Cours de
seconde
Biologie II
- Cellule, ADN et unité du vivant
(11 semaines)
Ce cours est une archive
2000-2005, un nouveau cours se trouve sur ces
pages
programme de la classe de seconde,
accueil, cours
- Page annexe: Introduction (les mots
de la vie, l'étude de la vie, rappels
et reformulation des acquis du collège)
- Plan de cette page:
- Annexe:
L'information est parfois un concept flou en biologie et on
pourrait souvent l'abandonner au profit de la forme et des
fonctions (partie plus particulièrement à
l'attention des collègues)
1. Des cellules... à la cellule, unité
du vivant
Un être vivant c'est une
forme
vivante.
Un être vivant c'est d'abord une forme, perçue
habituellement par le sens le plus développé chez
l'homme: la vision. Cette forme peut être décrite
géométriquement. La géométrie est
une branche des mathématiques qui étudie les formes
dans l'espace; c'est-à-dire qu'elle s'intéresse
à l'espace de tout un chacun (à trois dimensions) mais
sans s'intéresser au temps qui forme la quatrième
dimension de notre espace de vie. Il faut donc, en plus de la
géométrie une cinématique qui est la
science des corps en mouvement (des corps qui changent de
coordonnées dans l'espace au cours du temps). Enfin, comme les
formes se déforment avec le temps, il faut une
dynamique qui décrive ces transformations au cours du
temps.
Les biologistes ont l'habitude de décrire les formes en
termes de structures et les dynamiques et mouvements en termes
de fonctions. Mais ces termes recouvrent des notions qui n'ont
cessé de s'appauvrir au fil des siècles (et qui ne
parlent plus aux élèves) au point qu'il apparaît
urgent de les définir le plus précisément
possible avec l'outil intellectuel le plus performant et universel
qui soit (neutre éthiquement de plus): les
mathématiques.
Remarque: la notion de forme, notamment en
biologie, a été particulièrement
développée par un mathématicien
académicien, René Thom, récemment
décédé. Ses outils qui continuent d'être
développés en France pourraient s'avérer
être essentiels. Voir un aperçu dans une
page
annexe.
1.1 . La cellule est une unité de
structure
Parfois il est préférable d'utiliser le terme de
forme au sens courant de volume dans l'espace usuel à 3
dimensions plutôt que de structure mais dans ce cas il
est clair que c'est une forme statique (sans le temps) que l'on
s'efforce de décrire.
1.1.1 - Les formes vivantes les plus simples sont les
unicellulaires mais elles vivent rarement isolées
1.1.1.1 - Les plus petites formes vivantes sont des
bactéries, unicellulaires procaryotes, qui forment des groupes
d'un grand nombre d'individus
|
caractères
|
cellules procaryotes
|
|
taille
|
100 nm de diamètre pour les plus petits
mycoplasmes à 7 µm de diamètre pour
certaines bactéries bleues (Oscillatoria);
Escherichia coli, de taille moyenne mesure 1 à 1,5
µm de largeur sur 2 à 6 µm de
longueur. La plus grande cellule procaryote
connue a été découverte en 1999 par
Heide Schulz et col. (Inst. Max Planck, Brème) et
atteint 0,1 à 0,7 mm de diamètre
(Thiomargarita namibiensis : la perle de soufre :
Pour la Science, 260, juin 99, 26)
|
|
formes
|
très variées dans le détail mais
deux formes simples dominent: les sphères ; et
les bâtonnets plus ou moins allongés
(filaments), incurvés ou en hélice. A
cette forme générale il faut ajouter les
prolongements comme les flagelles qui peuvent être
très nombreux. Enfin les cellules peuvent rester
attachées après leurs divisions et donner des
chaînettes ou des longues files.
|
|
compartimentation*
|
habituellement un seul compartiment interne
délimité par une membrane s'oppose à un
compartiment externe comprenant la paroi; certaines
cellules comme celles des bactéries bleues
possèdent des sous-compartiments internes
délimités par des invaginations de la
membrane
|
|
contenu cytoplasmique
|
pas d'organites délimités par une
membrane donc pas de vésicules donc pas de
transport rapide de matière dans la cellule
|
|
matériel génétique
|
pas de noyau délimité par une
enveloppe (procaryote signifie "sans noyau") et jamais de
chromosomes lors de la division cellulaire
|
|
associations - écologie
|
les cellules procaryotes peuvent survivre isolées
à l'état de spore mais elles sont
habituellement en grand nombre dans un milieu car elles se
multiplient très rapidement. Dans un milieu liquide
elles se déplacent mais on tendance à former
des amas (voile bactérien par exemple) et sur un
milieu solide elles forment toujours des colonies
|
* cette notion est contestable pour les
compartiments internes; elle repose sur l'idée d'une membrane
qui limite structuralement alors qu'il est plus probable que les
membranes internes soit des traces de dynamiques (comme la membrane
externe est le bord (au sens topologique) de la cellule; la
compartimentation existe mais elle est plus dynamique ou
métabolique que structurale.
1.1.1.2 - Les unicellulaires eucaryotes restent de taille modeste
(micro-organismes) mais peuvent former de colonies de grande taille
(quelques dizaines de centimètres)
|
caractères
|
cellules eucaryotes
|
|
taille
|
Les micro-organismes eucaryotes sont des cellules de
quelques micromètres à quelques dizaines de
micromètres dans leur plus petite dimension. Mais
certaines cellules peuvent avoir des prolongements de
plusieurs dizaines de centimètres.
Des cellules peuvent fusionner pour donner
de grandes cellules contenant plusieurs noyaux
(plasmodes).
|
|
formes
|
La variété des formes est nettement plus
importante que pour les cellules procaryotes. On pourrait
dire qu'il y a une "souplesse" beaucoup plus importante de
la cellule dues à des structures du cytoplasme
permettant le mouvement. Des prolongements en filaments ou
en lamelles apparaissent. Une paroi existe souvent
mais l'on a aussi des tests (sorte de coquilles
externes) qui n'empêchent pas la sortie de
prolongements par des pores.
|
|
compartimentation*
|
Un compartiment principal interne
(délimité par une membrane) et un
compartiment externe comprenant souvent une paroi ou
un test. De nombreux sous-compartiments internes
(délimités ou non par des membranes)
permettant une division du travail du vivant (chaque
compartiment ayant une ou des fonctions
spécifiques)
|
|
contenu cytoplasmique
|
nombreux organites délimités par des
membranes simples ou doubles (mitochondries, chloroplastes,
vacuoles...); les transports de matières peuvent se
faire rapidement par des vésicules dans la
cellule
|
|
matériel génétique
|
noyau délimité par une double
membrane ou enveloppe nucléaire (eucaryote signifie
"avec un noyau vrai") et chromosomes apparaissant
lors de la division cellulaire
|
|
associations
|
les cellules eucaryotes forment soit des organismes
libres soit des colonies soit des organismes
pluricellulaires en se regroupant en tissus et en
organes qui se partagent le travail du vivant
|
* même remarque que
précédemment; la compartimentation est plus dynamique
(métabolique) que structurale; l'absence de membrane dans un
espace cytoplasmique n'empêche absolument pas une réelle
compartimentation. Il serait plus exact de dire que l'on ne
connaît pas le rôle des membranes en tant que traces de
dynamiques internes. On sait qu'elles sont le support de nombreuses
protéines qui agissent à l'interface entre le
compartiment supposé et son environnement.
1.1.2 - Les formes vivantes complexes de grande taille sont
composées de cellules de type eucaryote: ce sont les
pluricellulaires
Les cellules eucaryotes des pluricellulaires ne sont pas des
organismes vivants mais des cellules vivantes qui font partie d'un
tout, l'organisme vivant. Elles ne possèdent que partiellement
les fonctions globales (nutrition, relation, reproduction). Ces
cellules sont spécialisées ou
différenciées.
1.1.3 Tous les êtres vivants sont composés de
cellules et uniquement de cellules: c'est la théorie
cellulaire
On doit à T. S. Schwann (1839), un zoologiste, le premier
énoncé de la théorie cellulaire:
« il existe un principe général pour la
production de tous les corps organiques, et que ce principe est la
formation de cellules, aussi bien que les conclusions qu'on peut
tirer de cette proposition, peut être compris sous le terme de
théorie cellulaire». Cet énoncé fait
suite à une correspondance avec botaniste contemporain M. J.
Schleiden que l'histoire a donc associé à Schwann. Avec
nos mots du XXIème siècle cette théorie peut
s'énoncer ainsi:
Tout être vivant est
formé à partir de cellules et uniquement de
cellules.
Cette théorie, rapidement adoptée est
précisée par de nombreux biologistes dont le plus
célèbre est Virchow connu pour son axiome: Omnis
cellula e cellula (1858) : toute cellule est issue d'une
autre cellule. La branche de la biologie qui étudie les
cellules est la cytologie.
Pour un organisme unicellulaire, c'est la cellule unique qui
réalise tout le travail du vivant.
Pour un organisme pluricellulaire, il y a division du travail,
les cellules se différencient (se spécialisent)
et s'organisent en tissus puis en organes pour
réaliser une même fonction, division du travail de
l'organisme.
1.2 La cellule est une
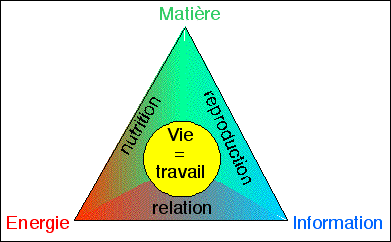
unité de fonctions globales
la vie de la cellule est un travail de relation, de nutrition et
de reproduction
Si le terme de fonction en biologie est familier aux
enseignants il ne l'est pas (plus) aux élèves. Une
fonction en mathématiques c'est une relation (y =
f(x)) représentée par une courbe dans un plan;
l'ordonnée étant la valeur de la fonction (y) et
l'abscisse (x), étant la variable. Parler de "fonction du
vivant" (ou de "fonctionnement") pour désigner la respiration,
la circulation ou une réaction chimique dans une cellule est
une image mathématique utilisée depuis très
longtemps (peut-être même d'abord utilisée en
biologie avant son utilisation en mathématiques
- voir René Thom, Analyse sémantique
d'un mot polysémique : la fonction, , 1993, Séminaires
de la Société de Biologie Théorique,
année 1992-1993, Acta Biotheoretica, 42 ; 1993f5.pdf).
Utiliser le terme fonction suppose que l'on puisse
représenter graphiquement (sous forme d'une fonction
mathématique) le phénomène que l'on
étudie. Cela est parfois possible mais échoue
toujours lorsque l'on s'intéresse aux caractéristiques
mêmes du vivant - ce qui fait qu'un être vivant est
vivant est non une machine-. On nomme ces caractéristiques des
grandes fonctions ou des fonctions globales,
c'est-à-dire des fonctions non locales (ou non
localisables) car elles ne peuvent être situées dans tel
ou tel organe, mais mettent en jeu d'innombrables parties agissant
ensemble. Je propose de parler de
travail. Toute partie de
l'être vivant (molécule, cellule, organe...) participe
à ce travail, c'est sa fonction participative. Cette
fonction participative, biologique, conceptuelle, s'ajoute à
une fonction propre, locale, qui peut être
explorée chimiquement (expérimentalement).
|
La
vie est un travail
|
|
Le travail de
nutrition comprend l'ensemble des
phénomènes de prise ou de rejet de
matière et d'énergie vis-à-vis du
milieu extérieur et qui permettent à
l'organisme de se maintenir en vie. (alimentation,
digestion, respiration, circulation,excrétion ...).
Les autotrophes (du grec "auto" =
"soi-même" et "trophé"= "nourriture")) se
nourrissent seuls sans dépendre d'autres êtres
vivants. Les hétérotrophes ou
plutôt allotrophes* (du grec
"hétéro" = "l'autre, en tant que
différent" et du grec "allo"= "l'autre, en tant qu'un
autre que moi") consomment d'autres organismes vivants ou
morts qui sont les proies.
|
La vie se construit à partir de la matière
qu'elle prend dans le milieu extérieur. L'être
vivant renouvelle ses structures et rejette des
déchets.
La vie coûte de l'énergie.
Certains êtres vivants se nourrissent aux
dépens d'autres.
|
|
Le travail de
reproduction correspond à la
capacité des êtres vivants à se
multiplier. La reproduction peut être asexuée
(solitaire) ou sexuée (entre deux organismes de sexe
opposé).
|
La vie colonise tous les milieux. Elle se propage sans
cesse. La vie se donne.
|
|
Le travail de
relation correspond aux communications entre
l'organisme et son milieu (écosystème) ou
entre les différentes cellules d'un organisme.
|
La vie est sociale. La vie est coordonnée. La vie
est altruiste.
|
|
Le
métabolisme (du
grec métabolé = changement)
désigne
l'ensemble des
mouvements du vivant. Le métabolisme
c'est la dynamique de la vie.
Pour les biochimistes, qui étudient le vivant avec
les outils et les concepts de la chimie, la vie est un
ensemble de réactions chimiques. Le
métabolisme désigne alors les synthèses
ou les dégradations de molécules chimiques qui
ont lieu dans une cellule. Le synthèses forment
l'anabolisme, et les dégradations forment
le catabolisme.
Pour un biologiste il est préférable de relier
le métabolisme aux fonctions globales du vivant et
dire qu'il existe un métabolisme de relation, un
métabolisme de nutrition et un métabolisme de
reproduction.
Remarque philosophique:
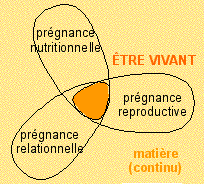
C'est le métabolisme qui donne sa forme au vivant. Ce
qui revient à dire que les dynamiques (de nutrition,
de reproduction et de relation) maintiennent la forme du
vivant (ce sont les prégnances de René Thom
que l'on peut assimiler aux fonctions globales du vivant).
La matière (au sens d'Aristote) est un continu qui
prend forme (ce qui conduit à une saillance dans le
vocabulaire thomien) grâce aux dynamiques qui
l'informent. Pour simplifier on pourrait dire que les
dynamiques correspondent à peu près à
l'énergie. L'interaction
matière-énergie étant qualifiée
d'information (informer = donner une forme). (Voir
compléments sur la page
des 4 causes d'Aristote en
SVT)
|
* allotrophie est parfois employé en
écologie pour désigner un commensalisme.
|
L'activité
cellulaire (le fonctionnement de la cellule) ce sont des
mouvements (des échanges et des
transformations):
|
|
- mouvements
(échanges et transformation) de matière
(pour se nourrir, pour
croître...)
