|
(2n = phase diploïde (2n=46 chromosomes chez l'homme), n = phase haploïde(n=23)) |
 |
I. Unicité génétique des individus et polymorphisme des espèces
Le texte en bleu correspond à une
refonte des éléments du programme dans un souci de
cohérence avec la conception de la vie présentée
dans le cours de seconde.
Les chapitres en rouge sont ceux qui ont
été traités en cours
les parties barrées correspondent aux
aménagements de programme du 29/10
1.1 Le génome comprend
l'ensemble des gènes
Il est indispensable de réviser la notion d'"expression
de l'information génétique" acquise en première
S (livre TS pp 8 à 14). Pour vous aider, voici un petit
vocabulaire:
|
|
portion d'ADN qui code pour la synthèse d'une chaîne polypeptidique ; un gène peut être exprimé (actif, transcrit et traduit en protéine) ou réprimé (inactif) |
|||||||||||||||||||||||||||||||||||
|
|
forme d'un gène (forme allèlique) ; un gène comportant de nombreux allèles est qualifié de polyallélique |
|||||||||||||||||||||||||||||||||||
|
|
Acide désoxyribonucléique, molécule en double hélice (double brin) formée de nucléotides, présente dans le cytoplasme des cellules procaryotes et dans le noyau et certains organites (chloroplastes, mitochondries) des cellules eucaryotes ; l'ADN peut être dénaturé (séparation des deux brins) par simple chauffage puis renaturé si l'on baisse à nouveau la température |
|||||||||||||||||||||||||||||||||||
|
|
association d'une base azotée (A=adénine,
T=thymine, C=cytosine, G=guanine, U=uracile), d'un sucre (le
désoxyribose ou le ribose) et d'un, deux ou trois
groupement phosphate (PO43-); selon le
sucre on a un désoxyribo- ou un
ribo-nucléotide, selon le nombre de groupements
phosphates on a un nucléotide mono-, di- ou
tri-phosphate
Les nucléotides sont utilisés dans tout le métabolisme énergétique (ATP, GTP) mais interviennent aussi dans les fonctions de relation (AMPcyclique, second messager de nombreuses hormones). Enfin, ils s'assemblent en polymères informationnels (ADN, ARNm) et structuraux (ARNr)... |
|||||||||||||||||||||||||||||||||||
|
|
Acide RiboNucléique, molécule formée d'une chaîne de nucléotides (monobrin) réunis par des liaisons de covalence; des repliements du brins(maintenus par des liaisons hydrogènes entre bases) déterminent la forme et la fonction de l'ARN (on distingue les ARNm (messagers, ARNr (ribosomiaux), ARNt (de transfert) mais certains ARN ont une activité enzymatique : on les qualifie de ribozymes...) |
|||||||||||||||||||||||||||||||||||
|
|
synthèse d'une molécule d'ARN à partir de l'un des brins de l'ADN (brin transcrit) par un complexe enzymatique: l'ARN polymérase |
|||||||||||||||||||||||||||||||||||
|
|
duplication de l'ADN par le complexe enzymatique ADN polymérase en phase S (synthèse) de l'interphase pour l'ADN nucléaire |
|||||||||||||||||||||||||||||||||||
|
|
synthèse d'un polypeptide (chaîne d'aa) à partir d'un ARNm (messager), de ribosomes (associés en polysomes), de nombreuses enzymes et d'énergie (GTP) ; cette synthèse est réalisable in vitro ; |
|||||||||||||||||||||||||||||||||||
|
|
correspondance entre les bases de l'ARN et les acides aminés (aa); à chaque triplets de bases (codon) de l'ARNm correspond un triplet de bases (anti-codon) de l'ARNt qui transporte un aa spécifique ; le code génétique est universel (quelques cas d'exceptions chez les Procaryotes), redondant (plusieurs codons correspondent au même aa) et dégénéré ou ponctué (certains codons ne correspondent à aucun aa (codons stop) et provoquent l'arrêt de l'allongement de la chaîne polypeptidique) |
1.2 Le génotype d'un individu est la combinaison originale de ses allèles
Si la compréhension de cette phrase ci-dessus reste limitée à l'information génétique, elle est vraie. Mais quelle méprise dans des phrases du genre: "l'unicité de l'individu repose sur l'unicité de son génome". Sans même parler de l'homme, il est nécessaire d'élargir à nouveau notre compréhension de l'individualité du vivant, comme les biologistes l'on toujours fait, l'information génétique n'est qu'une composante, même essentielle, de tout ce qui fait qu'un organisme est cet-individu-là et non un autre (voir cours de seconde). Les êtres vivants ne sont pas interchangeables, même une bactérie avec une autre bactérie.
La technique des empreinte génétiques (voir encadré ci-dessous) permet de visualiser de façon spectaculaire la variabilité de séquence de l'ADN entre individus éloignés. Elle permet aussi de faire des comparaisons intéressantes entre individus de même famille. MAIS ATTENTION les fragments d'ADN visualisés ne sont en aucun cas assimilables aux gènes (tout comme les bandes colorées des caryotypes qui ne correspondent qu'à des variations de fixation des colorants utilisés). Ce qui est mis en évidence par une empreinte est la variabilité de la séquence de l'ADN de chacun.
On peut aussi utiliser dans certains cas favorables des sondes moléculaires radioactives (ADN monobrin ou ARN) pour repérer des allèles spécifiques (programme de SVT de Spécialité).
