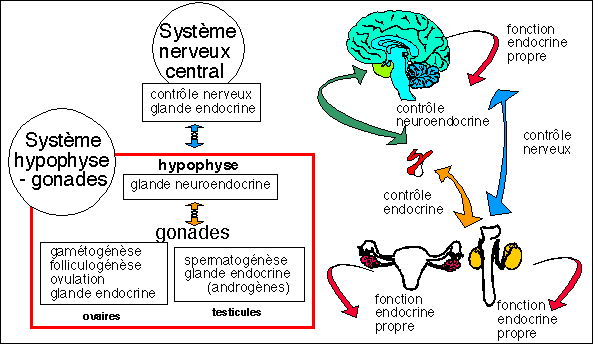
Les relations entre le SNC et les gonades : un glande s'interpose :
l'hypophyse...
on peut donc considérer le système
hypophyse-gonades comme un tout...
Tous les mécanismes
intervenant dans la mise en place de la fonction gonadique
comme dans sa modulation en fonction de paramètres
environnementaux ou internes semblent aboutir en fin de compte
à une accélération / ralentissement de la
fréquence ou à une augmentation / diminution de
l'amplitude de la libération hypophysaire des hormones
gonadotropes.
b. la régulation adaptative - la réponse
adaptative des gonades au milieu
(travail social de reproduction des gonades)
Le contrôle du système hypophyse-ovaires fait
intervenir une sécrétion pulsatile de GnRH,
neurohormone hypothalamique qui agirait sur les cellules
antehypophysaires sécrétant le LH et la FSH.
Des dosages de GnRH réalisés chez le macaque
rhésus mettent en évidence à 14 mois (stade
impubère) des taux extrêmement faibles (quelques
unités en pg/mL pour une durée d'enregistrement de 10
minutes) et continus pendant la journée. Vers 25 mois (stade
prépubertaire) des pics (de plusieurs dizaines de pg/mL
pendant 10 min) apparaissent le matin très tôt, en fin
d'après midi et la nuit. A 38 mois (puberté) des pics
de l'ordre de la dizaine de pg/mL sont enregistrés de
façon assez peu régulière, environ toutes les
deux heures et avec des maxima le soir et la nuit.
MAIS... ce n'est pas encore si clair... voici quelques
éléments d'études plus récentes:
le GnRH est un
décapeptide dont les 9 formes isolées différent
par un à cinq aa chez différents
vertébrés. Il est sécrété par des
neurones de l'aire préoptique et de l'hypothalamus
médian, mais d'autres neurones du cerveau semblent aussi
pouvoir le sécréter. La
sécrétion est pulsatile mais tous
les facteurs de l'environnement (éclairement,
température...) et les facteurs sexuels internes (hormones
sexuelles...) agissent aussi sur la pulsatilité de la
libération de GnRH. La puberté correspond à une
augmentation de fréquence des pulses alors que la gestation ou
l'allaitement correspondent à une baisse de fréquence.
Le "générateur de pulses" n'a pas été
identifié et l'on pense pouvoir invoquer une
sécrétion rythmique autonome et une système de
synchronisation. Mais ceux-ci restent à découvrir... Le
GnRH se fixe a un récepteur membranaire de type
glycoprotéine à la surface des cellules gonadotropes.
Le complexe hormone-récepteur est endocyté après
micro-agrégation (regroupement des récepteurs deux par
deux) et le GnRH est libéré dans la cellule. Il semble
agir au niveau du cytoplasme et du noyau et provoque rapidement une
libération des gonadotropines et à plus long terme une
stimulation de la synthèse des gonadotropines (par l'AMPc).
La sécrétion pulsatile des hormones gonadotropes
semble résulter directement de la sécrétion
pulsatile du GnRH.
Chez tous les vertébrés des deux sexes la castration
est suivie d'une augmentation de la fréquence et de
l'amplitude des pulses de GnRH entraînant une
élévation des niveaux plasmatiques des gonadotropines.
L'administration des stéroïdes sexuels entraîne un
retour plus ou moins complet vers les niveaux bas observés
avant la castration.
Chez la femme, la fréquence des pulses est plus
élevée dans la première partie du cycle sexuel
(phase folliculaire) que dans la seconde (phase lutéale).
On a isolé des récepteurs aux stéroïdes
sexuels au niveau des cellules hypophysaires mais par contre les
neurones à GnRH n'en possèdent pas (l'action des
stéroïdes sexuels pourrait se faire par
l'intermédiaire d'autres neurones sensibles qui stimuleraient
ensuite les neurones à GnRH par leurs neuromédiateurs).
Par contre, in vivo, et chez tous les vertébrés
étudiés, on a mis en évidence une diminution du
contenu en GnRH des neurones hypophysaires à chaque fois que
la concentration en gonadotropines augmente (castration,
décharge ovulante, variations saisonnières...). D'une
façon générale on peut dire que les
neuromodulateurs de la libération de GnRH agissent soit en
dépolarisant fortement la membrane, ce qui provoque la
libération de GnRH, soit en la dépolarisant faiblement,
ce qui favorise mais ne provoque pas cette libération, soit
enfin en hyperpolarisant la membrane, ce qui inhibe la
libération de GnRH.