- mouvements (échanges et transformation)
d'information (pour
connaître le milieu extérieur et pour
communiquer avec d'autres cellules ou
organismes)
- mouvements (échanges et transformation)
d'énergie (pour se
maintenir en vie.)
|
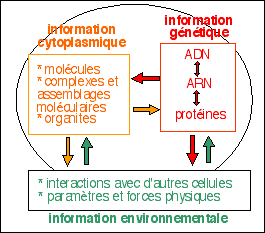
3 informations et non une
seule
|
la vie reste un
mystère
|
|
|
L'information
génétique, contenue dans l'ADN (Acide
DésoxyriboNucléique), conserve la
séquence des ARN (acides ribonucléiques) et
l'ordre dans lequel la cellule doit assembler les aa qui
composent les protéines.
|
L'information génétique est
conservée, dupliquée, manipulée et
exprimée par la cellule. Elle est aussi
modifiée par l'environnement et par le
cytoplasme.
|
|
L'information cytoplasmique
contrôle l'ensemble des activités de la cellule
(métabolisme, mouvement, ...) et conserve la
mémoire cellulaire.
|
L'information cytoplasmique est la personnalité
cellulaire.
|
|
L'information environnementale
est en permanence reçue et envoyée par
la cellule qui réagit à son environnement et
communique avec lui.
|
L'information extérieure permet à la
cellule de coordonner sa propre activité avec celle
des cellules voisines.
|
1.3 Les êtres vivants sont
classés en 5 royaumes* aux limites incertaines:
les procaryotes, les protistes, les mycètes, les plantes
et les animaux
* Linné distinguant classiquement dans le regna tria
naturae, les trois regnum animale, regnum vegetabile et regnum
lapideum (les trois royaumes: des animaux, des
végétaux et des pierres). Le terme latin
regnum a perdu progressivement sa signification
de royaume et a été malheureusement
traduit par règne (voir
http://www.necker.fr/sfbt/baillaud.html);
il est incontestablement plus clair de parler de royaumes
(kingdom en anglais et Reich en allemand) car cela
permet notamment des changements de souveraineté dont nous
laisserons l'initiative aux spécialistes.
|
royaumes*
|
exemples
|
travail de relation
|
travail de nutrition
|
travail de reproduction
|
Procaryotes
(Monères)
|
bactéries du yaourt (Streptococcus,
Lactobacillus)
|
Une bactérie ne vit jamais seule (sauf à
l'état de spore) mais forme une colonie dès
qu'elle est en présence d'eau et de nourriture. Deux
bactéries peuvent se conjuguer et échanger du
matériel génétique. Les
bactéries se déplacent souvent grâce
à des flagelles. Une paroi les protège.
|
Les bactéries ont un métabolisme
très peu élevé et souvent très
original. Si de nombreuses bactéries sont
allotrophes on en trouve aussi des autotrophes
comme les bactéries bleues (Cyanophytes) qui sont
chlorophylliennes. Certaines bactéries utilisent
même des minéraux (comme le soufre) comme
source d'énergie.
|
Les bactéries se divisent incessamment par
scissiparité (une cellule mère s'allonge puis
se sépare en deux).
|
Protistes
(unicellulaires eucaryotes)
|
Paramécie, levure de boulanger (Saccharomyces
cerevisiae)
|
Les protistes vivent presque tous dans l'eau et se
déplacent souvent par des flagelles ou des cils
(comme la paramécie). Certains sont
protégés par une paroi.
|
Certains protistes anciennement rattachés aux
animaux se nourrissent de matière organique
(allotrophe) comme la Paramécie (Protozoaire)
qui capture des bactéries. D'autres utilisent le
carbone de l'air, de l'eau et de l'énergie solaire
pour synthétiser (autotrophie) leur propre
matière organique (anciens végétaux ou
Protophytes) comme l'Entéromorphe qui forme des
colonies filamenteuses (les communes "algues vertes" de nos
côtes).
|
Les cellules peuvent se diviser seules (mécanisme
de la mitose): c'est la reproduction asexuée. Ou se
reproduire à deux: c'est la reproduction
sexuée : après s'être réunies
(fécondation) elles se séparent lors du
mécanisme de la méiose. Le bourgeonnement (par
exemple de la levure) est un mode de reproduction
asexuée.
|
Mycètes
(champignons)
|
Champignon de Paris, moisissure du pain
|
Les champignons vivent sous la forme de filaments
(mycélium). Ils ne se déplacent pas. Ils ont
une paroi. Ils se développent dans des milieux
humides.
|
Ils consomment la matière organique en
décomposition (saprophytes) ou capturent des proies
vivantes (vers Nématodes): ils sont allotrophes.
|
La reproduction sexuée des champignons est
très complexe, les spores germent et donnent des
mycéliums qui se fécondent et donnent d'autres
spores.
|
|
Plantes
|
algues rouges et brunes,
plantes sans fleurs (mousses, fougères...) et
à fleurs
|
Les plantes sont composées de nombreuses cellules
réunies en tissus. Leur paroi les protège et
réunit les cellules entre elles. La plupart des
plantes vivent en milieu aérien et certains arbres
sont les plus grands êtres vivants.
|
Les plantes sont autotrophes et chlorophylliennes
(la chlorophylle est le pigment des chloroplastes sensible
à la lumière). Cependant, elles
possèdent des cellules chlorophylliennes (des
feuilles et des tiges...) et des cellules non
chlorophylliennes (comme les cellules des racines qui n'ont
pas de chlorophylle et consomment des sucres apportés
par la sève).
|
Les plantes se reproduisent par reproduction
sexuée (par les fleurs et les graines ou des organes
équivalents). Mais leur reproduction asexuée
est souvent facile: on peut multiplier de nombreuses plantes
en les fragmentant (boutures).
|
|
Animaux
|
invertébrés et vertébrés
(Poissons, Amphibiens, Reptiles, Oiseaux,
Mammifères)
|
Les animaux sont aussi composés de nombreuses
cellules regroupées en tissus. Les animaux vivent
nombreux en milieu aquatique mais ils se déplacent
souvent facilement et ont conquis tous les milieux
terrestres. Leurs organes des sens sont très
développés, et ils communiquent entre eux.
Certains vivent en société.
|
Les animaux sont allotrophes: ils consomment tous
des proies vivantes ou mortes. Certains se contentent de
filtrer l'eau dans laquelle leurs proies vivent, d'autres
ont des mécanismes actifs de capture qui sont de
véritables méthodes de chasse. Certains sont
charognards. En milieu terrestre de nombreux animaux se
nourrissent des plantes comme les insectes.
|
Les animaux invertébrés sont souvent
capables de reproduction asexuée mais cette
faculté diminue fortement chez les
vertébrés qui ne se reproduisent que par
reproduction sexuée.
|
On peut ajouter, avec Gœffroy Saint Hilaire, un
sixième royaume: le royaume de l'homme ou royaume
hominien avec une seule espèce: Homo sapiens,
animal, mammifère et primate social (du point de vue
biologique) mais qui peut être aussi considéré
comme le seul à avoir un esprit (mais celui-ci sort du domaine
d'étude de la biologie).
2. Il existe deux grands types
trophiques: l'autotrophie
(se nourrir seul) et
l'hétérotrophie
(ou plutôt
l'allotrophie - se nourrir
des autres)
Cette notion est juste
citée ici, comme élément qui sera repris dans le
programme des classes ultérieures: elle ne fait pas l'objet
d'une étude. Quand à l'affirmation du programme:
«Les activités fondamentale des cellules telles que le
métabolisme et la division sont sous le contrôle d'un
programme génétique»; elle relève pour
moi de la pétition de principe et je refuse de la colporter
(certains manuels scolaires n'en touchent pas mot d'ailleurs (Belin)
ou sinon sous forme d'exercice (Bordas, quoique la nouvelle
édition en fasse un paragraphe du cours)).
Quelle nourriture un organisme
consomme-t-il ? Peut-on modifier son régime alimentaire ?
Quelle nourriture minimale doit-il consommer pour survivre ? ....
Tant de questions et tellement de réponses différentes
pour chaque organisme.
2.1 La nourriture des êtres vivants se compose de
matière et d'énergie.
|
énergie
|
capacité de travail;
force vitale; en physique, elle est exprimée en
joules (J) et caractérise la capacité d'un
système à modifier l'état d'autres
systèmes en interaction avec lui
|
|
matière
|
minérale
|
qui contient des minéraux, composants des roches;
s'oppose à organique.
Pour les chimistes la chimie
minérale s'intéresse à tous les
éléments chimiques autres de le carbone (C);
on dit aussi la chimie inorganique.
L'eau et ses
éléments minéraux dissous est une
substance minérale. Mais l'eau dans la cellule n'a
plus les propriétés des substances
minérales pures. L'eau dans la cellule n'est jamais
un solvant (voir page
sur la cellule).
Il faut donc préciser l'état de l'eau:
minérale sous les trois états à
l'extérieur de l'organisme, liquide dans la
cellule (en très petite quantité),
liquide dans les liquides sanguins ou lymphatiques,
mais surtout eau interfaciale (plus proche de
l'état solide que de l'état liquide): on
pourrait parler d'eau intracellulaire mais le terme
est vague. La biochimie manque d'outil pour désigner
les états de la MATIÈRE VIVANTE.
|
|
organique
|
qui contient des composés du
carbone ET que l'on trouve dans les organismes
vivants; s'oppose à minéral; en
biologie les substances organiques sont classés en
glucides, lipides,
protides et acides
nucléiques: ce sont
des CHONPS
(composés de C (carbone), H (hydrogène), O
(oxygène), N (azote), P (phosphore) et S
(soufre)).
Pour les chimistes la chimie
organique au sens large s'intéresse à tous
les composés carbonés qu'ils soient ou non
issus du vivant. Mais il existe aussi une acception qui
rappelle une conception plus ancienne (Lémery 1690)
d'une chimie des substances organiques du vivant, ou
encore, chimie des substances organiques
naturelles.
|
|
On ne peut pas dire que le
CO2 est une substance minérale. Un
autotrophe ne se nourrit pas de carbone minéral. Il
ne se nourrit pas uniquement à partir de substances
minérales (ni plus ni moins qu'un allotrophe). Il
consomme du carbone oxydé et absorbe des substances
minérales avec sa boisson.
|
|
vivante
|
En résumé:
la matière vivante est celle des êtres vivants.
La distinction organique-minéral étant
délicate à utiliser il est
préférable d'étudier pour chaque
élément et pour chaque substance l'état
de la matière dans l'être vivant.
* l'eau (H2O): gazeuse, liquide,
interfaciale (liée à de grosses
molécules)
* des éléments minéraux, sous
forme d'ions, le plus souvent liés à de
grosses molécules
* des gaz: CO2, O2, le plus
souvent sous forme ionisée
(HCO3- et CO32-
pour le CO2) ou liées à de grosses
molécules transporteuses
* de petites molécules (sous-entendu
carbonées): acides, alcools...
* de grosses molécules ou
macromolécules (sous-entendu carbonées)
qui sont des polymères
(voir ci-dessous):
glucides complexes (sucres), acides
nucléiques (voir prochain
chapitre
ci-dessous),
protéines
* les lipides (acides gras et graisses) sont
classés à part car ce ne sont pas des
polymères mais ils peuvent constituer de grosses
structures (gouttelettes, plans ou membranes...).
|
|
Un
polymère est une molécule composée de
sous-unités identiques (ou monomères)
reliées par des liaisons fortes (covalentes)
Des polymères... comme des
colliers de perles (chaque perle est un
monomère)
|
homopolymères:
les perles sont toutes identiques
(un seul type de monomères)
|
copolymères
:
les perles sont de taille différente
(ici 2 types de monomères)
|
-O-O-O-O-O-O-O-O-O-O-O-
ou
-o-o-o-o-o-o-o-o-o-o-o-o-o-o-
deux types d'homopolymères sont
possibles avec deux types de
monomères
|
-O-O-o-O-o-o-o-o-O-O-o-O-o-O-O-o-o-o-
de très nombreux types de
copolymères sont possibles avec deux types de
monomères (cela dépend de la longueur du
polymère)
|
Une nourriture est ce qui permet à l'organisme de se
nourrir. Elle doit comprendre de la matière et de
l'énergie.
2.2 Il existe deux grands types trophiques et 3 types
accessoires
La matière peut provenir soit de l'air soit des autres
organismes, sans oublier la boisson qui est nécessaire pour
tous.
L'énergie peut être le soleil ou l'énergie
chimique contenue dans les minéraux ou encore dans les
composés organiques (pour des détails
voir &2.4 de l'ancien cours
d'immunologie de TS).
On regroupe les organismes par type trophique (du grec
"trophein" = "se nourrir"). On désigne par là un
type de travail de nutrition.
Un être vivant peut se nourrir seul, c'est-à-dire
sans l'aide d'autres êtres vivants, il est alors à
proprement parler autotrophe
(au sens étymologique de "se nourrit seul").
Un être vivant peut se nourrir des autres de
façon plus ou moins étroite, il est
hétérotrophe au sens large ou, mieux,
allotrophe
(ceci est un néologisme
formé à partir du grec "allo" = "les
autres").
Au sens restreint l'autotrophie est la capacité à
trouver sa matière carbonée dans le dioxyde de carbone
de l'air. Au sens large l'autotrophie englobe aussi la
capacité à utiliser l'énergie lumineuse du
soleil comme source d'énergie (phototrophie).
Au sens restreint l'hétérotrophie est la
capacité à utiliser la matière organique
comme source de matière carbonée. Au sens large
l'hétérotrophie englobe aussi la capacité
à utiliser l'énergie chimique (chimiotrophie) de
ses proies vivantes ou mortes ou d'hôtes parasités ou
associés en symbiose.