La localisation de nombreux gènes est connue chez l'homme (voir carte par exemple du chromosome 11 de l'homme p 20). Le génome humain est en cours de séquençage. Je vous conseille d'aller voir par exemple le site du Généthon (http://exogene.genethon.fr/)
|
Une empreinte génétique est le résultat d'une technique de génie génétique. L'ADN est dénaturé (par chauffage) puis coupé en une multitude de fragments (par une enzyme dite de restriction). Une électrophorèse sépare ensuite ces fragments en fonction de leur taille (voir p395). On met ensuite l'électrophorétogramme (résultat de l'électrophorèse) en présence d'une sonde radioactive multi-locus (fragment d'ADN monobrin rendu radioactif et susceptible de se lier par liaison hydrogène à une séquence présente en un très grand nombre d'exemplaires dans l'ADN de chaque personne). Après rinçage, la sonde les fragments d'ADN ayant fixé la sonde sont repérés par autoradiographie (voir p 392). La taille des fragments d'ADN révélés par autoradiographie dépend donc à la fois de la présence d'un site de coupure par l'enzyme de restriction et d'un site de fixation de la sonde (site d'hybridation). Chaque électrophorétogramme est remarquablement spécifique d'un individu (certains estiment la probabilité d'avoir deux empreintes génétiques identiques réalisées avec la même sonde pour deux individus à 3.10-11). Cette technique est utilisée en criminologie ou en recherche de paternité. Lorsque l'on ne dispose pas d'une grande quantité d'ADN il est alors nécessaire d'utiliser la PCR (polymérisation en chaîne) pour multiplier un grand nombre de fois l'ADN récolté. Cette technique est cependant difficile à réaliser et demande d'énorme précautions pour ne pas amplifier de l'ADN parasite... |
La notion de génotype et de
phénotype ont beaucoup évolué et je
souhaiterais être le seul pour qui il était
définitivement établi que tout phénotype
était codé par le génotype. Deux petits exemples
pris dans "Exercices de génétique avec solutions", J-L.
Rossignol, J. Bernard et F. Quétier, Masson,
1980: Problème n°1: Question: Une solution (discussion !) du problème
n°1: Problème n°2: Question: Une solution au problème n°2:
On possède deux lignées d'Euglènes
(Protiste (donc unicellulaire eucaryote) avec une paroi
pectocellulosique, des mitochondries et souvent des
chloroplastes). La lignée A se multiplie en culture
(mitoses assez atypiques, notamment sans disparition de
l'enveloppe nucléaire d'après ce que j'en
sais) et donne des individus qui verdissent à la
lumière (on observe de nombreux chloroplastes dans
leur cytoplasme) et s'étiolent à
l'obscurité (les chloroplastes disparaissent, ils
deviennent franchement hétérotrophes
vis-à-vis du carbone). La lignée B se
multiplie en culture et ne donne que des euglènes
jaunes qui restent hétérotrophes et ne
verdissent jamais, même à la
lumière.
Que pouvez-vous dire sur le génotype et le
phénotype de ces deux lignées ?
Si l'on utilise la terminologie actuelle de
phénotype (tout caractère visible,
quelle qu'en soit l'échelle) et de
génotype (composition allèlique d'un
individu), il est évident que, si l'on
considère que chaque lignée possède un
génotype fixe (car transmissible par mitose),
ce génotype peut coder pour deux phénotypes
différents (lignée A) et inversement,
deux phénotypes (A étiolé et B)
identiques peuvent résulter de deux
génotypes différents.
Le déterminisme de
l'autotrophie/hétérotrophie, qui peut
être caractérisée par la présence
ou l'absence de chloroplastes mais qui n'est bien sûr
pas uniquement limité à cela, est
transmissible par reproduction asexuée (mitoses). Il
est gênant de parler de phénotype.
Personnellement, je préférerai que l'on
réserve le terme de phénotype à des
caractères dont la liaison avec le génotype
est reconnue. On pourrait parler de cytotype (très
laid) ou de morphotype (très galvaudé
car utilisé dans des sens différents).
Des expériences sont réalisées chez
Escherichia coli. Cette bactérie (souche sauvage) est
capable d'utiliser le glucose comme substrat respiratoire
mais aussi le lactose. On connaît une enzyme: la
ß-galactosidase qu'elle synthétise et
excrète dans le milieu. La présence ou
l'absence de cette enzyme peut être
révélée aisément de façon
expérimentale dans le milieu de culture.
On cultive 4 lignées A, B, C et D, qui ont un
comportement différent en fonction du substrat et de
la température du milieu de culture; voici le
résultat des cultures QUI SONT REPRODUCTIBLES SUR DE
NOMBREUSES GÉNÉRATIONS:
+ indique que la souche pousse, - qu'elle ne se
développe pas
ß+ indique la présence de la
ß-galactosidase dans le milieu et ß- son absence
lorsque celle-ci a été testée
Que peut-on en déduire sur les phénotypes et
les génotypes de ces souches ?
Une fois encore on ne sait rien sur les génotypes
à partir du seul phénotype sauf dans le cas
où l'on teste effectivement la présence de la
ß-galactosidase dans le milieu. Il faut séparer
deux caractères essentiels: d'une part la
capacité de la souche à se développer
sur milieu lactosé à telle ou telle
température (+/-), d'autre part la capacité de
la souche à synthétiser et excrèter la
ß-galactosidase dans le milieu (ß+/ß-).
Donc seules les souches D, A et B ont avec certitude le
gène de la ß-galactosidase car ce sont les
seules pour lesquelles la présence dans l'enzyme dans
le milieu a été testée. C n'a donc
peut-être pas le gène, en tout cas il n'est pas
exprimé. Quand à l'expression du gène
de A, B et D en fonction du substrat (présence de
glucose ou non), et de la température, elle est donc
différente pour chacune des souches. C'est une preuve
évidente du contrôle de l'expression du
génome. Toutes les hypothèses sont permises
notamment celles concernant un contrôle cytoplasmique
ou même un contrôle de la population
cellulaire... bien évidemment il peut aussi s'agir
d'un contrôle génétique mais ce n'est
pas du tout la seule hypothèse possible.