La GnRH stimule l'expression de ses propres récepteurs (on
parle d'up-regulation). L'administration continue de GnRH provoque
chez l'homme une désensibilisation par diminution du nombre de
récepteurs (down-regulation).
D'autre part il existe des contrôles en retour ou
rétrocontrôles de l'ovaire sur l'antehypophyse et sur
l'hypothalamus. On pense que les œstrogènes, en dessous
d'un certain seuil inhiberaient la sécrétion des
hormones antehypophysaires (rétrocontrôle
négatif), alors qu'au-dessus de ce seuil, elles les
stimuleraient (rétrocontrôle positif). La
progestérone semble inhiber la sécrétion
d'hormones antehypophysaires de type LH et FSH... Il existe d'autres
hormones (inhibine par exemple) qui ont été
isolées de l'ovaire. Il existe aussi des facteurs chimiques
agissant à courte distance comme dans le système
immunitaire...
Chez de nombreuses femelles de Primates et de mammifères en
général la période postovulatoire s'accompagne
de profonds changements comportementaux (agitation, signaux visuels,
olfactifs, sonores...), la femelle n'acceptant l'accouplement que
pendant cette période. On qualifie cette période de
rut ou d'œstrus, le cycle sexuel prenant le nom de
cycle oestrien car ce changement de comportement de la femelle
est de loin le phénomène sexuel le plus visible. Chez
de nombreuses espèces de mammifères domestiques les
cycles oestriens se succèdent pendant toute l'année
sans interruption (souris, ratte, vache, lapine, truie) alors que
chez certaines espèces domestiques et la plupart des
espèces sauvages, les périodes d'œstrus sont en
petit nombre (jument, brebis, chienne, chèvre, chatte...) et
correspondent habituellement au printemps et à l'automne...
Les durées des cycles oestriens varient de quelques jours (4-5
jours chez la souris) à par exemple 21 jours chez la vache ou
la truie.
L'action des stéroïdes sexuels au niveau des cellules
hypophysaires (rétrocontrôle) semble se faire
notamment par diminution du nombre de récepteurs à la
GnRH. Mais, in vivo comme in vitro, l'estradiol
semble avoir un effet biphasique : après avoir
inhibé la réponse des cellules hypophysaires à
la GnRH, il l'augmente. La progestérone semble ne pas avoir
d'effet propre sur l'hypophyse, la synthèse de ses
récepteurs dépendant de l'estradiol.
Les nombreux facteurs cités plus haut exercent aussi des
rétrocontrôles variées soit sur les neurones
à GnRH soit sur l'hypophyse...
La décharge ovulante de GnRH ne peut se produire que si
le système nerveux central et l'hypophyse ont
été soumis à l'action de l'estradiol, à
un niveau suffisamment élevé et pendant un temps
minimum. Chez les Mammifères, la présence d'un niveau
élevé de progestérone empêche une
décharge ovulante de se produire (cette
propriété est utilisé dans la conception des
contraceptifs chimiques de type dérivés progestatifs).
Chez la femelle de Primate normale la décharge ovulante de LH
se produit après la décharge de GnRH si les cellules
hypophysaires on été soumises à de fortes
concentrations d'estradiol. Si l'on remplace l'action de
l'hypothalamus (chez un animal opéré) par une injection
pulsatile de faible taux de GnRH on observe par contre une
libération importante de LH (et FSH) du fait de
l'élévation du taux d'estradiol
sécrété par les follicules en croissance sans
qu'il soit nécessaire d'administrer une forte dose de GnRH. Il
y a donc aussi une sensibilité directe de l'hypophyse à
l'estradiol.
II. La fécondation, travail de
coopération
Si l'on s'en tient au niveau des cellules, la fécondation
chez l'homme est la rencontre suivie de la fusion en milieu liquide
(les sécrétions de l'oviducte) de deux gamètes :
le spermatozoïde et l'ovocyte au stade II. Elle comprend les
événements principaux suivants :
- reconnaissance spécifique des gamètes puis
fusion.
La reconnaissance spécifique se fonde sur des
barrières comportementales, anatomiques et
moléculaires amis aucune de ces barrières n'est
absolue. On observe ainsi des zygotes issus de la
fécondation de deux espèces différentes, la
plupart du temps sans développement embryonnaire. Quelques
rares cas donnent des individus viables (mulet...) mais
stériles.
Les principales étapes sont : la reconnaissance entre
l'enveloppe de l'ovocyte et la membrane du spermatozoïde
(nécessitant la capacitation chez les mammifères),
la rupture de l'acrosome (réaction acrosomique) suivie de
la désagrégation de la membrane pellucide, la
reconnaissance des membranes plasmiques de l'ovocyte et du
spermatozoïde suivie de leur fusion (latérale chez
l'homme), enfin, début de pénétration du
noyau du spermatozoïde dont l'enveloppe nucléaire
commence à disparaître dès le début de
la fusion.
La capacitation, réarrangement des protéines de
surface de la membrane de l'apex du spermatozoïde (membrane
périacrosomiale), est une particularité des
Mammifères. Elle se fait habituellement lors du passage
à travers les voies génitales femelles (col de
l'utérus). Elle semble nécessaire essentiellement
étant donnée la durée de transit des
spermatozoïdes dans l'épididyme et les canaux
déférents, ce qui impliquerait des mécanismes
de protection permettant la survie des spermatozoïdes dans
les voies génitales mâles et le vagin.