(La colonne de droite
n'est pas à mémoriser);
|
se nourrir seul
"autotrophie"
au sens
large
|
se nourrir
d'air et de soleil
(autotrophie (au sens restreint
au carbone) et
phototrophie)
Plantes, nombreux Protistes, certaines bactéries
(comme les bactéries bleues)
|
se nourrir d'air pour le carbone et de
minéraux pour
l'énergie
(autotrophie (au sens restreint au carbone) et
chimiolithotrophie)
certaines bactéries
|
|
se nourrir des autres
hétérotrophie
au sens large
ou
"allotrophie"
|
ne se nourrir que
des autres
(hétérotrophie
(au sens restreint au
carbone) et
chimiotrophie)
Animaux, Mycètes, nombreux Protistes, nombreuses
bactéries
|
se nourrir des autres pour sa matière
carbonée mais de soleil pour
l'énergie
(hétérotrophie (au sens restreint pour le
carbone) et phototrophie)
certaines bactéries
|
se nourrir d'air pour le carbone
mais trouver son énergie dans la matière
d'autres organismes
(autotrophie (au sens restreint pour le carbone) et
chimioorganotrophie
rares bactéries
|
Dans cette "classification" on peut peut-être mieux voir en
quoi le royaume des bactéries innove du point de vue du
métabolisme (colonne de droite). On aurait tendance à
considérer ces pistes comme autant d'étapes
évolutives aujourd'hui abandonnées, qui ne sont
conservées que dans quelques groupes reliques.
|
royaume
|
autotrophie
|
hétérotrophie
|
|
Procaryotes
|
x
|
x
|
|
Protistes
|
x
|
x
|
|
Mycètes
|
|
x
|
|
Plantes
|
x
|
|
|
Animaux
|
|
x
|
Remarques (voir TP):
Les notions qui suivent NE
SONT PAS AU PROGRAMME
mais elles ont déjà été utilisées
au collège; il ne s'agit ici que d'employer les mots sans
approfondir les concepts;
La photosynthèse (synthèse de matière
organique à la lumière) fait partie du
métabolisme des autotrophes phototrophes comme les plantes et
de nombreuses bactéries. La réception de lumière
(énergie lumineuse) et sa conversion en énergie
chimique se fait grâce à des pigments comme la
chlorophylle (phase de phototrophie) et s'accompagne de
rejet de dioxygène suite à la coupure des
molécules d'eau (photolyse de l'eau). L'énergie
chimique sert ensuite à la synthèse de matière
organique à partir du CO2 qui est donc consommé,
ce qui est le signe de l'autotrophie vis-à-vis
du carbone.
La respiration fait partie du métabolisme de nombreux
organismes aussi bien autotrophes que hétérotrophes,
phototrophes ou chimiotrophes. C'est un échange gazeux entre
le milieu extérieur et l'organisme. L'organisme consomme du
dioxygène et rejette du dioxyde de carbone. Le
dioxygène est utilisé, et le dioxyde de carbone est
produit, dans les réactions de dégradation
(catabolisme) de substances organiques, essentiellement pour produire
de l'énergie chimique pour la cellule (chimiotrophie). La
plupart des cellules eucaryotes respirent puisqu'elles
dégradent de la matière organique, quel que soit leur
type trophique. Ainsi, même les cellules chlorophylliennes, que
l'on pourrait croire autotrophes réalisent aussi une
chimiotrophie. Il faut donc réserver ces types trophiques
aux organismes et non à une seule cellule d'un
pluricellulaire.
Du point de vue du bilan, et en termes d'échanges
gazeux, photosynthèse (rejet de dioxygène,
absorption de dioxyde de carbone) et respiration (rejet de dioxyde de
carbone, absorption de dioxygène) semblent
antagonistes (avoir des échanges
opposés), mais les gaz absorbés et produits n'ont
ni la même origine, ni le même rôle. Ce bilan sera
repris dans le chapitre
sur l'écosystème
terrestre.
|
Respiration cellulaire
|
O2 -------> H2O
|
|
C6H12O6
--------> CO2
|
|
Photosynthèse
|
H2O -------> O2
|
|
CO2 -------->
C6H12O6
|
|
C6H12O6 + 6
O2
|
(1)
------------------>
<------------------
(2)
|
6 CO2 + 6 H2O
|
(1) équivalent au bilan de la respiration et (2)
équivalent au bilan de la photosynthèse (il faut
ajouter la présence de lumière)
|
Un petit exercice d'histoire...
|
|
Jan Baptist Von Helmont (1577-1644), comme les chimistes
de son époque, considérait que la terre n'est
pas un élément car elle résulte de la
transformation de l'eau. Il démontra son
hypothèse en faisant pousser un jeune saule dans une
caisse de bois contenant une quantité de terre bien
déterminée. Après arrosage, durant cinq
ans, avec de l'eau de pluie filtrée sur tamis, il
observa que le poids de l'arbre avait augmenté de
74 kg, tandis que celui de la terre n'avait
diminué que de 57 g. La terre n'ayant
accusé aucune variation sensible de poids, c'est donc
l'eau qui s'est changée en bois et en racines,
c'est-à-dire en substances solides que l'on
qualifiait de « terre ». (in EU)
|
|
Sais-tu quels sont les aliments essentiels de la
plante, en plus de l'eau de "boisson", qui ont permis au
saule d'atteindre 74 kg en ne consommant que 57 g de
"terre".
|
Remarque
(d'après Esquisse d'une
sémiophysique, Thom, 1988, p
69-71):
la morphologie des organismes repose sans aucun doute sur le travail
de nutrition.
* Pour les organismes de petite taille, unicellulaires procaryotes et
eucaryotes, la forme globale est une boule
(topologiquement), plus ou moins allongée, avec plus ou moins
de prolongements et la nutrition demande soit un passage des
nutriments ou déchets à travers les structures
limitantes (paroi par exemple), soit une déformation de cette
limite (phagocytose, exocytose...).
* Pour les organismes de plus grande taille (pluricellulaires) les
allotrophes doivent mettre en jeu des dispositifs de
capture des proies (tentacules, lacets, filets, pièces
buccales...) qui vont de pair avec des organes locomoteurs et
sensoriels de plus en plus sophistiqués; alors que les
autotrophes s'identifient à leur milieu
environnant en occupant l'espace, on croissant vers la
lumière, ce qui fait que leur forme est ramifiante
(leur croissance est dite indéfinie car elle ne semble pas
avoir de limite nette dans l'espace). Les botanistes ont depuis
longtemps modélisé la croissance des plantes avec des
algorithmes basés sur les fractales.
* certaines associations symbiotiques renforcent ces idées en
montrant comment deux types trophiques peuvent
s'interpénétrer: les lichens, associations de
mycétes et d'unicellulaires autotrophes conduisent à
des formes végétales en lames plus ou moins
ramifiées; ou encore les coraux, associations entre des
animaux diblastiques allotrophes et des unicellulaires autotrophes
conduisent à des "arborisations animales" qui les ont
longtemps fait appeler zoophytes.
2.3 Il existe 4 relations trophiques chez les allotrophes
Chez les allotrophes les relations trophiques peuvent être
de type consommation (un
animal phytophage par exemple mais aussi une bactérie sur un
cadavre ou encore un vautour sur une carcasse de mammifère),
prédation
(capture de proies: capture d'une bactérie par
un unicellulaire par exemple; mais aussi d'un nématode
(animal) par un mycète; ou encore d'une mouche par une plante
carnivore comme une Drosera),
parasitisme (un organisme
parasite vit de façon obligatoire aux dépens d'un
autre, appelé hôte; une tique sur un chien par exemple)
ou encore
symbiose (une
association à bénéfices réciproques; des
unicellulaires chlorophylliens avec des mycètes dans les
lichens ou des unicellulaires chlorophylliens avec des coraux).
3. Le matériel
génétique comprend de l'ADN, des ARN et des
protéines
Ni les ARN, ni les protéines ne
sont au programme (explicitement du moins) mais il est bien plus
facile (et cohérent) de les traiter ici SIMPLEMENT (en tant
que molécules polymériques) plutôt qu'en classe
de 1ère. De plus il est impossible de parler d'information
génétique et de gène sans parler de code
génétique.
Le terme "matériel" désigne
clairement la matière (mais on peut aussi y voir une
information). Génétique est pris au sens
étymologique de ce qui est transmis lors de la
génération. C'est le sens d'héréditaire
mais sans se laisser enfermer dans une génétique
moléculaire basée sur un concept fourre-tout de
gène. Une hérédité cytologique ou plus
biologique pourrait-on dire. Matériel génétique
veut donc dire "matériel dont la fonction principale est la
transmission héréditaire".
3.1
Les acides nucléiques (ADN et ARN) sont de longs
polymères de nucléotides
3.1.1 Les acides nucléiques sont composés de
chaînes de nucléotides.
Les acides nucléiques sont des polymères
(du grec poly = plusieurs et merein =
partage) (voir ci-dessus)
car il sont composés d'unités identiques
(les monomères). Comme il peut y
avoir plusieurs types de nucléotides on parle de
copolymères.
Les nucléotides sont des molécules
composées d'une base azotée (A= adénine,
T=thymine, U=uracile, C=cytosine, G=guanine), d'un sucre (ribose ou
désoxyribose) et d'un, deux ou trois groupement(s)
phosphate(s). Les nucléotides ont de nombreux rôles dans
le métabolisme (voir cours de 1ère et
TS).
3.1.2 L'ADN est une longue molécule en hélice
à deux brins
L'ADN (acide désoxyribonucléique) est
formé de deux chaînes (brins) de nucléotides
dessinant une double hélice dont le diamètre est de
2 nm.
Dans l'ADN le sucre est le désoxyribose (et l'on
parle de désoxyribonucléotides). Les bases sont
A, T, C ou G.
- Les nucléotides d'une chaîne sont
associés entre eux par des liaisons covalentes (entre le
groupement phosphate d'un nucléotide et le désoxyribose
d'un autre nucléotide).
- Les deux chaînes sont associées par des
liaisons faibles entre les bases (qui forment donc des paires A avec
T (2 liaisons faibles) et C avec G (trois liaisons faibles). On dit
que les bases sont appariées. La molécule d'ADN
est fragile est les deux chaînes peuvent facilement se
séparer (l'ADN est dit dénaturé) par chauffage
modéré (<50°C) et se réassocier
(renaturation) si la température redescend.
3.1.3.- Les ARN sont des chaînes à un seul brin
Les ARN (acides ribonucléiques) sont des
chaînes (un seul brin) de nucléotides pouvant se replier
et s'apparier partiellement.
Les nucléotides de l'ARN, contiennent du ribose
(ce sont des ribonucléotides). Les bases sont A,
U, C et G. Donc l'Uracile (U) remplace
toujours la Thymine (T).
TD - ADN-ARN
(les acides nucléiques); forme,
structure des molécules; simualisation*
3D (attention applet Jmol de 540Ko
à télécharger)
(* les superbes
applications qui visualisent les molécules en 3D sont des
simulations; pour souligner ce fait je parlerai de
simualisation)
3.1.4 - Les protéines sont de longs polymères
d'acides aminés
Il existe 20 acides aminés dans les protéines
du vivant. Ce sont des molécules organiques donc
composées de C, H, O, N et parfois S. Elles ont toutes deux
groupes d'atomes que l'on appelle des radicaux et qui leur donnent
leur nom: un radical acide (-COOH) et un radical
amine (-NH2).
Les protéines font partie du groupe plus vaste des
peptides. Le terme de peptide désigne toute
chaîne d'aa (aa réunis par des liaisons covalentes). Les
oligopeptides sont formés quelques aa (10...);
les polypeptides sont formés de nombreux aa
(10-100) mais lorsque leur poids moléculaire est
supérieur à 10.000 on préfère parler de
protéines. Les protéines sont souvent
formées de l'assemblage de plusieurs molécules et
contiennent des éléments non
peptidiques. Cette distinction faite on
utilisera en seconde le mot protéine comme équivalent
de peptide.
3.1.5 - La séquence des molécules
polymériques constitue une information linéaire
La suite linéaire (en
ligne) des nucléotides d'une chaîne de
l'ADN (l'autre étant déterminée par
complémentarité des bases), ou de l'unique chaîne
des ARN, forme une séquence propre à chaque
molécule d'acide nucléique. Elle peut être
représentée par la suite des bases (par exemple
.....AACTAGGCTTAA....).
De même la suite linéaire des aa d'une
protéine est appelée séquence ou
structure primaire.
Les séquences constituent une information
linéaire qui n'est pas sans rappeler l'information
exprimée par la suite de lettres d'un mot. Mais ce n'est pas
la seule information que peut donner une molécule qui se
déploie dans l'espace avec une forme. Même si la forme
est indubitablement liée à la séquence elle n'en
est pas la résultante car la forme dépend aussi des
conditions de milieu et de molécules additionnelles pouvant
s'ajouter pour donner une structure complexe. Par exemple l'ADN
s'associe avec des protéines pour donner un chromosome chez
les eucaryotes et les protéines sont nombreuses à
s'associer entre elles ou avec des ARN pour donner des complexes.
3.1.6 - La séquence de l'ADN peut être
altérée par mutation mais est habituellement
réparée
L'ADN est une molécule fragile et peut
être modifiée par de nombreux mécanismes
chimiques (méthylation...).
La séquence de l'ADN est considérée comme
très stable au cours de la vie de la cellule.
Les biologistes moléculaires appellent mutation
génique une altération de la séquence de
l'ADN. Certaines mutations géniques peuvent être
provoquées par des rayonnements (UV, RX) ou par des produits
chimiques. Dans la nature la plupart des mutations géniques
sont réparées par la cellule.
Remarque:
Les différences dans les séquences des acides
nucléiques (ADN et ARN) permettent de classer les êtres
vivants en 3 domaines: Bactéries,
Archéobactéries et Eucaryotes
On appelle mutation chromosomique une altération du
nombre ou de la forme ou de la taille des chromosomes.
3.2 L'ADN est associé à des
protéines dans les chromosomes lors de la division cellulaire
des eucaryotes
Un chromosome est étymologiquement une particule
colorable (du grec chromos = la couleur et soma = le
corps). Ces particules ont été observés et
décrites en 1879 par Flemming lors de la division (mitose) de
cellules animales. Depuis des chromosomes ont été
observés lors de la division cellulaire des toutes le cellules
eucaryotes. Les chromosomes sont composés d'ADN
(acide désoxyribonucléique) et de
protéines. On parle parfois abusivement de chromosome
pour tous les filaments d'ADN de grande taille des cellules
eucaryotes ou procaryotes. On ne parle pas de
chromosome pour les plasmides ou pour l'ADN des mitochondries ou des
chloroplastes. Effectivement les chromosomes se colorent à
l'aide de colorants basiques (parce qu'ils sont composés de
molécules acides). Mais si l'ADN peut se colorer, même
sans être à l'état de chromosomes, pour que l'on
puisse le voir au MO il est nécessaire qu'il soit
condensé dans un chromosome.
L'ADN est très compacté dans un chromosome.
Dans une cellule humaine il y a 46 chromosomes (regroupées en
23 paires de même forme et taille) soit environ 2 m d'ADN
déroulé. Chaque chromosome humain ne dépasse pas
quelques µm de longueur.