1.3 Les mutations produisent de nouveaux allèles
Une mutation est une modification de la séquence de l'ADN. On distingue les mutations ponctuelles (affectant directement un seul codon), seules envisagées ici, et les mutations d'ordre supérieur qui modifient en bloc une grande partie du génome (réparation "excessive" du génome après cassure, échange de chromatides, perte d'ADN...). On distingue classiquement parmi les mutations ponctuelles (voir tableau p17): les mutations par substitution (substitution d'une base par une autre) ou par inversion (changement dans l'ordre des bases d'un codon), par délétion (suppression d'une base) ou par addition ou insertion (ajout d'une base). La fréquence d'apparition spontanée de mutations dans une culture bactérienne est de l'ordre de 10-6. On peut augmenter celle-ci par des agents mutagènes (U.V., produits chimiques variés).
Chez les procaryotes l'apparition de nouveaux caractères (résistance à un antibiotique par exemple) peut directement être mis en relation avec une mutation ponctuelle. On peut donc supposer que la mutation est à l'origine de l'apparition d'un nouveau caractère ou allèle. En fait la mutation provoque la plupart du temps une perte de l'expression d'un gène. Il existe par contre une certaine variabilité génétique propre de nombreuses espèces bactériennes qui, par des mécanismes de recombinaison génétique (échanges de portion d'ADN), entretiennent une grande diversité des produits de l'expression de leur génome. Il ne s'agit pas dans ce cas de mutations mais de la variabilité propre des espèces.
Chez les eucaryotes on a pu mettre en évidence dans certains cas très précis une relation directe entre la synthèse d'une protéine anormale et la présence d'un allèle modifié. C'est par exemple le cas de la drépanocytose où anémie falciforme. La forme des hématies en faucilles et les problèmes respiratoires dus au mauvais fonctionnement à basse pression de dioxygène peuvent être imputés à la présence d'une hémoglobine anormale (HbS) qui présente une substitution d'un nucléotide au codon n°6. L'origine de cette maladie est donc clairement établie comme étant une modification de la séquence d'un gène codant pour l'hémoglobine. Il s'agit donc d'une mutation qui remonte certainement à des temps très anciens de l'humanité.
1.4 Les mutations sont parfois visibles dans le phénotype
On distingue 2 types de mutations ponctuelles:
- les mutations non-sens: le codon muté est un codon
stop qui provoque l'arrêt prématuré de
l'allongement de la chaîne polypeptidique; la protéine
synthétisée est alors généralement non
fonctionnelle
- les mutations faux-sens: qui vont du changement d'un seul aa
au décalage complet du cadre de lecture et donc à la
synthèse d'une chaîne d'aa qui peut être fort
différente.
Une mutation , du fait de la redondance du code génétique, peut être neutre ou silencieuse et ne pas provoquer de changement dans la chaîne polypeptidique codée par le gène muté. Elle peut aussi être supprimée par l'intervention d'ARNt (dits suppresseurs) qui peuvent se lier à plusieurs aa différents.
1.5 Les mutations sont parfois transmises à une autre génération
On imagine qu'il y a des mutations somatiques (qui touchent les cellules du soma, c'est-à-dire les cellules non sexuelles) et des mutations germinales (qui touchent les cellules sexuelles ou germen). Seules les mutations germinales sont susceptibles d'être transmises à une autre génération. Une mutation peut ensuite, si elle est transmise à l'œuf, être exprimée dans toutes les cellules de l'individu.
Pour qu'une mutation soit exprimée chez un diploïde, elle devrait en théorie être présente à la fois dans les deux gamètes ayant fusionné lors de la fécondation, ce qui est fort improbable et oblige toute mutation à intervenir en même temps chez deux individus.
Il devient donc de plus en plus évident que les mutations, découvertes chez les procaryotes puis chez les eucaryotes ont permis d'innombrables progrès notamment en médecine par la découverte de maladies génétiques (voir page sur une histoire de la génétique). Mais, d'une part, toutes les maladies ne sont pas génétiques, d'autre part, les mutations ne sont pas les seules modifications héréditaires, chez les eucaryotes elles perdent franchement de leur importance étant donné la complexité de l'organisme. Les anomalies de développement se référant à des dérèglements touchant l'information cytoplasmique et surtout l'information extracellulaire, deviennent majoritaires (Un livre à lire: Rosine Chandebois: le gène et la forme" voir la bibliographie).
Une mutation, pour être transmise, ne doit bien sûr pas être létale (provoquer la mort de la cellule mutée).
1.6 Les mutations sont responsables du polymorphisme des populations
La théorie actuelle propose que les mutations soient à l'origine de la variabilité des individus dans l'espace et dans le temps. Les mutations non létales, transmises de générations en générations, s'accumuleraient au sein des populations. Quelques essais d'expériences sur des populations de drosophile (petite mouche du vinaigre Drosophila melanogaster) ont été conduites en ce sens. Dans les populations humaines l'application de cette théorie permet d'interpréter la distribution de certains allèles (par exemple les maladies sanguines de type thalassémies sur le pourtour de la thalassa ou Méditerranée) comme liés à des mouvements historiques migratoires des populations.
Cette théorie est
dépassée, elle repose sur une conception erronée
de l'individu, ici l'homme, qui résulterait de l'expression de
ses gènes et donc uniquement de son information
génétique. Il a toujours été
enseigné par les biologistes qu'un individu possédait
non seulement une information génétique, mais aussi une
information cellulaire, cytoplasmique, et enfin une information
extracellulaire, lui venant du milieu extérieur. Limiter
l'information à l'information génétique est une
réduction erronée. Pour quelques détails voir le
cours de seconde.
La répartition des allèles au sein des populations et
l'étude des mutations qui leur ont donné naissance
reste cependant tout à fait légitime tant que l'on ne
considère pas qu'un individu est le résultat de
l'expression de ses allèles, ce qui est rabaisser la personne
humaine à bien peu de choses.