- réponse du zygote à la suite de la fusion de
l'ovocyte avec le spermatozoïde.
L'activation du zygote se manifeste immédiatement par la
libération des granules corticaux (réaction
corticale) qui en libérant leur contenu à
l'extérieur du zygote forment la membrane de
fécondation qui est le résultat d'une modification
de la membrane pellucide. La reprise de le méiose est aussi
immédiate avec expulsion du deuxième globule
polaire. les changements métaboliques au niveau du
cytoplasme du zygote sont la cause ou les témoins de
l'activation (augmentation du Ca2+ intracellulaire libre et
modification du potentiel membranaire principalement).
La décondensation du noyau du spermatozoïde en fin de
pénétration se poursuit par la synthèse d'une
nouvelle enveloppe formant ainsi un pronucléus mâle
de diamètre important. Après l'expulsion du
deuxième globule polaire, les chromosomes ovulaires se
décondensent aussi et s'entourent d'une enveloppe
nucléaire pour former le pronucléus femelle. La
pièce intermédiaire du spermatozoïde contenant
les mitochondries et le centrosome (deux centrioles) reste au
voisinage du pronucléus mâle. Les mitochondries
mâles dégénèrent. Le centrosome
organise un aster de microtubules (spermaster) qui est responsable
des mouvements relatifs des pronucléi.
Enfin, il y a reprise du cycle cellulaire (phase G1 de quelques
heures, S de 1 à 3 heures, G2 de 24 heures), la disparition
des enveloppes des pronucléi n'intervenant qu'après
l'apparition des chromosomes (condensés) qui ne se
mélangent pas. Cette absence de mélange des
chromosomes paternels et maternels semblent durer toute la
vie.
Mais la fécondation peut aussi être
étudiée au niveau des organes (voir chapitre
précédent). En effet, la fécondation humaine ne
peut avoir lieu que en un certain point des trompes, à l'aide
d'organes copulateurs (pénis et vagin) au fonctionnement
réglé et coordonné. On a récemment mis en
évidence chez des mammifères (?) l'importance de la
température au niveau des cormes utérines et du
début des trompes. Les spermatozoïdes seraient
attirés par la chaleur de la zone de fécondation
(39°C par rapport aux 37°C de l'utérus). Ce
thermotactisme (un tactisme est un déplacement orienté,
ici par la température) reste mystérieux. Un
chimiotactisme (attraction chimique) prendrait le relais au voisinage
de l'ovocyte qui sécréterait des substances attractives
(voir Des spermatozoïdes sensibles à
la température, Pour la Science, 305, mars 2003, p
15).
Enfin, la fécondation, étudiée au niveau des
individus, est réalisée entre sexes
séparés, pendant le périodes et aux âges
d'activité sexuelle. Au niveau des populations, elle
dépend plus de caractères éthiques,
économiques, familiaux....
Tous ces paramètres ont parfois tendance à
être volontairement écartés car ils peuvent
paraître non nécessaires à certains. En effet, la
fécondation in vitro permet d'obtenir un zygote. Mais le
zygote n'est rien s'il ne se divise pas pour donner un massif
embryonnaire. De même l'embryon n'est rien s'il ne s'implante
pas dans un utérus.... Le travail de reproduction est global
et finalisé ou il n'est que manipulation technique du
vivant.
III. L'hérédité
L'hérédité est une question qui DOIT
être approfondie mais qui débouche sur des
théories. Pour tous ceux qui le souhaitent je vous conseille
de consulter les pages sur l'hérédité
du site lié à celui-ci.
La question peut se formuler de différentes façons
:
Qu'est-ce qui est transmis des parents aux enfants
?
Quelle est la part de l'inné et de l'acquis dans le
développement de l'enfant, dans son comportement, ses
capacités ?
La réponse quasi automatique fait référence
à un imaginaire social très fort : le patrimoine
héréditaire... pour simplifier : les
gènes. Je vous invite à lire le livre de Dorothy
NELKIN et Susan LINDEE (1998, Belin, Coll. Débats ): La
mystique de l'ADN. (ou au moins l'article du magazine "La
Recherche" qui y fait référence : Du gène
comme icône culturelle, Dorothy Nelkin et M. Susan Lindee,
La Recherche, 311, juillet-août 1998, 98-101).
En effet, si l'on vous questionne, vous finirez bien par revenir sur
votre première réponse et dire qu'effectivement les
gènes, ou plus exactement les chromosomes, ne sont bien
sûr pas les seuls éléments matériels
transmis sur lesquels l'hérédité peut se fonder
: le cytoplasme et l'environnement cellulaire sont certes tout aussi
importants. Mais quand il s'agit de proposer une théorie vous
vous trouvez devant l'unique théorie chromosomique de
l'hérédité, la seule qui soit enseignée
dans le secondaire. Le concept de programme
génétique repose lui aussi totalement sur des
positions philosophiques issues des découvertes de Morgan
(voir histoire
de la génétique). D'autres théories sont en
train de se faire une place dans les milieux scientifiques et
philosophiques mais ce sont des phénomènes assez lents.