Avant chaque division cellulaire l'ADN est dupliqué
(c'est la réplication de l'ADN), ce qui permet de
répartir un même matériel
génétique dans les deux cellules filles issues de la
division cellulaire. Pour les procaryotes la boucle d'ADN est
répliquée en deux boucles réparties dans les
deux cellules filles. Pour les eucaryotes la réplication de
l'ADN donne des chromosomes doubles (les deux parties
identiques de chaque chromosome ou chromatides sont
attachées au niveau d'un centromère). La cellule
les scinde en deux puis répartit chaque chromatide dans une
cellule fille (c'est la mitose). Elle divise ensuite son
cytoplasme (c'est la cytodiérèse).
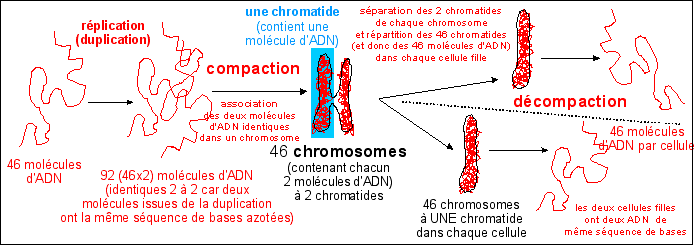
Duplication, compaction, association,
séparation, décompaction de l'ADN dans une cellule
humaine qui se divise en deux cellule filles.
Le chromosome comme structure TRANSITOIRE lors de la division
cellulaire d'un eucaryote (formation des chromosomes par compaction
de l'ADN, séparation des chromatides, disparition des
chromosomes par décompaction de l'ADN)
Remarque:
Un petit calcul classique: les chromosomes sont des pelotes
d'ADN.... une comparaison avec de la laine
|
|
dans une cellule
humaine
|
facteur de
multiplication
|
ANALOGIE avec une cellule agrandie
à la taille de la classe
|
|
|
volume de la cellule
(un cube de côté 10
µm)
|
103 µm3 =
environ 103. 10-18 m3 =
10-15 m3
|
la longueur du côté du
cube est multipliée par 0,5. 106
le volume du cube est multipliée par 1,25
1017....
|
53 m3 = 125
m3
|
volume de la classe
(un cube de 5 m de
côté)
|
|
en gardant ce même FACTEUR DE
MULTIPLICATION, on peut en déduire la longueur de la
molécule d'ADN et son diamètre à
l'échelle de cette classe
|
|
longueur total de l'ADN par cellule
à répartir en 46 filaments
(chromosomes)
|
2 m
|
0,5. 106
|
2. 0,5. 106 m = 1000 km
|
Un fil de 1000 km de long et 1 mm de
diamètre pourrait être équivalent
à l'ADN d'une cellule humaine réparti en 46
filaments.
Avant une division on peut considérer que les 46
filaments se dupliquent (46 doubles filaments) puis lors de
la division se condensent (en 46 paires de pelotes
accrochées 2 à 2).
Ces 46 paires de pelotes permettent ainsi de séparer
les 46 filaments doubles lors de la division, ce qui serait
"autrement coton" si les filaments étaient
déroulés !!!!
|
|
diamètre de la molécule
d'ADN
|
2 nm = 2.10-9
m
|
0,5. 106
|
2.10-9. 0,5.
106 m = 1 mm
|
|
Les chromosomes sont des
structures (protéiques) de compaction et de
séparation de plusieurs molécules immenses
d'ADN. Sans chromosomes les 92
molécules d'ADN d'une cellule humaine en fin de
réplication formeraient un fouillis
inextricable....
Remarque:
il est probable que de nouveaux modèles de chromosome
voient le jour avec non plus une molécule unique mais
de petits ADN circulaires correspondant chacun à une
sous-unité de lieu (dans le noyau) et de fonction
(liée au métabolisme). Voir remarque
ci-dessous.
Encore plus attirant un modèle de chromosome comme
structure stable de la catastrophe mitotique: l'œil du
cyclone en quelque sorte; où les filaments ne sont
plus que des artefacts d'une dynamique figée par le
MET.
|
3.3 Certaines séquences d'ADN, les
gènes moléculaires,
servent de modèle pour la synthèse de molécules
polymériques
IMPORTANT
Le terme de gène
désigne, historiquement une particule
héréditaire transmise, support des
caractères que l'on qualifie de mendéliens (en hommage
à Gregor Mendel).
Cette définition est
claire et ne génère aucune confusion. Mais on lui a
superposé une notion fonctionnelle découverte dans le
cadre de la biologie moléculaire. On s'est efforcé,
tant bien que mal, de les unifier à l'aide de la
théorie de l'information génétique (voir
cours
de terminale S, spécialité
SVT). Pour préparer
les élèves et éviter les confusions, je propose
de préciser dès la seconde (et aussi en
1ère) la notion à laquelle on se
réfère:
*
j'emploierai
gène
moléculaire
si c'est
notion de gène, unité
fonctionnelle de la biologie moléculaire.
* j'emploierai
gène
héréditaire
pour désigner la
particule
héréditaire,
sens qui a déjà été vu en
troisième et qui sera revu en terminale.
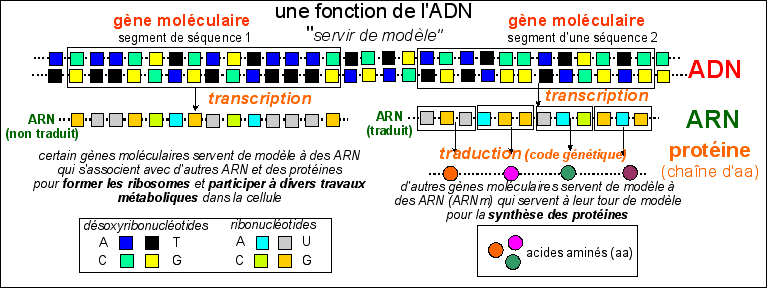
Cette ILLUSTRATION ne rend pas
compte de la grande taille des segments d'ADN impliqués ni de
celle des ARN (quelques centaines à quelques dizaines de
milliers de nucléotides); de même, les protéines
sont des molécules de plusieurs dizaines d'acides
aminés (aa) et il existe de petits peptides qui sont
synthétisés par d'autre voie que celle
représentée ici (par les NRPS, La Recherche, dec
2003).
Un
gène
moléculaire est un
segment d'ADN
dont la fonction est d'être copié en
ARN.
La copie d'un segment de l'un des brins de l'ADN en
ARN s'appelle la
transcription.
|
|
L'ensemble des
gènes moléculaires forme le
génome.
|
Certains ARN sont
traduits en protéines.
Les gènes moléculaires qui sont copiés
en ARN puis traduits (selon le dictionnaire du
code
génétique) en protéines forment
le protéome. On
parle des gènes moléculaires des
protéines. On dit que la cellule
exprime le gène
moléculaire d'une protéine lorsque le
gène est transcrit puis traduit en une
protéine.
|
On notera que la fonction de l'ADN
est passive. L'ADN est transcrit. C'est la cellule qui
synthétise la molécule, à partir du gène
moléculaire, on dit que « la cellule EXPRIME le
gène moléculaire ».
Le sens usuel d'
«expressions» est «un mot ou une suite de mots
présentant un sens qui dépasse le sens
littéral». On désigne peut-être par
là le fait que la fonction du gène moléculaire
va au-delà de l'ARN et englobe aussi celle de l'ARN qui peut
à son tour servir de modèle (au peptides). On peut
aussi dire, utilise, fait fonctionner..., ce qui fait moins appel
à un pouvoir
imaginaire.... On a aussi pris
l'habitude de dire qu'« un gène CODE pour une
protéine». Mais il faut bien comprendre que si la
présence d'une protéine nécessite un gène
moléculaire, l'inverse n'est pas vrai. On peut aussi dire que
tout l'ADN n'est pas "CODANT". Il serait préférable de
dire qu'un gène moléculaire "est associé"
à une molécule. En fait, au sens
expliqué ici, le gène moléculaire n'a
qu'une fonction: servir de
modèle; toutes les autres fonctions
dérivées sont imaginaires....car indirectes. Le
gène moléculaire est un modèle
matériel et non au sens propre une unité
fonctionnelle (au sens d'une
fonction globale car toute molécule a une fonction locale -
voir plus
haut).
La génétique est la science des
gènes
(moléculaires et
héréditaires).
La génomique est la science des gènes
moléculaires.
|
|
Le matériel génétique
est l'ensemble des molécules qui participent de la
fonction des gènes moléculaires. Il comprend
l'ADN, les ARN et, d'une façon dérivée,
les protéines.
|
L'information génétique STABLE
est l'information contenue dans les gènes
moléculaires.
Comme la seule fonction de l'ADN connue
précisément est la transcription,
l'information génétique est la l'information
contenue dans l'ADN et copiée (transcrite) en ARN:
c'est donc la séquence de l'ADN ou de l'ARN. Par
extension on peut aussi dire que l'information
génétique comprend la séquence des
protéines traduites à partir des ARN.
L'information
génétique est la séquence de
l'ADN ou de l'ARN ou encore des protéines.
La seule information génétique STABLE est
celle de l'ADN.
L'information génétique est une
information linéaire.
C'est la séquence (suite des monomères)
des molécules polymériques (ADN, ARN,
protéines).
|
On peut modifier les gènes moléculaires de certains
organismes, par transgénèse, pour leur faire
produire des molécules qu'ils ne produiraient pas
ordinairement. On peut ainsi ajouter de nouveaux gènes
moléculaires ou remplacer des gènes moléculaires
déficients. Mais il est évident que la cellule
n'utilise pas toujours ces gènes moléculaires
étrangers, surtout si l'on ajoute à un organisme un
gène moléculaire qui vient d'un autre organisme
très éloigné. La plupart du temps la cellule est
incapable d'EXPRIMER un gène moléculaire
étranger.
On peut aussi injecter dans une cellule des ARN qui parfois sont
traduits. Mais comme leur durée d'utilisation est très
limitée (voir 1ères S) la cellule n'est pas vraiment
transformée.
Tout l'ADN n'a pas de
fonction connue et n'est donc pas organisé en gènes
moléculaires.
Pour l'homme, le protéome constitue 1,2% de l'ADN et moins de
10% de l'ADN a une fonction
connue
(voir
cours de 1ère
S).
Une page d'analyse
des expériences de F. Griffith
(1928)
On peut en retirer d'une
part la très grande qualité du travail de Griffith et
l'excellence de sa publication; d'autre part l'exploitation plus
controversée qui a été faite de certaines de ses
hypothèses, et de l'importance grandissante de la biologie
moléculaire dans les années qui suivirent sans aucun
doute à relier à la prospérité de
l'institut Rockfeller aux Etats-Unis.
3.4 Le matériel génétique est au centre des
théories de l'hérédité
3.4.1 - Allèles, génotype et
phénotype sont des notions héréditaires
associées aux chromosomes (donc aux eucaryotes)
Les notions
d'allèles , de génotype et de phénotype
ne sont pas des notions de la biologie moléculaire mais des
notion héréditaires ; les termes ont été
utilisés pour la première fois semble-t-il en 1902 par
Bateson - voir cours
de Terminale S, spécialité
SVT. Les allèles
sont des traits de caractères reliés à un seul
gène héréditaire .Un gène
héréditaire est une particule
héréditaire (unité mutable appartenant
à un groupe de liaison ou chromosome). Un
allèle c'est une portion de chromatide.
|
|
définition HISTORIQUE claire
|
|
gène
héréditaire
|
Un gène héréditaire
est une portion de chromosome associée à la
transmission d'un caractère héréditaire
(uniquement chez les eucaryotes)
|
|
allèle
|
Un allèle est la
forme d'un gène
héréditaire.
|
|
génotype
|
L'ensemble des formes d'un
gène héréditaire constitue le
génotype de ce gène
héréditaire.
|
|
phénotype
|
Le phénotype d'un
gène héréditaire c'est la forme
visible de ce gène, c'est-à-dire les
caractères visibles associés à
la possession de ce gène
héréditaire.
|
Pour cette partie je recommande
chaudement l'étude de la transmission de caractères
héréditaires à l'aide de la théorie
chromosomique de l'hérédité, ce qui est
accessible à un élève de seconde. Allèles
dominants et récessifs, regroupement des caractères qui
sont transmis ensemble et qui correspondent à un chromosome
qui est ainsi assimilable au groupe de liaison des
généticiens. Enfin, plus délicate mais
accessible la notion de disposition linéaire des gènes
sur le chromosome, ce qui nécessite la notion de recombinaison
intrachromosomique dont la fréquence dépend de la
distance génétique. Ceci n'implique absolument pas que
l'on parle de crossing-over et de méiose. On laisserait alors
la partie de biologie moléculaire pour la terminale afin d'y
montrer les incompatibilités posées actuellement entre
une théorie chromosomique aux applications très
limitées et une biologie moléculaire en perte de
vitesse.
Gène
moléculaire et gène héréditaire NE SONT
PAS DES NOTIONS SUPERPOSABLES
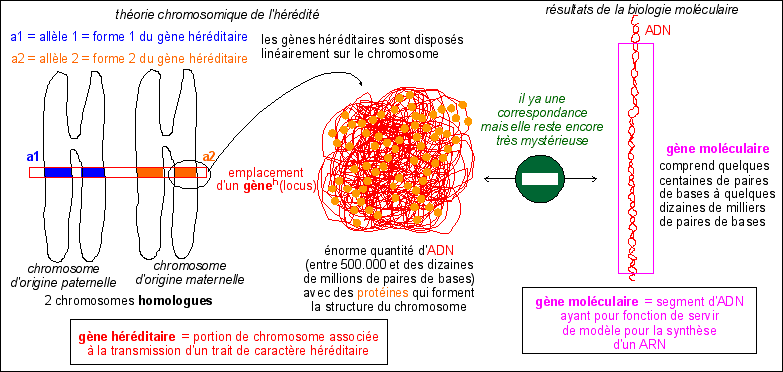
Les progrès de la biologie
moléculaire ayant été fait sur des procaryotes
il a été facile d'imaginer que l'ADN circulaire de la
bactérie qui comporte des gènes moléculaires
linéairement alignés était le reflet de
l'alignement des gènes héréditaires sur le
chromosome eucaryote. Mais c'est aller un peu vite. Il y a à
entre 10 et 1.000 ordres de grandeur entre la taille d'un gène
moléculaire et celle d'un gène
héréditaire. C'est pourquoi il est important de laisser
la place à des théories qui essayent de structurer le
chromosome autrement qu'avec une seule gigantesque molécule
où les gènes moléculaires seraient
alignés.
Il y a clairement une incapacité de
la biologie moléculaire à définir quand la
variation dans la séquence d'ADN, par mutation par exemple,
cesse d'autoriser à parler de variation d'un même
gène moléculaire ou d'apparition d'un nouveau
gène puisque la fonction de l'ADN est passive (l'ADN
est transcrit).