Il est donc clair qu'un individu, même du point de vue
strictement biologique, n'est pas le résultat de la
combinaison de ses allèles. Son génome est
manipulé par ses cellules et il forme un être vivant
autrement plus complexe qu'une simple sommation d'expressions de
gènes. Comparer des individus à l'aide de
séquences d'ADN est une méthode dangereuse dans le sens
où elle ne tient pas compte de la globalité de
l'être vivant.
2. Mécanismes fondamentaux de la reproduction sexuée et du développement
En ce qui concerne le travail de reproduction il faut souligner l'importance des mécanismes cytologiques de la division cellulaire. Vous devez bien comprendre que la mitose ou la méiose ne sont pas des mécanismes de répartition du matériel génétique mais d'authentiques mécanismes de DIVISION CELLULAIRE qui impliquent donc non seulement les chromosomes mais tous les organites cytoplasmiques (division préalable des mitochondries, du REG, du Golgi, des centrioles, ..; répartition égale ou non des ribosomes etc.).
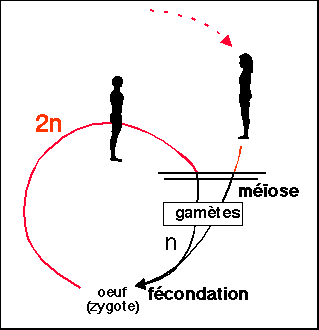
2.1 Le cycle de développement des organismes fait alterner phase haploïde et phase diploïde
Il est caractéristique que l'on présente comme élément essentiel du cycle de développement d'un individu uniquement sa garniture chromosomique. Les biologistes ont toujours parlé de génération, terme auquel il est préférable de revenir. Le cycle de l'homme ne présente qu'une seule génération alors que le cycle des champignons en comporte 2 ou 3. Par souci de simplification, on risque de passer à côté de l'essentiel.
La phase haploïde résulte d'une méiose, la phase diploïde succède à une fécondation. Les cycles haplophasiques sont à haplophase (phase haploïde) dominante, ou phase diploïde réduite au zygote (cellule œuf). Les cycles diplophasiques sont à diplophase (phase diploïde) dominante, ou phase haploïde réduite aux gamètes.
|
(2n = phase diploïde (2n=46 chromosomes chez l'homme), n = phase haploïde(n=23)) |
|
|
2.2 La méiose assure le passage de la phase diploïde à la phase haploïde
Ce titre, qui est de moi, je m'en excuse, et qui est dans l'air du temps (car je sais que c'est la seule chose qui va être demandée à mes élèves au bac), est fondamentalement réducteur. Lors de la méiose les deux divisions successives sont accompagnées de maturations très originales qui seront étudiées lors de l'étude de l'ovogénèse et de la spermatogenèse (dont les mécanismes cytologiques et pour partie physiologiques ont été retirés du programme). Il n'est pas indifférent que ces phénomènes de gamétogénèse se produisent au sein des organes sexuels sous le contrôle d'hormones sexuelles. L'aspect chromosomique de la méiose n'est qu'un aspect très limité des mécanismes de formation des gamètes intervenant dans la reproduction sexuée.
La méiose typique comporte deux divisions successives (2 cytodiérèses (séparation du cytoplasme) donnant donc lieu à 4 cellules filles) dont les étapes chromosomiques (caryodiérèses ou séparations du noyau) sont séparées arbitrairement en 8 phases (prophase I, métaphase I, anaphase I, télophase I, prophase II, métaphase II, anaphase II et télophase II) (voir p31). On retiendra 3 étapes chromosomiques essentielles:
|
|
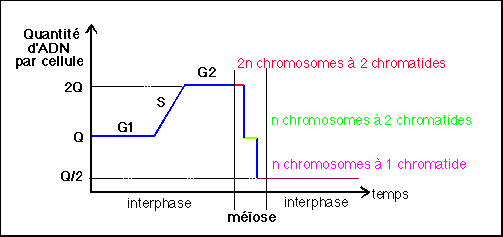 |
Chez l'homme, la méiose est typique et dure environ 24 h et se déroule lors de la spermatogenèse. A partir d'un spermatocyte I on obtient 4 spermatides. Chez la femme, la méiose est atypique, elle commence avant la naissance et ne se termine éventuellement qu'après la fécondation. Lors de l'ovogenèse, l'ovocyte I (bloqué en prophase de première division de méiose depuis la vie embryonnaire) donne naissance (12h avant l'ovulation) à un ovocyte II ou ovocyte mâture et à un globule polaire (à la suite d'une cytodiérèse inégale). L'ovocyte II se bloque immédiatement en métaphase de deuxième division et ne donne naissance à un ovule et à un deuxième globule polaire qu'après l'éventuelle pénétration du noyau d'un spermatozoïde lors de la fécondation.
2.3 La fécondation restaure la diploïde à partir des gamètes haploïdes
Même remarque que pour le titre précédent. La fécondation est d'une part la fusion de deux cellules et donc non seulement la réunion du matériel génétique mais aussi de certains éléments cytoplasmiques (l'inégalité de participation entre l'ovocyte II et le spermatozoïde sont des éléments essentiels) et d'autre part, ce qui est loin d'être négligeable, un événement qui a lieu dans un lieu déterminé (fécondation interne ou externe) avec tous les contrôles que cela suppose, tant sur le déroulement de la fécondation elle-même que sur le devenir de l'oeuf.
Lors de la fécondation chez l'homme qui a lieu dans les trompes, le spermatozoïde doit dissocier les cellules folliculaires grâce à des enzymes qu'il sécrète. Il pénètre ensuite à travers la membrane pellucide grâce à la libération des enzymes contenus dans sa vésicule acrosomiale (acrosome) présente à la tête du spermatozoïde. La tête du spermatozoïde s'accole latéralement à l'ovocyte II, leurs membranes fusionnent (il y a cytogamie ou fusion des cytoplasmes MAIS il semble que seul le noyau du spermatozoïde pénètre dans le cytoplasme de l'ovocyte, ceci est important car, par exemple, on suppose alors que l'ensemble des mitochondries sont d'origine maternelle...). L'ovocyte fécondé se débloque alors et continue sa méiose (anaphase II puis télophase, expulsion du deuxième globule polaire), excrète vers l'extérieur le contenu de petites vésicules qui transforment la membrane pellucide en membrane de fécondation (obstacle essentiel à la polyspermie ou fécondation par plusieurs spermatozoïdes d'un seul ovocyte). La caryogamie (fusion des noyaux que l'on appelle les pronuclei, le pronucleus mâle étant plus gros que le pronucleus femelle) est retardée de quelques heures. La première mitose de l'œuf intervient quelques heures après la fécondation. Le développement se poursuit dans les trompes puis l'utérus après nidation.