Je vous ai parlé de la théorie de Rosine Chandebois qui
est à mon avis très intéressante mais il y en a
certainement d'autres que je ne connais pas.
Que doit-on, que peut-on enseigner à l'école
élémentaire sur l'hérédité ? Le
premier élément est de se garder soigneusement de
présenter les caractères de l'homme (couleur des yeux,
taille....) comme des caractères mendéliens (voir
les éléments de réflexion toujours dans la
même page
sur la génétique pour ceux qui sont
intéressés). La deuxième précaution
est de bien présenter l'hérédité comme un
phénomène biologique global (travail de
reproduction) et non une simple transmission mécanique,
matérielle, de particules déterminées. La part
de l'inné et de l'acquis, devrait pouvoir être
présentée de façon encourageante pour l'enfant,
même s'il n'y a pas de réel support biologique à
telle ou telle théorie : les capacités de chacun sont
différentes mais tout enfant à un patrimoine moyen
suffisant qui, mis à profit au bon moment de son
développement, est susceptible de lui permettre de
s'épanouir au sein de la société. On peut aussi
dire que chacun a un point fort qui doit être
valorisé.
Une question liée de façon étroite à
l'hérédité est celle de la définition du
sexe. Sans aborder de front la question en prenant l'exemple de
l'homme, on peut se contenter de présenter la séquence
acceptée actuellement par l'ensemble de la communauté
scientifique : le sexe génétique ou
chromosomique est définit par une paire de gonosomes
(hétérochromosomes c'est-à-dire non homologues
sur toute leur longueur) : XX chez la femme et XY chez l'homme. Ce
sexe génétique est à l'origine du
déterminisme du sexe phénotypique ou
somatique sur lequel repose la morphologie de l'individu
(organes sexuels externes et internes (dimorphisme sexuel),
caractères sexuels secondaires...voire le comportement). Ce
passage du sexe génétique au sexe phénotypique
est le résultat d'interactions hormonales (et autres
médiations et contacts) complexes mis en place chez l'embryon
puis le fœtus et passe par un stade intermédiaire qui est
le sexe gonadique ou gamétique qui
contrôle le type de gamètes produits.
Cependant tout n'est pas simple : si l'on regarde d'un peu plus
près les chromosomes sexuels ont peut être surpris des
résultats obtenus qui sont loin d'être simple à
interpréter. En voici quelques-uns juste pour vous
surprendre:
- L'appariement très partiel des
gonosomes en prophase de première division de méiose
permet de les repérer. Les spermatozoïdes peuvent
être triés automatiquement selon leur "sexe"
après coloration vitale par cytométrie de flux
(que l'on s'entende : un spermatozoïde n'a pas de
sexe, mais son chromosome sexuel est le déterminant
quasi obligatoire du sexe du fœtus auquel il peut donner
naissance après fécondation avec un ovocyte
mûr) du fait de la différence entre les
chromosomes sexuels dont un seul exemplaire est bien sûr
porté normalement par spermatozoïde.
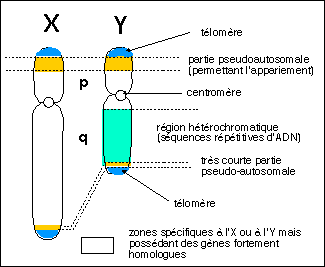
Une cartographie très simplifiée des chromosomes
sexuels (gonosomes)
(d'après "La reproduction des
vertébrés", Masson, 1998).... on est loin
de la reproduction de la cartographie schématique des
ouvrages scolaires.... en fait je ne dis pas que celle-ci est plus
vraie je veux simplement montrer qu'une cartographie peut
être manipulée selon ce que l'on a à dire....
Ici mon message est de montrer que la génétique
"scolaire" ou même "universitaire" qui "fabrique de toutes
pièces" des exercices de génétiques en
plaçant des gènes dans la partie propre de l'X ou
celle de l'Y ne se basent pas sur des résultats de la
cytologie chromosomique mais sur une représentation qui
date de plus de dix ans. Pour voir quelques techniques de marquage
des chromosomes et des données de génétique
récentes, je vous invite à visiter les pages du
cours de terminale de
spécialité.
- on connaît des cas (rares) d'inversions sexuelles
chez l'homme : le sexe phénotypique est opposée au
sexe génétique normalement attendu après
l'analyse du caryotype. On explique par exemple le syndrome du
testicule féminisant chez l'homme de sexe mâle mais
de phénotype féminin par mutation d'un codon du
gène codant pour le récepteur aux androgènes
(ce qui conduit à rendre insensibles à la
5aDHT (métabolite,
biologiquement actif, de la testostérone) les cellules
cibles des testicules, du tractus génital, du
système nerveux....).
IV. les applications médicales et
non médicales
Il n'est pas rare d'entendre citer parmi les applications
médicales ... la contraception. Si l'on peut dire que toute
médecine soigne (la médecine
curative soigne les effets;
la médecine préventive, soigne les
causes) je crains par contre qu'il n'y ait rien
de médical à ces actes de contrôle de la
reproduction, sauf dans certains cas limites de médecine
préventive pour laquelle une conception mettrait gravement en
danger la vie de la mère. Par contre, ils doivent sans aucun
doute se faire sous contrôle médical. Leur choix
relève de l'éthique et non de la médecine.