Dans le cadre de la théorie chromosomique de
l'hérédité, un allèle
désigne un trait de caractère
héréditaire. On exprime par le terme
d'allèle la variations des traits de caractères
reliés à un même gène
héréditaire. On ne peut relier ce trait de
caractère à une gène moléculaire qu'en
supposant une liaison unique et causale entre l'ADN et le trait de
caractère.
Ces notions d'allèle, de
génotype et de phénotype ne sont pas valables pour les
procaryotes mais uniquement pour les eucaryotes qui ont des
chromosomes. Pour un organisme qui possède un nombre pair
de chromosomes (on dit "diploïde")
hérités pour moitié d'un organisme paternel et
pour moitié d'un organisme maternel, les allèles sont
les formes différentes de chaque portion de chromosome ou
gène héréditaire portés par des
chromosomes homologues. Les chromosomes homologues portant alors les
mêmes gènes héréditaires aux mêmes
emplacements (c'est toujours la théorie chromosomique de
l'hérédité mais cela n'est ABSOLUMENT PAS UN
RÉSULTAT DE LA BIOLOGIE MOLÉCULAIRE. On ne voit pas de
gène moléculaire sur un chromosome. Les sondes
fluorescentes (FISH) ne peuvent pas localiser des gènes
moléculaires comme cela est parfois écrit dans les
manuels scolaires. Ils se fixent uniquement sur des séquences
répétitives d'ADN. On n'est pas au même niveau de
compréhension du chromosome. Le gène
héréditaire comprend des millions de paires de bases et
est probablement relié à de très nombreuses
segments d'ADN (circulaires ?) et donc de très nombreuses
protéines et donc de très nombreux gènes
moléculaires. Mais la liaison est loin d'être claire et
il est tout à fait probable qu'il n'y ait pas de disposition
linéaire des gènes au niveau d'un ADN
déroulé identique à celle de l'ADN
condensé chromosomique.
|
Le noyau contient les chromosomes
déroulés
ou
l'ADN et les protéines nucléaires se
condensent en chromosomes lors de la division
cellulaire
On a utilisé des colorants
fluorescents spécifiques de chacun des
chromosomes chez une petite "biche" femelle d'Asie (le
Muntjac) qui possède 6 chromosomes (les
cellules sont des fibroblastes).
Le noyau de gauche est au repos et on distingue les
territoires de chaque chromosome, bien
séparés; il faut donc comprendre le
matériel génétique comme une forme dans
un espace propre et non seulement comme une molécule
chimique.
Le passage de la
forme déroulée à la forme filamenteuse
(chromosome) peut s'expliquer par un modèle
mathématique où les chromosomes sont les
singularités associées à la catastrophe
mitotique (voir René
Thom).
(d'après la couverture de
Chromosome Research, vol 6, n°3, avril 1998,
modifiée et reproduite sans autorisation avec un
résolution si faible qu'elle empêche toute
utilisation autre que pédagogique...)
|
|
Des résultats complémentaires ont
été obtenus sur des fibroblastes humains (ce
sont des cellules conjonctives qui interviennent dans la
réparation des tissus endommagés, la
cohérence et la liaison des tissus entre eux)
|
Un article en anglais accessible sur le net librement
(licence Creative Commons Attribution) est
à l'adresse http://biology.plosjournals.org/archive/1545-7885/3/5/pdf/10.1371_journal.pbio.0030157-S.pdf.
en voici un extrait d'image (Fig. 1-D -
réf: 10.1371/journal.pbio.0030157.g001)
particulièrement accrocheur:
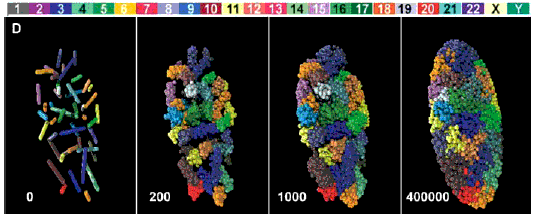
Le positionnement des 46 chromosomes (dans un état de
décondensation croissant de la gauche vers la droite)
d'un fibroblaste humain en reconstitution 3D simulant les
résultats obtenus par microscopie (voir l'article
pour des précisions)
J'aime beaucoup la légende ajoutée par La
Recherche (brève du n°388,
juillet-août 2005, 15):
Paysage
nucléaire
, qui rappelle le paysage
épigénétique de Waddington
(voir page
annexe).
Références de l'article:
Three-Dimensional Maps of All Chromosomes
in Human Male Fibroblast Nuclei and Prometaphase
Rosettes
Andreas Bolzer, Gregor Kreth, Irina Solovei, Daniela
Koehler, Kaan Saracoglu, Christine Fauth, Stefan
Müller, Roland Eils, Christoph Cremer, Michael R.
Speicher, Thomas Cremer
PLoS
Biology Vol. 3, No. 5,
e157 DOI: 10.1371/journal.pbio.0030157
|
S'il ne tenait qu'à moi je
garderai la notion d'allèle pour la terminale mais le
programme et les ouvrages scolaires ne cessent de l'employer depuis
même le collège (voir le programme
de seconde et les documents
d'accompagnement du programme
de
troisième)!!!!
À chaque fois que vous lirez allèle
(dans vos manuels scolaires, dans un article de journal, dans un
exercice scolaire...) vous comprendrez que l'on veut souvent parler
de la séquence d'un gène moléculaire.
Cette définition est fausse. Vous auriez intérêt
à garder celle donnée au collège de
"portion de chromatide" qui est incomplète mais
exacte car elle fait bien référence au chromosome des
eucaryotes. Et utiliser le terme de génotype
moléculaire à la place d' "allèle
moléculaire", ce qui ne veut vraiment rien dire.
|
|
définition
HISTORIQUE claire
|
sens
analogique
|
sens analogique
étendu
|
|
gène
|
Un gène
héréditaire est une portion de
chromosome associée à la transmission d'un
caractère héréditaire (uniquement chez
les eucaryotes)
|
Un gène
moléculaire est un segment d'ADN transcrit en
ARN et éventuellement en protéine (aussi bien
chez les procaryotes que chez les eucaryotes)
|
|
allèle
|
Un allèle
est la forme d'un gène
héréditaire.
|
 AUCUN sens analogique légitime
AUCUN sens analogique légitime
(ne pas employer "allèle" pour désigner une
séquence d'un gène
moléculaire)
|
|
génotype
|
L'ensemble des formes
d'un gène héréditaire constitue le
génotype d'un gène
héréditaire.
|
Par ANALOGIE les
séquences d'un gène moléculaire
constituent le génotype de ce gène
moléculaire.
|
Le génotype
moléculaire d'une cellule désigne
PAR EXTENSION toutes les séquences de tous ses
gènes moléculaires.
|
|
phénotype
|
Le phénotype
d'un gène héréditaire c'est la
forme visible de ce gène, c'est-à-dire les
caractères visibles associés à la
possession de ce gène.
|
Le phénotype
moléculaire d'un gène désigne PAR
ANALOGIE la forme visible de ce gène
moléculaire au niveau de la cellule,
c'est-à-dire l'ensemble des produits (ARN et
peptides) résultants de sa transcription et
éventuellement de sa traduction.
|
PAR EXTENSION le
phénotype moléculaire d'une
cellule désigne l'ensemble de tous les
produits de tous ses gènes
moléculaires.
|
On notera que l'on ne peut pas parler du phénotype
moléculaire ou du génotype moléculaire d'un
ORGANISME pluricellulaire.
Ces notions sont expliquées dans une page
annexe pour les 1ère
S.
3.4.2 Les mutations ne sont pas toujours associées
à des modifications du matériel
génétique
Ce paragraphe reprende et complète ce
qui a été dit dans le paragraphe 3.1.6
Le sens courant de mutation ne comporte pas de
référence implicite au hasard. C'est pourtant le cas
maintenant en biologie.
Une mutation c'est un changement
brusque mais stable (qui se perpétue au
cours des générations), ce qui permet de la
différencier d'une anomalie ou d'une malformation, non
transmissibles héréditairement.
Le mot, d'origine juridique ou économique
depuis le 13ème siècle, semble avoir été
utilisé par des biologistes comme Buffon (1766,
Histoire Naturelle des Quadrupèdes) ou Lamarck dans
le sens de changement biologique avant d'être utilisé
dans le sens de changement héréditaire par De
Vries (1903) puis étendu aux changements chromosomiques
par Morgan (vers 1910).
Petit à petit le biologiste s'est habitué à
considérer qu'une mutation a une cause inconnue (vague) en
rapport avec le matériel héréditaire. Il est
évident que tout changement brusque ne trouve pas son origine
dans le matériel génétique. Il faut donc bien
utiliser les adjectifs géniques et chromosomiques qui
précisent la pensée.
Une mutation biologique au niveau de l'organisme se définit
différemment selon le règne auquel appartient cet
organisme:
- pour un procaryote l'apparition d'un caractère inhabituel:
une déficience dans la synthèse d'un enzyme, un
changement brusque de la forme d'une colonie...est assez souvent mais
pas toujours relié à un changement dans l'ADN et donc
à une mutation génique.
- pour un eucaryote unicellulaire, on connaît des mutants
métaboliques pour lesquels des mutations géniques ont
été identifiées mais il serait bien
déraisonnable de penser que le terme de mutation puisse rendre
compte de la très grande variabilité ou des adaptations
plus ou moins héréditaires (pensez à
l'autotrophie des euglènes) de ces organismes. Tout changement
adapatatif stable transmissible héréditairement ne doit
pas être relié à l'ADN sans questionnement.
Une mutation peut aussi, dans un sens élargi, désigner
une modification stable du matériel génétique
qui peut donc toucher les chromosomes: modification du nombre de
chromosomes ou perte ou ajout d'une portion de chromatide d'un
chromosome. Ce sont les mutations chromosomiques. Parmi
les plus connues ont peut citer la trisomie qui est l'ajout d'un
chromosome supplémentaire , le plus souvent à la paire
21, ou encore les mutations observées par De Vries en
1901-1903 chez une petite plante appelée l'onagre (nom de
genre Œnothera, nom d'espèce lamarckiana),
qui lui servirent pour élaborer sa théorie
héréditaire des mutations associées à des
pangènes (les gènes héréditaires
étaient ainsi nommés, voir
cours de terminale
spécialité) et qui sont dues probablement
à des réarrangements chromosomiques (in
EU article "mutations").
Enfin, il existe de nombreux changement brusques stables,
transmissibles héréditairement, que l'on ne sait pas
relier à un changement du matériel
héréditaire. Ce sont les mutations au sens le
plus large.
Plus j'essaie de préparer
mon cours, moins le chapitre suivant du programme (Parenté et
diversité des organismes) me paraît cohérent. Et
pourtant la compréhension de l'embryologie est une clé
pour celle des mécanismes de l'évolution. Mais la
réduction des mécanismes embryonnaires au
contrôle génétique (et principalement celui de la
drosophile) est d'un telle partialité qu'il m'est impossible
de l'enseigner de façon cohérente (voir une page
détaillée ancienne, qui date de l'année 2000 :
le
travail du
développement
où des arguments pour une théorie plus biologique sont
développés). Si l'on se contente des exemples et
documents pris dans les manuels scolaires, les élèves
vont en retirer des schémas simplistes de contrôle
uniquement génétique qu'il me paraît dangereux de
colporter. Et combien les données sur les plans de
symétrie et d'organisation paraissent arriver comme un cheveu
sur la soupe, alors qu'ils sont sans aucun doute un fondement de la
classification. Ce chapitre tel que je vais le présenter est
une seconde version, toujours provisoire.
4. Parenté et
diversité des organismes
La parenté des organismes
résulte de la reproduction. Elle découle
directement de la théorie cellulaire élargie à
l'organisme pluricellulaire: on ne connaît d'être vivant
qu'issu d'un autre être vivant.
La diversité repose sur une théorie qui est celle de
l'évolution: les êtres vivants se sont
transformés au cours des temps géologiques.
4.1 - Les espèces regroupent des individus qui se
ressemblent, vivent ensemble et dérivent les uns des
autres
Cette définition est voisine de celle de Lamarck:
«un ensemble» ou «une collection d'individus
semblables ou presque semblables», «qui furent
produits par d'autres individus pareils à
eux» (La naissance du transformisme,
Lamarck, entre Linné et Darwin, Goulven LAURENT,
Collection inflexions, Vuibert/Adapt, 2001, p 19).
Vous devez connaître la définition de Mayr de 1989:
« une espèce est une
communauté reproductive de populations (reproductivement
isolée d'autres communautés) qui occupe une niche
particulière dans la nature »
(Mayr, 1989, cité dans l'article "A propos de
la notion d'espèce", Louis Allano et Alex Clamens,
Biologie-Géologie (Bulletin de l'APBG), n°3-1996, 471-472
).
Une niche écologique étant l'«ensemble des
conditions dans lesquelles vit et se perpétue la
population» (Hutchinson - 1957) .
4.2 - La reproduction est une multiplication des individus au
sein d'une espèce
On peut décrire la reproduction en terme de matière
et d'information (voir
ci-dessus).
Se reproduire signifie
produire un autre être vivant de son
espèce.
L'espèce (du latin "specia" = "ce qui se voit")
c'est la forme, non pas uniquement au sens
géométrique (la forme dans
l'espace) mais aussi au sens dynamique (ce qui
fait qu'un être vivant présente cette forme changeante
au cours de sa vie; pour l'homme par exemple, depuis l'embryon
formé d'une seule cellule au vieillard, en passant par toutes
les étapes de sa vie; pour une bactérie cette forme est
plus simple mais change aussi au cours de sa brève vie du fait
de sa croissance).
En fait on peut même parler de
forme au sens philosophique d'Aristote du principe de vie de la
matière (il divise l'être de toute corps matériel
en matière et forme). On pourrait donc dire que l'organisme
qui se reproduit donne vie à un nouvel être vivant
composé de matière et de forme. La
matière est en partie héritée (chromosomes,
cytoplasme...) et en partie nouvelle (synthétisée selon
le type trophique à partir de matière minérale
ou organique venant du milieu extérieur...). La forme est elle
aussi en partie héritée (forme génétique,
ou information génétique, forme cytoplasmique ou
information cytoplasmique et forme environnementale, ou information
environnementale) et en partie nouvelle (déformation,
transformation....tous ces mots désignant des processus
pouvant modifier la forme à transmettre par
hérédité).
Pourquoi a-t-on besoin de parler d'espèce ? Pourquoi ne
parle-t-on pas de reproduction au seul niveau d'un organisme ?