3. Brassage génétique assuré par la reproduction sexuée et unicité génétique des individus
Toute cette partie doit être
renommée: il ne s'agit plus ici de faire de l'analyse
génétique formelle (c'est-à-dire une analyse des
résultats de divers croisements aussi bien chez des
haploïdes que des diploïdes: voir pages d'histoire
de la génétique) mais
d'illustrer des notions vues auparavant. Les résultats doivent
être interprétés à l'aide des
mécanismes chromosomiques connus de la méïose ou
supposés du crossing-over. On peut déplorer le fait
qu'il ne soit plus demandé à l'élève que
d'expliquer des résultats à partir de ses connaissances
mais étant donné les sujets posés au
baccalauréat il me semble que l'on n'attend aucune
réflexion critique de la part du candidat, ce qui est fort
dommage.
C'est pour cette raison que je ne vais pas
faire de cours dans cette partie car les règles
élémentaires de l'analyse expérimentale ne sont
pas respectées. Je me contenterai donc de présenter des
exemples (que je ne reporte pas ici) qui sont ceux que l'on peut
demander de restituer sans réflexion au candidat.
|
Étude de la transmission d'un caractère lors du croisement de deux spores chez Sordaria Et si le crossing-over n'existait pas ? |
|
On croise une spore noire avec une spore jaune. Nous avons utilisé comme la plupart des lycées les souches du commerce (Sordalab). On notera que les souches commercialisées sont d'une part la souche sauvage à spore noire, d'autre part des souches manipulées pour éviter des autofécondations. On est donc loin d'un modèle naturel. Comme il est beaucoup trop long de faire une analyse statistique des asques obtenues, le TP se résume à une simple observation. Si l'on veut pousser un peu l'analyse on est franchement catastrophé par la difficulté rencontrée par les élèves... on est pourtant dans un cas d'étude d'un seul caractère...et on voudrait nous faire croire que c'est beaucoup plus simple chez l'homme... |
|
On obtient 6 types d'asques avec les pourcentages habituels. Par exemple (N: spore noire, J: spore jaune) sur 692 asques obtenues: 575 possèdent 2 fois 4 spores de même couleur alignées qui se suivent avec 280 (JJJJNNNN) et 295 (NNNNJJJJ) et 117 asques plus complexes: 26 (JJNNJJNN), 29 (JJNNNNJJ), 32 (NNJJNNJJ) et 30 (NNJJJJNN). |
|
But du jeu: trouver d'autres mécanismes que le crossing-over pour expliquer ces résultats; en fait il s'agit d'essayer de se retrouver dans les conditions de l'expérimentateur qui a observé pour la première fois ce croisement |
|
Une solution: Explication:
|
|
Ce jeu est à mon avis un
excellent exercice. Le risque d'embrouiller l'esprit de
l'élève me semble compensé par la
stimulation qu'il reçoit avec une forte tendance
à éviter les idées reçues. |
|
Ceci n'est donc pas qu'un jeu. Mais il est évident que si l'on étudie la transmission de deux caractères chez des champignons haploïdes, les mécanismes seront différents (je propose une autre interprétation avec trois gènes dans la partie sur la reproduction sexuée). Mon propos n'est pas de juger ni de réinterpréter ce qui a certainement été fait et très bien fait par d'excellents expérimentateurs mais de stimuler l'esprit de mes élèves. |
3.1 Le brassage intrachromosomique est réalisé par crossing-over en prophase de la première division de méiose
3.2 Un brassage interchromosomique est réalisé à l'anaphase de la première division de méiose
3.3 Un autre brassage interchromosomique est réalisé lors de la fécondation
4. Diversité génétique des
populations
4.1 Tous les allèles n'ont pas la même
fréquence dans les populations
4.2 Le milieu exerce une pression sélective sur les
allèles
4.3 Le polymorphisme allèlique des populations est
le résultat de croisements dus aux mouvements des
populations
4. Prévisions en génétique humaine
Du point de vue biologique, la transmission de la vie ne se
résume pas à des facteurs génétiques
comme nous l'avons montré ci-dessus.
Prévoir c'est rechercher les causes
connues. Il faut donc connaître avec certitude une cause
génétique (recherche des causalités d'une
défiscience, d'une maladie), puis la rechercher chez un
individu (dépistage). On peut comprendre que ces deux aspects
de la prédiction soient du ressort du scientifique.
Mais la prévision ne sert à rien en elle-même,
elle doit déboucher sur l'action. Une information sur un
risque génétique fournie à un patient en dehors
du cadre médical me semble détournée de son but.
L'information prédictive doit donc déboucher sur un
acte thérapeutique. Elle doit donc être le fait du
médecin. Ce n'est plus le rôle du biologiste.
Classiquement on distingue la médecine curative (qui
soigne les effets) et la médecine préventive
(qui soigne les causes). Mais la médecine garde toujours le
même but: soigner. Il me semble, mais peut-être ai-je
tort, que l'on confond un peu parfois la notion de médecine
préventive et la notion de médecine
prédictive. La médecin n'est pas devin, ni
mathématicien probabiliste, ni scientifique (au sens
d'exprérimentateur): il applique son art. Ses connaissances
scientifiques, étayées par sa pratique, lui permettent
d'être efficace. Comment peut-on soigner une maladie que le
patient n'a pas encore mais pour laquelle il présente un
risque. Un conseil d'hygiène de vie est approprié et
suffisant, le reste relève de la liberté individuelle.