Le contrôle des naissances n'est certes pas un acte nouveau et
il semble rapporté dans des temps très anciens.
Qu'apportent les connaissances actuelles ? Par la prise de
molécules chimiques dérivées des hormones
naturelles et à des doses très éloignées
des doses biologiques, sous forme de comprimés (pilules), ou
d'injections (sous-cutanées), une femme peut soit
empêcher l'ovulation, soit empêcher la fécondation
(mécanismes contraceptifs, soit enfin empêcher la
nidation (mécanisme contragestatif). En dernier lieu
des molécules artificielles peuvent aussi provoquer un
avortement (mécanisme abortif). On est loin de bien
connaître les modifications physiologiques liées
à l'emploi des contraceptifs chimiques et les femmes qui les
utilisent assez massivement dans certains pays comme la France, on
servi à ajuster les doses, qui ont fortement diminué en
40 ans, et prévoir quelles sont les associations dangereuses
avec certaines maladies et certains médicaments.... On est
loin d'un cours de biologie.
Par contre il existe de réelles applications
médicales : soigner des déficiences ou des
incapacités à la reproduction sont bien des actes
médicaux, mais je n'ai aucune compétence pour les
enseigner. On notera qu'ils relèvent aussi de
l'éthique, comme tout acte de l'homme. Le désir
d'enfant, légitime, de certains couples, est poussé
à un tel paroxysme que l'enfant est instrumentalisé,
n'étant plus que l'objet du désir de ses parents. Dans
ce domaine encore la psychologie, la sociologie avec son volet
économique essentiel et toujours l'éthique sont les
sciences adéquates, certainement pas la biologie qui n'a pas
grand chose à voir avec les techniques comme la FIVETE
(fécondation in vitro et transfert d'embryon).
Comme vous pouvez être interrogés sur ces questions
voici quelques éléments de réponse issus de "La
reproduction des vertébrés" (Masson, 1998).
- la fécondation in vitro (FIV) nécessite avant
tout le respect de la température corporelle (37°C)
mais le reste des conditions est largement empirique. On note
qu'il est nécessaire d'avoir une très forte
concentration de spermatozoïdes (105) alors que
dans les trompes seuls quelques spermatozoïdes arrivent au
voisinage de l'ovocyte. C'est comme cela que l'on explique les
nombreux cas de polyspermie observés lors des FIV.
- la technique récente de FIV par ICSI (intracytoplasmic
sperm insemination) réalisée depuis 1992 donne des
taux de grossesse de 30% (au lieu des 20% par la technique du
transfert) mais les anomalies génétiques
(délétions au niveau du locus AZF du chromosome Y
situé dans le bras p) qui altèrent la formation des
spermatozoïdes apparaissent spontanément et son
transmissibles à la descendance.
- dès 1952 (Briggs et King), on a réalisé
des transplantations nucléaires sur des œufs de
grenouille activés puis énucléés dans
lesquels étaient injectés un noyau venant d'une
cellule d'une blastula: on obtenait ainsi un clone de grenouilles.
Les cas de clonage sont à différencier de la
parthénogenèse (développement sans
fécondation mais souvent avec doublement naturel ou
artificiel du nombre de chromosomes...) qui ne conduit jamais
à un embryon viable chez les mammifères. Bataillon
en 1910 réalisait des débuts de développement
par parthénogenèse provoquée chez la
grenouille en piquant l'ovocyte mais il lui fallait ensuite
injecter une hématie (nucléée chez les
amphibiens) dans l'ovocyte afin de passer au stade de la
segmentation. Les récents résultats (?)
médiatisés (!!!) sur Dolly sont encore trop
discutés pour faire l'objet de ce cours. Je vous renvoie
aux dossiers spéciaux du magazine La
Recherche sur ce sujet.
Remarque:
Un petit article récent d'Israël Nisland
(Eugénisme et diagnostic ; Quels sont les objectifs
réels de l'échographie des femmes enceintes ? Pour la
Science, 269, mars 2000, p8) met le doigt sur la finalité des
examens échographiques.
« La loi française admet aujourd'hui
deux types d'avortement : l'interruption volontaire de grossesse, qui
se pratique jusqu'à la douzième semaine de grossesse,
sans justification de la femme, et l'avortement médical
prescrit par les médecins à partir
d'échographies ou d'analyses. Dans ce cas, un médecin
est conduit à proposer un avortement si l'atteinte foetale est
très grave ou si la grossesse met la vie de la mère en
danger. L'avortement médical est ainsi une forme d'euthanasie
active.
Or les progrès récents ou prévus du
dépistage prénatal et la prolongation du délai
légal d'avortement nous mènent vers une situation
où un eugénisme institutionnalisé serait
possible : les femmes sauraient, avant la fin du délai
légal d'interruption volontaire de grossesse, que leur enfant
a une anomalie même légère - par exemple, une
fente labiale ou un doigt en moins - et elles décideraient une
interruption volontaire de grossesse sur la base des informations
données par le médecin. Examinons pourquoi cette
situation serait grave.