Parce que cette fonction globale n'est pas toujours le propre d'un
organisme mais est toujours celle de l'espèce. Non pas dans la
capacité à se reproduire, qui est bien une fonction de
l'individu, mais dans son utilisation. C'est le cas de nombreux
unicellulaires (les amibes acrasiales par exemple) et même de
bactéries (bactéries dites sociales) qui vivent en
colonies et qui peuvent réaliser cette fonction non pas chacun
séparément mais en la "déléguant"
à quelques individus. On considère que c'est un premier
pas vers la pluricellularité.
C'est donc une propriété de la société du
moins dans sa globalité. Un individu peut ne pas se reproduire
et dans ce cas sa vie s'arrête à sa mort. Mais c'est
impossible pour une espèce car sinon cela signifie qu'il y a
extinction. Une extinction signifie qu'aucun descendant ne
perpétue l'espèce. On voit que la notion
d'espèce, comme population interféconde qui se transmet
la vie sous une forme particulière (species en latin
c'est "ce qui se voit", c'est-à-dire la "forme visuelle" au
sens le plus courant de "forme"), est indissolublement liée
à la reproduction.
On arrive ainsi tout naturellement aux deux autres fonctions globales
de relation et de nutrition qui sont aussi liées à
l'espèce mais peut-être de façon moins
évidente. Dans la définition de Mayr de l'espèce
qui date de 1989 il est fait mention de la niche écologique.
Le mot écologique dérivant du grec oïkos
(habitat) on peut non seulement y voir l'espace, le lieu mais bien
aussi les habitants et donc les fonctions de relations entre les
êtres vivants sont très certainement comprises dans
cette définition. De la même manière la nutrition
comporte toujours un volet relationnel que cela soit pour la
recherche de l'énergie lumineuse ou la capture des proies. On
peut à mon avis avancer comme argument que la
délégation de ces deux fonctions à des cellules
spécialisées au cours de l'évolution, tout comme
pour les fonctions de reproduction, est un indice de cette
socialisation en cours chez les pluricellulaires.
4.2.1 Il existe deux types de reproduction: la reproduction en
solitaire (asexuée) et à la reproduction entre deux
individus de sexe opposé (sexuée)
(partie provisoire en cours de
rédaction)
- Lors de la reproduction asexuée, la
matière et l'information d'une cellule mère sont
réparties entre deux cellules filles.
Une cellule mère donne toujours par reproduction
asexuée deux cellules filles "identiques" vis-à-vis
de leur forme, c'est-à-dire de la même espèce.
Et cependant les cellules filles n'ont pas exactement la
même matière ni la même information que la
cellule mère.
La division cellulaire est au programme
de la classe de 1ère S. Nous
n'en voyons ici que des généralités.
Une cellule divise son cytoplasme (et répartit donc les
organites qu'il contient) en deux lorsqu'elle atteint
habituellement deux fois sa masse initiale après
croissance. Le matériel génétique est lui
aussi dupliqué avant la division puis réparti entre
les deux cellules filles. On peut penser que l'information qui lui
est associée est ainsi répartie
équitablement. Mais des modifications peuvent
ultérieurement apparaître, notamment chez les
pluricellulaires chez qui les cellules se spécialisent (se
différencient) et sont ainsi modifiées.
On remarquera que l'on peut aussi
considérer que l'une des deux cellules est toujours la
cellule mère (diminuée) et appartient à la
lignée maternelle, qui devient alors sans fin, tant que les
cellules ont des descendants. Le descendant fille étant
à l'origine d'une autre lignée... et ainsi de
suite.
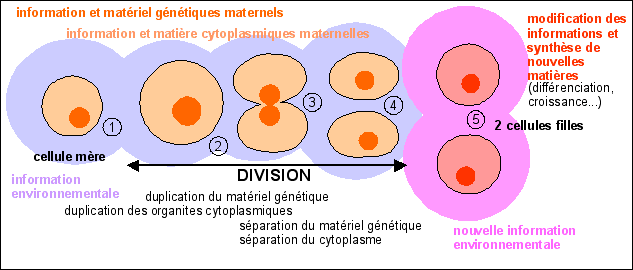
La reproduction en solitaire ou
asexuée
Les trois types d'information et de matière sont
nécessaires à toute division. Pour stopper une division
cellulaire on peut, soit modifier l'information ou la matière
extracellulaire (si on extrait un zygote (œuf
fécondé) de grenouille de sa gangue, il ne se divise
plus), soit modifier l'information cytoplasmique (la colchicine,
substance toxique extraite des racines des Colchiques (plante
à fleurs violettes qui apparaissent dans les champs en
automne), empêche la division des cellules eucaryotes), soit
modifier l'information génétique (si l'on enlève
le noyau d'une cellule eucaryote, elle ne peut plus se
diviser).
La reproduction asexuée est le
mode habituel de reproduction de très nombreux organismes
(Procaryotes, Protistes...). Pour certains c'est même le
seul mode de reproduction connu. Les Plantes (pensez au
bouturage...) et les Animaux invertébrés, par
exemple, se reproduisent facilement par reproduction
asexuée. Un fragment d'un organisme peut ainsi par
divisions, dans des conditions favorables, donner un individu
complet. Dans le cas d'un pluricellulaire comme toutes les
cellules ne sont pas identiques, pour se reproduire il faut non
seulement se diviser mais aussi que certaines cellules se
différencient (des cellules différentes se forment
à partir de cellules identiques, c'est ce que l'on appelle
la différenciation, qui sera vue en en 1ère
S).
- Lors de la reproduction sexuée deux organismes
de sexe opposés produisent des cellules sexuelles
(gamètes) qui fusionnent.
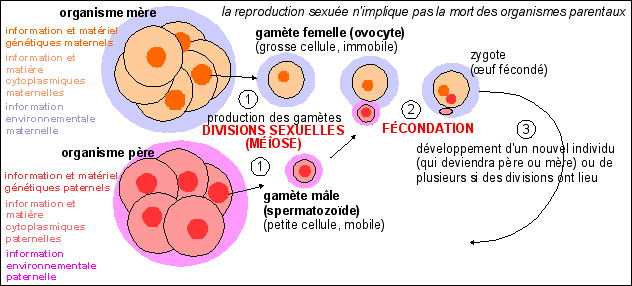
Une illustration des deux principales
étapes
de la reproduction sexuée (méiose et
fécondation);
les modalités en sont tellement différentes chez les
différents organismes des 5 royaumes qu'il est difficile de
faire un schéma global; celui-ci est bien imparfait. Il existe
par exemple des organismes à la fois mâles et femelles
(comme chez de nombreuses plantes à fleurs) mais la plupart du
temps les individus s'échangent des gamètes
mâles; ils ne pratiquent pas ainsi l'autofécondation (se
féconder soi-même). Dans le schéma proposé
on notera que la matière et l'information cytoplasmique
viennent quasi exclusivement du gamète femelle alors que le
matériel génétique et l'information
génétique du zygote proviennent pour moitié de
chaque cellule parentale. Enfin l'environnement est parfois maternel
(œuf, gestation...), très rarement paternel. La plupart
du temps les embryons se développent dans le milieu
extérieur mais avec des protections
particulières.
Remarque:
On trouve dans certains manuels scolaires une référence
au soma et au germen. Voici quelques mots sur cette très
ancienne théorie (le mot "soma" semble faire
son apparition dans ce sens en 1902 d'après le Petit
Robert).
Cette théorie stipule que les cellules germinales (sexuelles)
seraient immortelles car elles se transmettent par reproduction,
alors que les cellules somatiques (du grec soma: le corps)
seraient mortelles. En fait, en culture, on sait actuellement
maintenir en vie des cellules somatiques plus facilement que des
cellules sexuelles. Le maintien en culture passe cependant par des
possibilités de division et on observe toujours un
vieillissement de la culture qui se solde par un arrêt des
divisions, même pour les cellules cancéreuses qui,
pourtant, se divisent très facilement spontanément.
Remarque: l'opposition soma-germen a
peut-être un sens en embryologie mais cette distinction me
paraît artificielle chez un adulte; il faut peut-être
lier ces termes à l'association du psychologique (de l'esprit)
et du somatique qui a donné le terme psychosomatique
qui s'efforce de décrire l'union indissociable de l'esprit et
du corps dans la nature humaine cartésienne.
4.2.3 Se reproduire nécessite toujours trois types de
matière et d'information: environnementales, cytoplasmiques et
génétiques
Mais expérimentalement, afin de comprendre la part de
chacun, on peut s'efforcer de dissocier les éléments
intervenant dans la reproduction. On sait ainsi faire des transferts
de matériel génétique ou orienter l'information
génétique ou encore modifier le développement en
contrôlant l'environnement. Ne jamais oublier que provoquer la
divisions de cellules in vitro n'est pas reproduire un organisme. La
reproduction est un travail qui englobe tout l'organisme et les
manipulations in vitro ne sont que des moyens partiels d'en
reproduire quelques aspects.
Pour essayer de comparer l'importance de ces trois types
d'information (TOUTES TROIS NÉCESSAIRES CEPENDANT), voici
quelques observations et expériences:
* (Belin p 207) transplantation nucléaire d'un noyau d'une
cellule de glande mammaire de brebis adulte dans un ovocyte
énucléé (cellule sexuelle femelle mûre) de
brebis: certains techniciens affirment avoir obtenu un embryon (in
vitro) puis une brebis (par gestation dans un utérus de
brebis): Dolly (1996). Mais on a un très fort
taux d'échec et de nombreux animaux obtenus ainsi sont
anormaux. L'information extracellulaire est fournie par divers
procédés chimiques lors de la fécondation in
vitro puis par l'utérus pendant la gestation. L'information
cytoplasmique est celle de la brebis donneuse d'ovocyte.
L'information génétique est celle de la brebis donneuse
du noyau de cellule de glande mammaire. La brebis obtenue
possède de nombreux caractères extérieurs
(couleur du pelage...) voisins de ceux de la mère donneuse du
noyau. Cependant, tous les caractères qui font qu'une brebis
est une brebis et non... une chienne ou une souris, peuvent aussi
être fournis par l'un des trois types d'information. Il est
important de souligner que tous les essais de transplantation d'un
noyau d'une espèce dans un cytoplasme d'une autre
espèce (du moins pour des organismes pluricellulaires avec des
cellules différenciées) ont été des
échecs.(voir page sur le développement,
1.3
Expériences sur les ovocytes,
l'article de Axel Kahn - Clonage humain : un non
événement? Pour la Science, janvier
2002, 28-31 (où, malheureusement, il y a des amalgames entre
les termes d'ovule et d'ovocyte et aussi entre les différents
types d'information de la cellule) et une petite mise au point sur le
vocabulaire des manipulations
cellulaires (en cours de
rédaction))
* (Belin p 210) Si l'on fragmente une cellule eucaryote géante
d'Acetabularia, seul le fragment contenant le noyau et donc
l'information génétique est capable de survivre et
même de régénérer une cellule
entière. Cela montre la nécessite la présence
des 3 types d'information et aussi de la possibilité de
séparer l'information cytoplasmique en sous-unités
autosuffisantes.
Remarques: La présentation de ces
expériences (dans le
Belin et dans le Nathan) est
falsifiée:
* l'algue
Acetabularia n'est pas un simple unicellulaire mais un
cœnocyte,
c'est-à-dire un organisme possédant plusieurs noyaux
pour un seul territoire cytoplasmique (en fait l'acétabulaire
n'est cœnocytique que lors de sa reproduction mais elle est par
contre siphonnée c'est-à-dire que l'on peut
déterminer des territoires cytoplasmiques non
cloisonnés qui peuvent être considérés
comme autant ce cellules anucléées avant la phase de
reproduction...). Le genre
Acetabularia fait partie des Dasycladales, regroupées
dans les Chlorophycées cœnocytiques (thalles
siphonné à plastes chlorophylliens): leur thalle,
uninucléé à l'état
végétatif, est constitué par un axe
dressé (dans un gel calcifié) portant des ramifications
verticillées. Chez les Acetabularia , où l'axe
dressé peut atteindre 4 à 6 cm de haut, les
rameaux verticillés sont très minces (et caduques) et
plusieurs fois ramifiés; le noyau unique et très
volumineux est localisé à la base de l'axe. Lors de la
fructification, il se forme au sommet de l'axe un nouveau verticille
de rameaux qui constituent, par coalescence latérale, les
secteurs rayonnant d'un disque en forme de parasol ou de champignon.
Le noyau unique se fragmente alors en multiples petits noyaux qui
migrent dans les rayons du disque; ils deviendront les noyaux des
cystes; ceux-ci, entourés d'une paroi cellulaire
épaisse pourvue d'une fente circulaire délimitant un
opercule, sont libérés par destruction de la plante
mère; après un temps de repos plus ou moins long, de
nombreux petits gamètes biflagellés s'en
échappent et produiront, par copulation, des zygotes. (E.U.
article "Chlorophytes")
* Chez Acetabularia il y
régénération de l'algue complète avec
chapeau à partir d'un fragment de pédicelle
anucléé. Cependant
le cytoplasme capable de cette régénération doit
contenir des ARN spécifiques et donc de l'information
génétique.
(L'expérience 3 du Belin p 210 n'indique que le rôle du
noyau dans la reproduction (formation d'un nouveau verticille en
forme de chapeau après que le chapeau végétatif
soit tombé) et non dans la régénération
d'un chapeau végétatif).
Chez l'algue Acetabularia
mediterranea , les fragments anucléés
constitués d'un fragment du pédicelle non seulement
survivent jusqu'à deux mois, mais
régénèrent des parties manquantes telles que le
«chapeau». En pratiquant des greffes
interspécifiques entre deux espèces morphologiquement
différentes (A. mediterranea et A. crenulata ),
Hämmerling a cependant démontré que les substances
morphogénétiques proviennent obligatoirement du noyau
et diffusent dans le cytoplasme, où elles persistent et
déterminent les caractères morphologiques des segments
anucléés, pendant quelque temps seulement. On sait
aujourd'hui que ces substances «morphogènes» ne sont
autres que les ARN synthétisés dans le noyau et
transmis, associés à des protéines, au
cytoplasme.
L'utilisation moderne des techniques de greffe nucléaire,
mises au point dès 1939 par Comandon et de Fontbrune, a
conduit à préciser la nature des échanges
nucléo-cytoplasmiques. On a ainsi démontré, sur
des Amibes, que le noyau transmet des ARN au cytoplasme, tandis que
ce dernier synthétise et transmet des protéines au
noyau (Goldstein et coll.). Les échanges se font donc dans les
deux sens et, sur les Acétabulaires,
Hämmerling a montré
que le cytoplasme
contrôle le comportement du noyau tout autant que ce dernier
contrôle l'activité du
cytoplasme (E.U. article
"noyau cellulaire").