La société n'a pas le droit d'utiliser les
médecins ou les scientifiques pour faire un dépistage
systématique d'un risque (je souligne bien le fait que je en
parle pas d'une anomalie mais d'un risque) même avec l'accord
des personnes car il y a toujours une certaine coercition à
proposer une analyse gratuite. Si l'on ne peut pas soigner un risque,
tout simplement parcequ'il n'y a pas de maladie à soigner, je
crains que ce que l'on ne cherche en réalité soit de
voir disparaître les inégalités
génétiques: ce n'est plus du soin, c'est de
l'eugénisme. Je sais que le mot est fort mais il je
crois qu'il faut être clair et qu'il ne peut pas y avoir
d'attitude neutre. Le dépistage systématique du risque
est pour moi une atteinte grave à la liberté
individuelle alors qu'elle se présente comme
libératrice. Il est compréhensible qu'une
société cherche à minimiser ses
coûts de santé (il est moins cher de proposer un
dépistage génétique puis des avortements (voir
plus loin) plutôt que de soigner des malades sur de longues
années) mais n'est pas acceptable que cela se fasse au
détriment du respect des personnes. C'est ici qu'il y a un
problème majeur d'éthique à soulever.
Le danger me semble beaucoup plus important que l'on veut nous le
faire croire, mais je me trompe peut-être. Rechercher un
"défaut" génétique, c'est-à-dire par
exemple un allèle muté qui, à l'état
homozygote, est cause d'une maladie grave, n'est pas un acte
bénin. Que va-t-on proposer au vu de résultats positifs
? Ne pas procréer ? Choisir un conjoint en fonction de cet
allèle ? Dans ce cas tout le monde se recrie (voir Bordas
cours par exemple, p 85 &3 à propos des
thalassémies en Italie.... je me demande quelle est la
fiabilité de ces informations. A mon avis elles sont tout
simplement fausses). C'est encore pire si l'on continue notre
lecture: "Cependant, le nombre d'enfants thalassémiques
diminue rapidement du fait des progrès du diagnostic
prénatal qui permet de détecter les foetus
homozygotes". Il manque la fin de la phrase que l'on imagine
aisément : ...et proposer alors un avortement
thérapeutique.... A moins qu'il existe un moyen de soigner
ces enfants précocement ou même in utero mais je crains
que cela ne soit pas le cas. Le fait de tuer un foetus n'est pas un
acte médical. Le foetus est vivant, il n'est pas encore
malade, il se développe. On le tue par peur, par refus de ce
qu'il pourra souffrir ou être plus tard. C'est une lourde
responsabilité, certainement pas celle d'un médecin. En
tout cas je ne vois pas comment, en tant que professeur de biologie
(j'ai donc la vie pour profession), je peux présenter de
telles attitudes comme normales, courantes, morales (en accord avec
la fin de l'homme) à mes élèves.
Un article dans la revue de la
MGEN, Valeurs mutualistes (n°211, mai 2001, "Gare
à la tentation d'un eugénisme
doux"), un entretien avec Alain Etchegoyen (ancien
membre du Conseil national d'éthique), me
paraît tout à fait aller dans le même
sens: en voici quelques extraits
« EH : Quels doivent être selon vous les
objectifs d'une loi sur la bioéthique ?
A.E. : La loi doit avant tout interdire certaines offres.
Car à partir du moment où l'offre existe, la
demande existe forcément et peut se faire pressante.
Que se passerait-il, par exemple, si à l'occasion
d'un diagnostic pré-implantatoire, les
médecins disaient aux parents :
«Voilà, nous pouvons faire en sorte que votre
futur enfant soit exempt de telle ou telle maladie
très grave dont vous êtes porteurs dans la
famille, et si vous voulez, nous pouvons aussi éviter
qu'il ait un bec de lièvre ou un doigt plus court que
l'autre». Il est très difficile de
résister à de telles offres, d'autant plus
qu'une économie de marché accroît encore
la pression. Si vous avez par exemple la possibilité
technique d'éviter que votre futur enfant soit
asthmatique, que se passera-t-il si vous la refusez ? Une
compagnie d'assurance ne pourra-t-elle pas tirer argument de
votre attitude pour refuser de prendre en charge les soins
de l'enfant ? Une décision historique a
été prise par Tony Blair cet été
: il a autorisé les compagnies d'assurances à
demander les analyses génétiques de leurs
assurés lorsque ceux-ci ont des maladies
génétiques dans leur famille. En France, il
n'en est pas question pour le moment. Il existe un
moratoire, mais la question n'est pas définitivement
réglée.
KM. : Vous dites qu'il ne faut pas faire porter aux
individus des responsabilités
«artificielles».
A.E : Dès lors qu'une offre est terrible et
culpabilisante, dès lors que la pression du
marché pèse de tout son poids, les individus
se retrouvent effectivement devant des choix auxquels ils ne
devraient pas être confrontés. Il ne faut pas
non plus sous-estimer la compétition entre les
médecins et entre les hôpitaux. Parler de
"corps médical" est une illusion, car il y a en fait
très peu d'union et beaucoup de rivalités !
Cette compétition contribue elle aussi à nous
faire entrer dans un cycle où l'on ne maîtrise
plus rien. Une innovation, quelle qu'elle soit,
débordera toujours de ces objectifs initiaux à
partir du moment où elle entre sur le marché.
On l'a vu avec l'hormone de croissance : les circuits les
plus fous ont été mis en place afin de
poursuivre l'approvisionnement. On est allé
cherché des hypophyses jusqu'au fin fond de la
Roumanie ! Je me souviens d'une publicité pour un
institut médical qui disait : « Mieux vaut
une consultation de plus que dix centimètres de
moins.» Après les cas de nanisme, l'offre
médicale s'était élargie à toute
les personnes de petite taille, comme si c'était
inconcevable de mesurer moins d'un mètre cinquante.