Jusqu'en 1998, les avortements
thérapeutiques de grossesse étaient prescrits par les
seuls échographistes, qui étaient alertés par
des observations effectuées au cours de l'échographie
de dépistage prénatal. Une remarque du médecin
à sa patiente transformait parfois une grossesse
désirée en un désir d'avortement, et les
médecins étaient alors poussés par les femmes
à des attitudes contre leur avis et contre leur conscience. La
situation était aggravée par le risque de poursuites
judiciaires par des parents, lorsque le pronostic de la maladie de
l'enfant avait été sous.estimé par le
médecin : il est arrivé que des médecins,
craignant ces poursuites, accèdent à la demande
d'interruption médicale de la grossesse alors que celle-ci
pouvait être évitée.
La loi de 1994, suivie de décrets en 1998, impose aujourd'hui
que les dossiers d'avortement médical ne se décident
plus dans le cabinet de l'échographiste, entre le seul
médecin et sa patiente. Quand un avortement médical est
demandé, le dossier médical est soumis à un
«centre pluridisciplinaire de diagnostic prénatal»,
où une commission pluridisciplinaire accepte ou refuse
l'avortement. Ces centres sont composés
d'échographistes, de généticiens, de
médecins des diverses disciplines, de psychiatres... Les
dossiers sont discutés publiquement et collectivement.
Ainsi, les échographistes échappent aux pressions des
patients et sont protégés du risque
médico.légal : les juges n'ont plus la
possibilité de les condamner injustement pour des fautes
qu'ils n'ont pas commises. Doit-on rappeler que la médecine a
une obligation de moyens, et non de résultats ? Qu'elle doit
soigner, mais ne peut pas toujours guérir ?
Ces centres doivent aujourd'hui prendre des
décisions sur des bases claires, mais ce serait une erreur
qu'ils en viennent à décider ce qui est normal et ce
qui ne l'est pas. L'institutionnalisation d'une norme fait
évidemment peur, car elle pourrait conduire à un
eugénisme d'état. En outre, que cherche-t-on à
voir lors de l'examen ? Le diagnostic prénatal n'a toujours
pas d'objectifs bien définis. Je maintiens que
dépister l'absence d'un doigt à une main n'apporte rien
à la grossesse : l'enfant qui naîtra ne sera pas moins
homme. La médecine n'a pas décidé ce
qui est grave et ce qui ne l'est pas, et la technique médicale
a progressé plus vite que la réflexion sur
l'utilisation de cette technique.
Pis encore, les progrès de l'échographie relancent les
risques de dérive. Aujourd'hui, les échographies
vaginales en trois dimensions dépistent d'éventuelles
anomalies avant le délai légal d'interruption
volontaire de grossesse : les femmes risquent de décider de
telles interruptions de grossesse sur la foi d'échographies,
comme avant que ne soient créés les centres de
diagnostic.
De surcroît, des discussions sont en cours pour repousser la
date légale d'interruption volontaire de grossesse. Si l'on
décidait finalement un allongement du délai, je
proposerais de repousser aussi la date d'échographie de
grossesse (aujourd'hui fixée à 13 semaines de
grossesse).
Revenons à la question centrale : «
À quoi sert le diagnostic prénatal ? ». Selon moi,
ce dépistage n'a de sens que s'il engendre une
modification de la conduite obstétricale : il faut
dépister les malformations létales ou très
graves, afin de traiter in utero celles qui peuvent l'être, ou
opérer à la naissance et sauver l'enfant.
L'acte thérapeutique modifie la répartition des
gènes dans la population, mais ce « dysgénisme
actif » est vieux comme la médecine : la
césarienne est un tel dysgénisme. Ce dysgénisme
relève bien de la mission médicale : soigner. Au
contraire, l'eugénisme national est dangereux : les
médecins n'ont pas oublié que certains des leurs, tels
Alexis Carrel ou Charles Richet, préconisaient de tuer les
voleurs, les handicapés, d'organiser la société
pour l'es gens sains et non, clamaient-ils, pour des individus
malades.
La technique peut être perfectionnée, mais les
médecins doivent se demander à quoi ils servent.
L'éthique doit précéder la technique ; on doit
réfléchir pour savoir comment on la met en oeuvre et,
même, si on la met en oeuvre. Jamais je n'admettrai le point de
vue des techniciens qui veulent faire de belles images sans
réfléchir aux conséquences de leurs actes. Les
échographistes doivent être des médecins, pas des
manipulateurs : science sans conscience... Et le foetus n'est pas un
produit de transformation ; c'est un être humain. J'appelle au
débat. » Israël NISAND est
obstétricien au Centre hospltalo-universitaire de Strasbourg,
qui abrite le Centre pluridisciplinaire de diagnostic prénatal
d'Alsace.
V. Ethique de la reproduction.
Pour une définition de l'éthique je renvoie au cours
de spécialité de
terminale.
L'éthique est la science des
actes humains (science de l'agir) considérés selon leur
orientation à la fin dernière de l'homme
(moralité). La moralité d'un acte humain est son
orientation à la fin dernière de l'homme.
La fin de l'homme comprend sans doute aucun la multiplication
de l'espèce, même si un individu peut s'y
soustraire. La reproduction doit donc être ordonnée
à sa fin : la multiplication de l'espèce. Toute
pratique qui cherche une autre fin avec les moyens physiologiques
reproducteurs est une pratique détournée de sa fin.