* Chez Acetabularia, on avait noté que près de 50% des
plastes ne possédaient pas d'ADN. Cette particularité
est maintenant expliquée par une hérédité
plastidiale monoparentale, résultant d'une
incompatibilité entre le génome nucléaire du
zygote et celui de l'une des lignées plastidiales. Cette
incompatibilité entraînant la
dégénérescence du nucléooïde des
plastes issus de l'un des gamètes (Biologie
végétale, t. 1, D. Robert et J.C. Roland, Doin, 1989, p
180).
4.3 Du zygote à
l'animal, le travail du développement
Le deuxième exemple du programme concerne le travail de
reproduction et d'une façon plus générale encore
le travail de développement chez les animaux:
c'est-à-dire le passage de la cellule œuf
fécondée (le zygote) à l'organisme adulte.
Nous choisirons un exemple dont nous essaierons de suivre le
développement en classe: l'hermelle, un vers marin de nos
côtes bretonnes.
TP 9
- Fécondation
et premières étapes du développement
embryonnaire chez l'hermelle
4.3.1 - Le travail du développement est fondamentalement
autonome
- 3.1.1 Le spermatozoïde est une petite cellule
sexuelle qui contient surtout une information
génétique
- 3.1.2 L'ovocyte est une grosse cellule sexuelle qui
contient une information génétique mais
surtout une information cytoplasmique ou mémoire
ovocytaire
- 3.1.3 Le zygote, cellule résultant de la
fécondation de l'ovocyte par le spermatozoïde,
est une grosse cellule protégée,
totipotente et polarisée.
- 3.1.4 L'embryon se divise d'une façon autonome
(segmentation) puis différencie des populations
de cellules spécialisées (tissus
embryonnaires) qui mettent en place les organes
(organogenèse) selon une progression autonome
- 3.1.5 L'individu termine son développement par le mise
en place de systèmes coordonnés ou
appareils
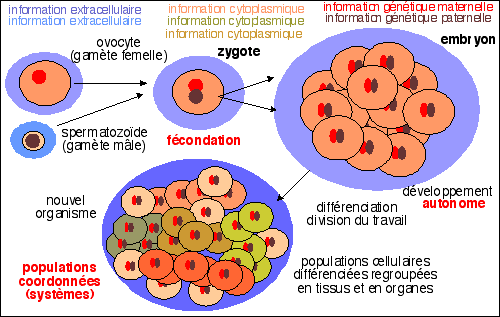
Une vision très simpliste du développement d'une animal
après la fécondation.
On notera que la multiplicité des informations
extracellulaires n'a pu être représenté dans le
nouvel organisme. Par contre la permanence d'une information
génétique identique dans chaque cellule est un point
important: cette information n'étant pas bien sûr
exprimée de la même façon dans chacune des
cellules.
|
Deux théories scientifiques
du développement...
|
Le développement embryonnaire ou
ontogenèse (du grec ontos, l'être
et genesis la naissance) semble obéir à
une croissance orientée qui semble
résulter elle-même d'interactions permanentes
entre les différentes populations cellulaires qui
s'engagent dans un processus de
progression autonome ou
différenciation autonome.
La première étape étant
l'activation de l'œuf (ou plutôt de
l'ovocyte, cellule sexuelle femelle, par la
fécondation (fusion avec le spermatozoïde) ou
même sans spermatozoïde, ce qui n'est pas un cas
si rare chez les animaux : le mécanisme étant
nommé la parthénogenèse).
Un individu est un ensemble composé de populations
cellulaires. A chaque instant, ce qui détermine
les caractères d'une population cellulaire, sont les
interactions qu'ont les cellules de cette population
entre elles et avec les populations avoisinantes. Une
cellule appartenant à un tissu donné,
présente telle ou telle activité par ce
qu'elle est issue d'une division qui résulte de
toutes une série d'événements et
d'interactions qui ont déterminé, à un
moment précis, son engagement dans tel ou tel
processus : le type de travail d'une cellule ou d'une
population dépend de son histoire
embryonnaire. (on remarquera que, dans cette
théorie, les informations essentielles sont les
informations extracellulaires et les informations
cytoplasmiques, alors que l'information
génétique n'intervient que comme une
information manipulée, exprimée par la cellule
en vue de réaliser tel ou tel travail qui
dépend de son histoire.
Cette théorie repose sur une
conception globale du vivant qui a de loin ma
préférence. Elle a été
élaborée dans le détail par Rosine
Chandebois, professeur d'embryologie à
l'Université de Provence (voir bibliographie,
et la page
sur le
développement).
|
|
Le développement résulte de la mise en
route progressive et coordonnée des différents
gènes de chaque cellule (libération
progressive de l'information génétique qui
contiendrait le plan complet et déterminé de
l'animal en construction). Il existerait un programme
génétique qui résulterait de
l'agencement des gènes et des interactions
déterminées et fixées à l'avance
entre les différents gènes, qui devraient
être activés à tel ou tel moment du
développement. Certains gènes appelés
gènes "architectes" ou gènes
homéotiques coderaient par des organes entiers ou des
"déterminants" de ces organes.
Cette théorie semble avoir
été émise pour la première fois
par un illustre physicien : Erwin Schrödinger
(1887-1961) qui dans son livre "Qu'est-ce que la vie ?",
publié en 1944, parle déjà de programme
génétique (voir une
histoire de la
génétique).
Elle a été développée par les
biologistes moléculaires et a surtout conduit
à des travaux sur la drosophile, petite mouche du
vinaigre. Elle semble actuellement dans une impasse (on a
pas trouvé dans le génome le début
d'une piste qui puisse nous faire penser à un
programme et bien évidemment elle soumet toute
information à l'information génétique,
ce qui est assez réducteur comme vision du vivant)
malgré le fait qu'elle soit enseignée
notamment dans l'enseignement secondaire, alors qu'elle est
en cours d'abandon par de nombreux
chercheurs.
|
4.3.2 - La part respective des informations extracellulaires,
cytoplasmiques et génétiques n'est pas la même
à toutes les étapes du développement
Quelques expériences du manuel Belin à
interpréter:
* n°4 p 251: que provoque une irradiation du zygote avec des
U.V. ? (pour une discussion voir page
sur la lumière du site associé): ici un arrêt
des mouvements cytoplasmiques nécessaires à la mise en
place de la symétrie embryonnaire à partir de la
mémoire ovocytaire. Il s'agit ici clairement d'une information
cytoplasmique.
* n°2 p 252: la séparation de 2 massifs embryonnaires au
stade 4 cellules ne permet de reconstituer un embryon complet chez le
Xénope que si le massif contient au moins une moitié du
croissant gris. Il s'agit encore clairement de l'information
cytoplasmique liée à la mémoire ovocytaire.
* n°2,3,4 p 256 ; 5 p 257; 1,2 et 3 p 260-261 : des mutations
homéotiques montrent clairement que l'information
génétique, manipulée par les cellules est une
composante importante du développement.
La racine latine homœo orthographiée
homéo en français viendrait du grec
homolos = semblable et n'est donc pas différente de la
racine homo. Les homéogènes ou
gènes homéotiques ont été
définis chez la drosophile par les travaux de Edward Lewis
à partir de 1948 (Edward B. Lewis, Christiane Nusslein-Volhard
et Eric F. Wieschaus ont ainsi reçu le prix Nobel de
physiologie et de médecine en 1995 pour leurs travaux
concernant le contrôle génétique du
développement embryonnaire). Ces gènes "architectes" -
comme les a nommé rapidement le grand public - avaient
été postulés car on observait une modification
importante et reproductible du plan d'organisation de la mouche
adulte (l'homéose ou homéosis désignant le
changement d'une partie du corps en une autre) à la suite
d'une mutation. Des homéoses ont été
décrites bien avant la connaissance des gènes : on cite
notamment William Bateson, qui , en 1894, en étudiant les
variations intraspécifiques chez un coléoptère,
observa notamment l'apparition de pattes à la place des
antennes. Il fit des observations similaires chez les
végétaux, où les étamines pouvaient par
exemple être remplacées par des pétales. Il n'y a
pas de raison, sauf idéologique, de penser que toutes les
homéoses sont d'origine génétique.
Les termes "gènes du développement " désignent
aussi des groupes de gènes isolés d'abord chez la
drosophile puis chez des vertébrés et dont certains
possèdent des séquences communes avec certains
homéogènes (voir le travail du
développement). Plus on étudie ces gènes
plus il devient apparent qu'ils sont très répandus chez
de nombreux organismes, qu'ils présentent plusieurs fonctions
différentes et qu'ils ne contrôlent pas une partie
spécifique de l'organisme mais peuvent être
activés dans des parties différentes et à
différents moments du développement.
* des expériences de greffes (déplacement de parties
embryonnaires) chez les amphibiens montrent que, selon les
étapes du développement et selon les tissus
déplacés, il peut y avoir compensation (les
tissus greffés perdent leur spécificité et
s'intègrent aux tissus en place) ou formation d'un organe ou
tissu surnuméraire: dans ces expériences, la
composante extracellulaire de l'information est primordiale.
* si l'on place une blastula d'amphibien dans un milieu salé,
la gastrulation se fait vers l'extérieur (exogastrulation) et
non vers l'intérieur, ce qui est aussi une indication d'une
information extracellulaire déterminante pour la
cohésion et les déplacements relatifs des cellules.
4.4 - La théorie de l'évolution affirme que les
espèces se sont transformées au cours des temps
géologiques
4.4.1 Des théories évolutives
L'évolution ou transformisme s'oppose au
fixisme selon lequel les espèces ne se transforment pas
ni ne se sont transformées. Comme l'évolution a des
arguments paléontologiques très solides on peut
l'affirmer scientifiquement comme théorie au
passé: les espèces se sont
transformées. Mais pour en faire une théorie
actuelle (les espèces se transforment actuellement) c'est un
autre problème que nous réservons pour la classe de
terminale. Il vous suffit de savoir qu'il y a plusieurs
manières de voir l'évolution et surtout les
mécanismes qui nous semblent importants selon que l'on est
paléontologue, anthropologue, botaniste, biologiste cellulaire
ou... informaticien. Les théories de l'évolution
soutenues par les uns et les autres ne se retrouvent guère que
sur l'idée de base d'une évolution au passé.
4.4.2 Des classifications phylogénétiques
Nous avons maintenant tous les éléments pour
comprendre les principes de la classification des êtres
vivants. Classer nécessite de comparer. A la profonde
unité structurale et fonctionnelle vue au niveau cellulaire et
au niveau de l'information génétique dans les chapitres
précédents, on va substituer une recherche des
fondements de la diversité, notamment au cours du
développement des pluricellulaires.
Cette classification doit pouvoir rendre compte de l'évolution
des espèces au cours des temps géologiques. Les
espèces dérivant probablement les unes des autres (la
phylogénie, mot créé dans le sens actuel
par Haeckel, vient du latin phylum= la lignée, ou du
grec phulon = la race; et génie, genèse =
la génération ou de genus = le genre), les
mécanismes évolutifs doivent s'appuyer sur des facteurs
de diversité à partir de points communs à
l'ensemble du vivant.
|
facteurs d'unité
|
facteurs de diversité
|
|
la cellule avec tous ses composants et organites, y
compris son information génétique,
universellement composée d'ADN
|
cellule procaryote ou eucaryote
|
|
une seule cellule (unicellulaire)
|
plusieurs cellules différenciées
(pluricellulaire) se répartissant le travail du
vivant (division du travail)
|
|
des cellules presque identiques issues d'une cellule
mère et formant une colonie (du point de vue de
l'information génétique, on parle de clone
lorsqu'on a une population de cellules qui ont la même
information génétique)
|
des cellules diversifiées
(spécialisées) en populations (tissus) qui
restent cependant unies et coordonnées (organes) pour
former un organisme
des organismes différents formant une
société
|
|
des gènes en nombre fini codant pour des
molécules particulières, des gènes
très voisins se retrouvent dans des groupes
très différents
|
des allèles variés correspondant à
des versions différentes de ces gènes
un même gène, dans deux cellules
différentes, ne conduit pas forcément aux
mêmes produits: l'expression de l'information
génétique dépend de l'environnement
cytoplasmique et extracellulaire
des mutations peuvent affecter les gènes et
être parfois conservées
|
|
chaque organisme maternel produit des cellules sexuelles
(ovocytes) qui gardent et transmettent la mémoire des
caractéristiques du groupe (mémoire
ovocytaire)
|
l'information génétique des cellules
sexuelles maternelles ou paternelles (germinales) est
différente de celle des cellules non sexuelles
(somatiques)
|
|
les conditions de développement sont toujours les
mêmes au sein d'une espèce (information
extracellulaire)
|
en modifiant les conditions de développement on
peut modifier le développement embryonnaire de
façon grossière (embryon à deux
têtes ou autres monstruosités) mais aussi
beaucoup plus fine avec des produits injectés dans
l'embryon ou en réalisant des greffes
|
|
le nombre de types cellulaires formant des tissus
(histologie) est réduit (par exemple les
systèmes nerveux des animaux sont composés de
quelques types cellulaires très semblables dans les
différents groupes)
|
l'arrangement dans l'espace, les symétries, la
forme des organes, la répartition des types
cellulaires (anatomie) permet la réalisation d'une
grande variété de formes organiques
|
Un exemple: une classification des
animaux
(les noms en italiques correspondent à des
embranchements, les points de suspension indiquent qu'il y a
d'autres groupes non cités):
|
diblastiques:
animaux possédant deux feuillets embryonnaires (le
feuillet externe donne une couche de cellules
orientées vers le travail de relation et le feuillet
interne possède des cellules plutôt
orientées vers le travail de nutrition et de
reproduction) - pas de symétrie - tous marins
|
Spongiaires (éponges),
Cnidaires (coraux et autres organismes
possédant des phases méduses sexuées et
des phases polypes, asexués),
Cténaires
|
|
triblastiques:
animaux possédant trois feuillets embryonnaires (le
feuillet externe est surtout composé de cellules qui
s'orienteront vers le travail de relation, le feuillet
interne vers le travail de nutrition et le feuillet
intermédiaire vers le travail de reproduction et de
relation) - la plupart du temps symétrie
bilatérale pouvant être
altérée dans les phases terminales de
l'embryogenèse
|
acœlomates:
pas de cœlome, c'est à dire une cavité
qui creuse le feuillet intermédiaire de l'embryon
|
Plathelminthes (vers plats)...
|
|
pseudocœlomates:
le feuillet interne se creuse secondairement d'une
cavité après le développement
embryonnaire
|
Rotifères,
Némathelminthes (vers ronds)..
|
|
cœlomates:
le feuillet interne est creusé chez l'embryon d'une
ou de plusieurs cavités
|
prostomiens:
l'orifice de la blastula ou blastopore donne la bouche de
l'embryon
|
Annélides (vers annelés),
Mollusques (à corps mou dont les
Gastéropodes, les Lamellibranches et les
Céphalopodes),
Arthropodes (à "pattes"
articulées dont les crustacés, les
insectes...)...
|
|
deutérostomiens:
l'orifice de la blastula donne l'anus de l'embryon
|
Echinodermes (dont les oursins et les
étoiles de mer à symétrie
pentaradiée),
Tuniciers,
Vertébrés...
|
N.B. dans cette classification, toujours en usage, les
critères principaux sont d'origine embryonnaire. Ce sont eux
qui déterminent les types cellulaires et les symétries.