Dans notre société, la normalité
devient obsédante.
VM. Selon vous, la démocratie n'est pas un rempart
contre les dérives les plus extrêmes ?
A~E. : Il y a un événement qui se produit
très souvent lorsque je donne une conférence
sur la bioéthique : à la fin, une personne
aveugle, ou atteinte d'un handicap quel qu'il soit, vient me
trouver en disant : «Finalement, ils voudraient que
je n'existe pas.» Et je pense que nous vivons
effectivement dans une société très
normalisatrice, où la tentation d'un eugénisme
«doux» est très présente. Certains
scientifiques récusent l'utilisation du terme
«eugénisme» , qu'ils relient uniquement aux
régimes totalitaires. Et pourtant, à mon sens,
l'eugénisme démocratique est le pire qui soit.
Justement parce qu'il se présente sous forme indolore
et presque invisible. Un acte comme le diagnostic
pré-implantatoire est très
médicalisé, abstrait, mais il produit bel et
bien un eugénisme d'un type nouveau, qui n'est pas
plus acceptable que de supprimer des enfants à la
naissance.
Ces considérations sont
peut-être inutiles dans ce cours car ce qui est effectivement
demandé à un élève de terminale est
beaucoup plus innocent mais à mon avis peu valorisant
intellectuellement : on lui demande simplement d'appliquer ses
connaissances sur la méïose à l'analyse des arbres
généalogiques. Là encore, le titre du programme,
beaucoup trop ambitieux, ne reflète pas le niveau des sujets
que l'on peut poser. Pourquoi ne pas parler d'analyse des arbres
généalogiques ?
Il y a une autre question que je me pose. J'espère qu'elle est
complétement idiote mais je n'en suis pas si sûr.
Certains sujets du baccalauréat présentent à la
fois des arbres généalogiques classiques et des
résultats modernes issus des techniques d'hybridation de
sondes radioactives. Je me demande parfois si ces deux types de
résultats ont bien été obtenus pour la
même maladie et pour les individus indiqués. On a
parfois l'impression que ces exercices sont théoriques,
un peu comme des exercices de mathématiques... J'espère
rêver et que personne n'aurait osé falsifier des
résultats en les proposant à l'analyse sans qu'ils
soient réels, c'est-à-dire expérimentaux... mais
je me demande parfois..... Si l'intention est louable le
procédé resterait inacceptable, même à des
fins pédagogiques, car il remet en cause le fondement
même de notre science expérimentale.
4.1 Une
génétique de l'homme
L'homme présente des caractéristiques
spécifiques au niveau biologique: un cycle de
développement long (9 mois de développement
embryonnaire et fœtal intra-utérin, une dizaine
d'années avant la maturité pubertaire), une
fécondité faible (en regard d'autres organismes vivants
capables par reproduction sexuée ou asexuée de donner
naissance à des milliers d'individus; en tout cas il n'y a pas
la possibilité d'effectuer des statistiques sur une seule
descendance humaine), une formule chromosomique complexe (23 paires
de chromosomes)...
L'homme présente aussi des spécificités qui,
pour ne pas être du domaine biologique, n'en sont pas moins
incontournables: le respect de la liberté humaine fait que
l'on ne peut pas imaginer des croisements expérimentaux (ne
serait-ce qu'à l'état de suggestion...). De même,
toute prévision, recherche ou information communiquée
à l'homme implique sa personne et souvent un couple, le
scientifique doit alors laisser la place au médecin ou au
psychologue ou à d'autres spécialistes de la nature
humaine qui tiendra compte de cette dimension éthique.
|
Elles sont quatre, assises en demi-cercle, devant mon
bureau, cheveux, yeux, stature de filles Viking, comme leur
mère (comme leur père aussi je l'apprendrai
plus tard), debout à l'écart. Toutes sauf une,
Nathalie la dernière, brune aux prunelles sombres. Et
la mère me raconte cette histoire incroyable :
lorsque Nathalie est née il y a quinze ans, un
visiteur ami lui a dit à la clinique, mi-rieur,
mi-sérieux : «cette fille-là ne peut
être de ton mari ... c'est dans tous les livres de
génétique». Telle la calomnie
chantée par don Basile, cette plaisanterie
discourtoise et maladroite s'est enflée
démesurément. D'autres amis du couple l'ont
reprise, les soirs de rencontre. Le mari blond aux yeux
clairs a douté. Nathalie a demandé un jour :
«Quel est mon vrai père, puisque je ne suis pas
la fille de papa ? ». |
4.2 L'étude d'un arbre généalogique permet parfois de prévoir le phénotype des descendants: des cas d'hérédité mendélienne
4.2.1 Transmission des groupes sanguins du système ABO (groupes de Landsteiner)p76, présentation formelle; p88 - interprétation moléculaire
4.2.2 Hérédité liée au sexe: daltonisme et hémophilie, myopathie de Duchenne
p 80-81
4.2.3 Analyse simplifiée des arbres généalogiques
Il est préférable de parler de transmission d'un caractère, même si presque tout le monde accepte que l'on parle de transmission d'un allèle, forme d'un gène associé à un caractère.
Un caractère transmis héréditairement (c'est-à-dire présent dans un arbre généalogique chez de nombreux individus et donc supposé être porté par les chromosomes au niveau d'un ou plusieurs gènes), est dit récessif lorsqu'il apparaît chez un enfant sans apparaître chez au moins l'un de ses parents (certains parlent de "saut de génération"). Il est dit dominant dans le cas contraire mais il n'y a jamais de certitude de dominance, uniquement des indications (nombreux individus présentant ce caractère dans l'arbre alors qu'il est rare dans la population, un enfant possédant le caractère supposé dominant a toujours au moins l'un de ses parents présentant ce caractère...).