Mais il est certain que la reproduction n'est pas non plus la fin
dernière de l'homme. L'homme peut donc contrôler,
volontairement, une fonction naturelle.
Il y a une différence entre contrôle et utilisation dans
un but autre que la génération des enfants. On peut
contrôler une fonction sans pour autant la détourner de
sa fin.
- Un contragestatif abortif (pilule RU 486) est clairement
contraire à l'éthique, non pas de la fonction
reproductrice mais de la vie elle-même , car il tue un
embryon vivant et le respect de la vie est le premier devoir de
conscience de l'homme (voir cours de
spécialité ).
- Un contraceptif chimique, de type pilule microdosée par
exemple, qui modifie essentiellement les cycles sexuels hormonaux
féminins, s'il est utilisé pour empêcher une
fécondation au cours d'un acte reproducteur, est clairement
utilisé afin de détourner un acte de sa fin
première. A ce titre l'utilisation d'un préservatif
possède la même qualification éthique. La fin
de l'acte sexuel "protégé" étant alors la
recherche du plaisir sexuel (mais certainement pas la seule... on
arrive ici dans des domaines très intimes de la conscience
humaine où la qualification morale d'un acte dépend
beaucoup de la connaissance de la loi naturelle et des fins
réelles de cet acte... il est clair que l'on sort du
domaine de ce cours).
En fin de compte la profonde cohérence de l'éthique
de la reproduction est l'affirmation claire et dénuée
de tout tabou ou sens caché que l'homme a l'honneur insigne de
pouvoir toujours orienter grâce à sa liberté,
tous ses actes, vers la fin la plus haute que lui désigne son
intelligence et sa volonté.
retour accueil
résumé, formulation
par cycle
Annexe
... généralisation sur la
fonction endocrine
et sur les
substances informatives ...
à partir de l'exemple du travail
de reproduction
Il semble nécessaire de faire une petite mise au point
concernant les substances chimiques dans l'organisme.
IL NE FAUT PLUS RAISONNER EN TERMES DE MOLECULES MAIS EN TERMES DE
FONCTIONS.
Que veut dire une substance informative ? Que signifie une
information dans le vivant ?
Je vous renvoi aux pages sur le vivant .
En quelques mots, on se réfère ici au travail du vivant
qui peut être décrit en termes de matière,
d'énergie et d'information (ou d'ordre).
Toute information n'est pas supportée par une
molécule (on peut très bien parler de l'information
spatiale (position, environnement d'une cellule) ou temporelle
(histoire d'une cellule) ou encore d'information de
température, de pression ou de toute autre paramètre
qu'il est quasiment impossible de relier à une ou à un
ensemble de molécules, du fait de la complexité des
déterminismes). Si l'on établit
d'intéressantes comparaisons avec les théories de
l'information, je crois avoir entendu une phrase qui avait
attiré mon attention : "le message c'est le
média". Si l'on reprend aussi l'étymologie du terme
communiquer (du latin communicare : mettre en relation,
mettre en commun, partager, participer à ...) ou du verbe
informer (du latin informare : façonner, former,
mais aussi, enseigner, instruire, ou encore, former une idée,
former un concept de l'esprit, concevoir...). Le terme
média est aussi intéressant : il vient
semble-t-il de l'américain mass-media (masse-media)
forgé à partir du latin media (en bas latin,
mediatio signifie le moyen et vient de l'adjectif medius,
a, um : du milieu, moyen ou médiocre ; et du nom
medium,ii : le milieu, l'espace intermédiaire, ce qui
est du domaine commun, public..). Une molécule informative
est considérée comme telle parce qu'on lui attribue un
certain effet, une finalité. Son message c'est son être,
c'est elle-même...(je me souviens de discussion de
spécialistes de l'information qui insistaient pour faire
comprendre à leurs étudiants-présentateurs
publics que ce qui compte ce n'est pas le message c'est leur
être, leur personnalité...).
Toute substance chimique
informative peut être qualifiée de
médiateur.
Une substance informative est émise
(libérée), transmise (transportée), reçue
(réception) et doit être suivie d'un effet .
Tout médiateur pour pouvoir agir implique la présence
de récepteurs plus ou
moins spécifiques sur les cellules
cibles.
On distingue alors :
- les neuromédiateurs : synthétisés par
des neurones, libérés sous contrôle nerveux (PA)
au niveau des extrémités axonales la plupart du temps
synaptiques
- les hormones ou médiateurs endocrines :
synthétisés et sécrétés par des
cellules endocrines en permanence, libérés dans le
milieu intérieur et donc circulants à un certain
taux.
- les médiateurs paracrines : substance chimique
à diffusion locale, dans le milieu extracellulaire et
rapidement inactivé
- les médiateurs autocrines : stimulent à la
fois la cellule sécrétrice et les cellules avoisinantes
par effet paracrine.
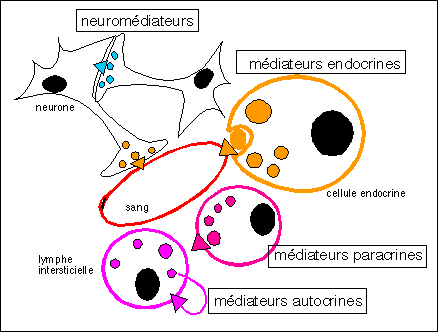
Une classification des substances chimiques informatives ou
MEDIATEURS par fonction
(les hormones sont les médiateurs endocrines)
Tous les grandes fonctions ou, en utilisant le vocabulaire qui
m'est cher, toutes les expressions du travail du vivant sont donc
sous le contrôle nerveux (contrôle rapide et
intégré au niveau des centres en fonction des signaux
sensitifs reçus, ce sont eux qui réalisent
essentiellement l'adaptation de l'organisme au milieu : le
contrôle nerveux AU NIVEAU CELLULAIRE se faisant par
libération d'un neuromédiateur). Le contrôle
nerveux s'exerce sur de nombreux organes présentant une
fonction endocrine, lui-même ayant une fonction endocrine. La
fonction endocrine n'étant qu'un aspect des communications
chimiques dans l'organisme, que l'on peut donc regrouper sous le
terme de communication par médiateurs.
quelques exemples
de...Médiateurs
substances chimiques informatives
|
|
neuromédiateurs
|
Catécholamines dérivées de la
tyrosine (aa)
|
adrénaline, noradrénaline, dopa,
dopamine
|
|
sérotonine ( 5-hydroxytryptamine), GABA (acide
gamma-aminobutyrique), acide glutamique,
acétylcholine
|
médiateurs endocrines
= hormones
|
hormones stéroïdiennes (liposolubles)
|
Testostérone, œstrogènes,
progestérone, corticostéroïdes, vitamine
D3
|
|
hormones hydrosolubles dérivées de la
tyrosine
|
catécholamines (noradrénaline,
adrénaline), hormones thyroïdiennes
|
|
hormones hydrosolubles peptidiques et
protéiques
|
petits peptides (hormones hypothalamiques de 3 à
14 aa, angiotensine (8aa), glucagon (29aa), insuline
(51aa)...)
|
|
protéines (hormone de croissance (GH), LH, FSH,
prolactine...), certaines cytokines (IL1, IL6, THF: Tumor
Necrosis Factor...)
|
|
médiateurs paracrines
|
nombreuses cytokines (IL2, interférons...),
prostaglandines (ces molécules sont
particulièrement intéressantes car elles
semblent provenir de l'acide arachidonique, issu des
phospholipides membranaires ; le médiateur est donc
libéré par la cellule émettrice
directement à partir de ses constituants
membranaires, mais après un passage sous forme de
prostaglandine cytoplasmique...)
|
|
médiateurs autocrines
|
nombreuses cytokines (lymphokines...)
|
Un essai de synthèse sur le système nerveux, la
fonction endocrine, les molécules informatives et le
système immunitaire...
Le système nerveux forme un ensemble de cellules
homogène (même origine embryonnaire). C'est le
système de contrôle, de coordination surtout
tourné vers l'extérieur, le système
d'accès à la vie sensitive, à la communication
avec le milieu extérieur, à l'adaptation au milieu, au
déplacement rapide... peut-être pourrait-on dire pour
garder une expression facilement mémorisable :
le système nerveux c'est le
système de contrôle et d'intégration de
l'adaptation de l'organisme au milieu
Le système immunitaire est de plus en plus
considéré comme l'élément essentiel
interne, tourné vers l'intérieur,
c'est-à-dire vers la défense interne mais aussi le
contrôle de toutes les fonctions internes. A chaque fois que
l'on s'intéresse aux médiateurs au niveau d'une
fonction (l'exemple du programme était la fonction ovarienne),
on retrouve les médiateurs immunitaires. Le système
immunitaire, au sens le plus large, comprenant des organes, des
cellules et surtout le MILIEU INTÉRIEUR, semble être un
bon candidat pour remplacer la notion de système endocrine. Il
y a encore certainement beaucoup de travail à faire mais on
peut déjà comprendre comment les relations entre
populations cellulaires, au cours de l'embryogenèse, comme
plus tard chez l'organisme adulte, passent par des mécanismes
que l'on a souvent considéré comme faisant partie d'un
système de défense mais que l'on pourrait voir comme un
système de contrôle, d'intégration,
réalisé par des médiateurs mais aussi par des
contacts cellulaires. La fonction endocrine ne serait plus qu'un des
aspects de la communication entre cellules réalisée
notamment à l'aide des médiateurs. De la même
façon que pour le système nerveux, on peut donc dire
:
le système immunitaire c'est
le système de contrôle et d'intégration des
populations cellulaires de l'organisme
Remarque:
cette conception n'a rien d'original, j'en suis conscient : ce n'est
qu'une formulation d'idées que l'on retrouve dans de nombreux
supports pédagogiques. Je voudrais citer par exemple la
cassette vidéo de chez Nathan: Sciences de la Vie d'Eric
Périlleux : Le système immunitaire et la
communication entre cellules. Ce film n'est pas un modèle
de pédagogie à mon avis, loin s'en faut, mais la notion
que je présente ici y est sous-jacente et il y a de nombreuses
séquences filmées (longues) tout à fait
exploitables dans le sens que je propose.
retour accueil
résumé, formulation
par cycle