Une fois établie la dominance ou récessivité du caractère considéré et uniquement alors, on regarde la transmission du caractère relativement au sexe. Là encore les cas de certitude sont extrêmement rares (et attention, il ne s'agit pas d'une certitude totale, elle n'est posée que dans le cadre de l'interprétation héréditaire mendélienne qui reste une hypothèse).
On prendra soin d'utiliser un vocabulaire précis montrant bien que la localisation autosomale ou gonosomale n'est qu'une interprétation chromosomique d'une transmission héréditaire. Lorsque l'on avance cette hypothèse on n'affirme pas qu'il y a un lien direct, absolu, physique entre les chromosomes en question et le caractère héréditaire.
La dernière étape consiste à formaliser, certainement de façon abusive (simplificatrice), ces hypothèses. A chaque caractère on fait correspondre un gène qui peut se présenter sous plusieurs formes alléliques. La localisation du gène expliquant la transmission héréditaire du caractère. Dans certains cas il faudra avancer l'hypothèse de trois allèles ou de plusieurs gènes (groupes sanguins de Landsteiner par exemple).
L'analyse statistique de la descendance de l'homme est réduite à l'énoncé des proportions théoriques des gamètes dans le cadre de l'hypothèse chromosomique avancée plus haut (hérédité mendélienne). La fréquence allélique au sein des populations relève d'une toute autre démarche et ne sera pas évoquée ici (cette notion a été retirée du programme à ma plus grande joie).
4.3 De la maladie héréditaire
à la maladie génétique : une analyse qui n'est
pas si simple
La liaison entre le génotype et le phénotype, le
passage de la notion de transmission héréditaire
à la caractérisation d'un gène, ... sont loin
d'être précisément connues et toujours
réductibles à des modèles simples. (voir
Une histoire de la
génétique en cours de rédaction... cette
partie ne peut être abordée sans déboucher sur
une analyse fine des marqueurs génétiques et donc
empiéter sur le programme de spécialité
SVT).
C'est Archibald
Garrod
(1857-?) qui exprime la première fois la possible relation
entre un gène et une enzyme. Il travaille sur une anomalie
métabolique humaine: l'alcaptonurie, qui affecte le
métabolisme de la tyrosine et de la phénylalanine. Il
propose en 1909 dans un article intitulé "Les erreurs
innées du métabolisme", de justifier des
déficiences enzymatiques héréditaires de l'homme
(albinisme, cystinurie, pentosurie...) par des anomalies
génétiques dues à l'inactivation de gènes
codant pour certaines enzymes....
Depuis de nombreuses maladies à transmission
héréditaire que l'on qualifie donc souvent de
maladies génétiques peuvent être
dépistées par des dosages enzymatiques. Un exemple
classique est la maladie de Tay-Sachs qui est indécelable chez
le nouveau-né puis se manifeste vers l'âge de six mois
par des réactions de frayeur excessive au moindre bruit. Se
développent ensuite une cécité progressive, une
paralysie, un retard mental et un accroissement du volume
crânien. L'issue toujours fatale, intervient vers trois ou
quatre ans, est due à une dégénérescence
progressive du système nerveux. C'est en 1969 que O'Brien et
Okada, on prouvé que la maladie pouvait être mise en
relation avec l'absence d'une enzyme: l'hexosaminidase A (Hes.A),
indispensable à la dégradation de certains lipides. Le
taux d'Hes.A est quasiment nul chez les enfants atteints de la
maladie de Tay-Sachs et abaissé de moitié chez les
individus supposés hétérozygotes
vis-à-vis du gène codant pour cette enzyme par rapport
au taux normal observé chez les individus sains n'ayant jamais
eu d'ascendants ni de descendants atteints par cette maladie. Si ce
marqueur enzymatique semble bien corrélé à la
maladie on ne peut pas cependant affirmer que la maladie est due au
seul dysfonctionnement de ce gène. On peut même affirmer
sans grande crainte de se tromper qu'une maladie aussi complexe ne
dépend probablement pas d'un seul gène. Nous avons donc
à notre disposition un marqueur qui peut être
affecté d'un certain coefficient de fiabilité.
Malheureusement il n'existe pas de moyen simple de mesurer cette
fiabilité. Les seules méthodes dont on dispose sont
statistiques. Elles dépendent du recensement médical
des cas et des résultats des dosages enzymatiques
rapportés aux cas déclarés.
De la maladie génétique, on est vite passé à la maladie génique, c'est-à-dire à une maladie causée par un seul gène, ou du moins par un ensemble de gènes connus. Le cas le plus simple (?) est celui des ß-thalassémies dont la plus connue est la drépanocytose pour laquelle il semble y avoir une corrélation totale entre la mutation au niveau du gène de la chaîne ß de l'hémoglobine et le fonctionnement anormal de cette hémoglobine, qui serait à l'origine de tous les symptômes de cette maladie. Mais là encore, il est loin d'être certain que cette mutation est réellement la seule en cause. Si elle est toujours présente elle n'est pas cependant forcément la seule. On peut imaginer des dysfonctionnement au cours de l'embryogenèse, peut-être dus en partie au fonctionnement de ce gène ou à d'autres mécanismes causés par ce gène, peut-être même tout simplement dans l'approvisionnement en oxygène du fœtus par une mère hétérozygote... bref on est toujours dans le domaine des hypothèses et il est dangereux de considérer que l'on connaît LE gène et que donc la maladie est définitivement connue.
Les techniques de dépistage par hybridation de sondes radioactives ou fluorescentes ne changent pas le raisonnement de fond. Quel est le lien entre le gène trouvé et la maladie héréditaire ? On ne peut jamais répondre par autre chose qu'une série d'hypothèses, plus ou moins fiables. Ces hypothèses trouvent leur justification dans des les progrès faits dans la mise au point de thérapies. Ces études sont réservées aux élèves de spécialité.