La phylogénie est sous-jacente si l'on prend en compte la
complexification et la diversification des tissus entre la
première ligne et la dernière ligne du tableau.
Une remarque pédagogique: à mon avis une classification
utilisable est une classification
mémorisable.
Annexes:
L'information est parfois un concept flou en biologie et on pourrait
souvent l'abandonner au profit de la forme et des fonctions
(partie plus particulièrement à
l'attention des collègues)
Comme le mot information est souvent flou
(voir
annexe) je vous propose ici
quelques expressions de remplacement, évidemment plus
complexes car faisant moins appel à l'imaginaire:
|
Notion floue
|
notion
précise
|
théorie de l'information
génétique
(celle de vos livres scolaires)
|
Comme l'information génétique
est passée du côté
obscur de la ... causalité,
voici des affirmations non
contestables par un biologiste moléculaire (me
semble-t-il) laissant le champ libre à d'autres
manières de voir
|
|
Un gène est une unité d'information
génétique.
|
Un gène
moléculaire est un segment* de l'ADN dont
la séquence de nucléotides est
associée à d'autres molécules
polymériques.
Les gènes peuvent correspondre à des
séquences de l'ARN et aussi correspondre, par
l'intermédiaire des ARNm et par un code, le code
génétique**, à la séquence
des peptides, polymères d'aa.
|
|
L'information génétique de la cellule est
contenue dans son ADN.
|
L'information
génétique est une information
linéaire pour des molécules
polymériques: elle est contenue dans l'ADN
et certains ARN. Une cellule utilise son information
génétique pour synthétiser ses ARN et
ses protéines. Tous les gènes ne sont pas
utilisés en même temps par la cellule. Certains
gènes, chez des cellules âgées de
pluricellulaires, semblent être définitivement
hors d'usage.
L'ensemble des gènes d'une
cellule forme son génome. Tout l'ADN
n'a pas une fonction de type gène. Certains
gènes se chevauchent. Certains ARN courts peuvent
servir de modèle lors de la synthèse de
séquences d'ADN qui s'intègrent à l'ADN
du noyau. La matériel génétique
(héréditaire) n'est pas limité à
l'ADN mais comprend aussi des ARN et des protéines.
Les chromosomes n'ont pas fini de livrer leurs secrets.
|
|
Deux cellules d'un clone ont la même information
génétique
|
Deux cellules d'un clone ont seulement en commun
la même séquence d'ADN (du moins au
départ). Elles sont construites artificiellement par
des transferts d'ADN.
Mais comme on utilise la plupart du temps ce
terme pour désigner des cellules issues de
divisions binaires d'une cellule mère, ces
cellules ont en commun bien plus que la
séquence de leur ADN. Leurs cytoplasmes (avec tous
leurs organites si ce sont des cellules eucaryotes) sont
aussi hérités des divisions successives (et
des synthèses-formation successives) d'un unique
cytoplasme d'une cellule mère qui a donné
naissance à toutes les cellules du clone. Ce sont des
cellules "jumelles vraies"; pour les bactéries on
parle de colonie (voir manipulations
cellulaires)
|
* segment fait référence à une
molécule "linéaire" et donc à une portion de
droite.
** le
code génétique est
précisément et uniquement la correspondance entre des
groupes de trois bases de l'ARNm (codon) et un des 20 acides
aminés composant les protéines portés par des
ARN particuliers dits de transfert (ARNt) présentant des
triplets de bases complémentaires (anticodon). C'est donc une
correspondance entre des triplets de l'ARN et des aa. Il ne faut pas
employer ce terme à la place d'information
génétique.
Remarques:
* Les fonctions de l'ADN
puisque le gène est une unité de fonction
(modèle), à quoi peuvent bien servir les autres
séquences de l'ADN ? Ou plutôt l'ADN n'a-t-il pas
d'autres fonctions ?
On s'est focalisé sur la séquence des gènes
puisque l'on cherchait une information génétique
linéaire. Il est temps d'ouvrir les yeux. À
dématérialiser la notion de gène (La
mystique de l'ADN, Dorothy NELKIN et Susan LINDEE, 1998, Belin, Coll.
Débats) on en a oublié de s'intéresser de
près à la forme et à la fonction du chromosome
mais aussi des molécules d'ADN, dans leurs différents
états au cours de la vie d'une cellule.
Le chromosome est un élément matériel, il
possède donc un forme et donc à la fois
une composition chimique, une structure, une dynamique, une
cinétique de formation, de déplacement, de clivage, et
de décondensation,... et bien sûr des fonctions ....bref
il est tellement plus qu'une particule héréditaire ou
qu'une pelote d'ADN.
L'ADN (à l'état non condensé dans le chromosome)
est aussi une molécule matérielle et donc
possède aussi une forme en un lieu donné;
ce qui veut aussi dire qu'il n'y a pas que la composition chimique ou
sa séquence qui soit importante. Il faut s'intéresser
aussi à son degré de tension (on parle aussi de son
"tortillement"), aux molécules qui lui sont associées
(groupements méthyle, protéines, ARN courts...),
à son accessibilité, à sa dynamique de
synthèse, de réparation, de transport, de
dégradation.
* Linéarité et
circularité
L'ADN est un filament. Pourquoi ? À quoi sert d'habitude un
fil (plus ou moins élastique...) ? À lier (attacher),
à relier deux points (un chemin courbe), à
étendre - déployer (l'ADN se déploie dans la
cellule en se mettant à la portée d'un maximum de
molécules au sein du nucléoplasme (Eucaryotes) ou du
cytoplasme (Procaryotes)). Comment imaginer un filament d'une
longueur aussi monstrueuse stable dans la cellule ? À partir
d'arguments géométriques, René Thom propose (in
SSM (stabilite.pdf) , p 338) que chaque filament d'ADN se scinde en
de multiples petites sous-unités circulaires, chacune au
centre d'une sous-unité métabolique et
énergétique (énergide) lors de l'interphase. On
sait que l'ADN bactérien est sous forme circulaire, celui des
mitochondries, des chloroplastes, des plasmides aussi. Des ADN
viraux, non circulaires, doivent se circulariser avant de pouvoir
être répliqués.... La structure proposée
pour le chromosome n'étant qu'un modèle très
perfectible, il est possible d'imaginer qu'en fait l'ADN s'y trouve
sous forme de boucles et non d'UN SEUL filament.
|
Je ne demande pas à mes élèves de
comprendre les confusions liées à la notion
d'allèle employée à tort en biologie
moléculaire mais je veux bien essayer de l'expliquer
POUR DES COLLÈGUES. Cette question est très
difficile car elle nécessite de sortir du paradigme
génétique dominant.
|
|
Voici quelques exemples qui montrent les limites de cet
emploi:
|
lisez
|
comprenez
...et posez vous des
questions
|
|
un allèle est la forme d'un
gène
|
la séquence d'un gène
moléculaire peut varier
légèrement sans que l'on puisse dire
que le gène est différent; les
séquences très voisines sont
appelées des allèles ...
mais elles ne peuvent pas
être situées n'importe où car
deux allèles d'un même gène ne
peuvent pas être situées sur le
même chromosome à moins que le
gène existe en plusieurs
exemplaires.
|
|
... donc définir un
allèle en biologie moléculaire
demande de définir une
séquence + une position dans
un chromosome eucaryote spécifique,
définie héréditairement; un
allèle n'est pas la séquence d'un
gène moléculaire.
|
|
chaque cellule diploïde possède (au
moins) deux allèles de chacun de ses
gènes moléculaires
|
deux séquences identiques situées
à différents endroits dans le
même génome sont
considérés comme des copies d'un
gène unique. Deux
séquences légèrement
différentes doivent, pour être des
allèles, être situées sur deux
molécules d'ADN différentes, l'une
d'origine paternelle, l'autre maternelle, ce qui
implique bien un lien avec
l'hérédité qui n'existe pas en
biologie moléculaire.
|
|
des traits de caractères
différents légèrement (par
exemple la forme des graines des pois
mendéliens) sont des allèles
|
deux traits de caractères (allèles
au sens exact de traits héréditaires)
ne sont pas obligatoirement dus ni tous les deux
à des modifications de l'ADN, ni liés
à un même gène
moléculaire. Il existe bien d'autres
déterminismes cellulaires de la forme qui
sont plus simples que l'intervention de
l'information génétique (ceci n'est
pas une phrase gratuite c'est une évidence
de bon sens: la forme d'un tégument qui se
ride, due par exemple à un départ
d'eau, ne peut être que très
indirectement reliée à une
information génétique; vouloir
à tout prix relier toute concentration,
toute composition chimique, tout agencement dans
l'espace à une information linéaire
est un OBSTACLE à la
compréhension).
Et si l'on tient absolument à rester
près du génome, voici un autre
exemple: on connaît des couleurs
panachées de certains pétales qui
sont dus à des ARN qui interfèrent
avec l'ADN: les ARNi (pour une
revue simple mais bien évidemment dans le
paradigme dominant voir par exemple
http://www.ac-creteil.fr/svt/Big_mag/Actualite/ARN/ARN.htm).
Les allèles ne sont donc dans ce cas pas
liés à un unique gène mais
bien à différents gènes ; on
parle de phénocopies (voir paragraphe
suivant). Dans ce cas précis je crois que
les ARNi ne sont pas hérités mais
synthétisés... mais ce n'est
peut-être pas le cas général.
Pour quelques données plus originales sur
les ARN hérités voir les travaux
précurseurs de Beljanski sur la page
d'histoire de la génétique.
D'autres données... que l'on ne trouve
pas dans les manuels scolaires:
Pour tous les
mutants de la Drosophile connus dans les
années 40 (travaux de Richard Goldschmidt)
et pour de nombreux autres mutants chez
différents organismes on connaît des
phénocopies (de phéno =
visible, en grec), c'est-à-dire des
individus qui présentent le même
aspect (par exemple une deuxième paire
d'aile comme pour le mutant bithorax) alors
que leur génome n'a pas subi de mutation
(par exemple pour le mutant bithorax de la
drosophile la phénocopie bithorax est
obtenue par simple traitement à
l'éther de l'embryon normal). Les
phénocopies de nombreux papillons sont
obtenues de façon tellement
fréquente, par exemple par simple
refroidissement des larves, qu'il est probable que
les très nombreux mutants (variants)
reconnus dans la nature ne sont en fait que des
phénocopies qui ne doivent donc pas
différer des autres papillons
génétiquement
(d'après Peter T. Saunders,
à l'adresse http://www.utc.fr/arco/publications/intellectica/n16/16_06_Saunders.pdf
; Intellectica est par
ailleurs une revue très recommandable
à tous les esprits curieux) .
Ces données, insuffisamment
diffusées, montrent combien le rôle de
l'environnement est essentiel dans le
développement. On connaît aussi des
phénocopies (notamment chez le ver
Cœnorhabditis elegans) qui seraient dues
à des ARN induisant des répressions
de gènes (voir cours de
1ère S) que l'on nomme ARN
interférants; ce qui serait dans ce cas une
information cytoplasmique.
|
On dit qu'un
allèle est la forme
d'un gène, ou encore, la version d'un
gène pour garder l'analogie avec
l'information écrite. Mais, comme un gène
moléculaire est défini par sa séquence,
où se trouve la limite entre un gène
(de séquence donnée) et un autre gène ?
On voit bien qu'il est nécessaire
- soit de définir le gène comme ayant une
position fixe (par rapport à un point origine sur la
boucle d'ADN bactérienne ou par rapport à des
repères fixes comme des bandes colorées pour
les chromosomes eucaryotes) - mais dans ce cas on supprime
aux gènes la possibilité de se combiner les
uns aux autres de façon variable (et il faut donc
trouver une forme qui en soit responsable) -
- soit de définir l'allèle non pas au niveau
de l'ADN mais au niveau des molécules
polymériques associées donc au niveau d'une
fonction à laquelle le gène moléculaire
participe mais qui n'est pas sous son contrôle
direct.
On peut dire que quelques mutations ponctuelles affectant la
séquence d'un gène moléculaire
n'altèrent pas la fonction de gène
héréditaire et que l'on a des
allèles mutés. Mais est-on certain
qu'une petite modification de séquence ne peut pas
changer radicalement le type de caractère
héréditaire transmis, c'est pourtant ce que
l'on affirme quand on dit qu'un gène
homéotique est un gène qui contrôle le
développement. Un petite modification du gène
moléculaire dit homéotique ne conduit
qu'à une petite modification de la protéine
associée mais à une grande modification de la
forme globale de l'individu. Pour cela on suppose fixe la
position et la fonction du gène moléculaire.
C'est fort discutable. Quand on étudie les
gènes héréditaires et qu'on les
regroupe en allèles on n'est jamais sûr qu'ils
correspondent à des formes voisines d'un même
gène moléculaire. Dans certains cas
particuliers on a bien mis en évidence qu'une
variation de la séquence du gène
moléculaire conduisait à une variation
allèlique mais ces cas ne sont-ils pas des exceptions
? Quand on travaille dans le sens biologie
moléculaire vers caractères
héréditaires on est toujours dans le
même paradigme sans jamais essayer d'en sortir; on n'a
pas la possibilité de proposer comme
interprétation que le changement d'ADN
réalisé par manipulation
génétique par exemple n'a pas seulement
changé la séquence d'une protéine mais
aussi sa fonction et pourquoi pas celle de beaucoup d'autres
qui lui sont associées. Ce n'est donc plus le
même gène héréditaire même
si on est parti d'une séquence voisine.
Si l'on insère (ou si on supprime) dans une cellule
une séquence d'ADN voisine d'une séquence
d'ADN connue (appelée gène moléculaire)
et qu'on observe que quequechose est changé dans le
fonctionnement cellulaire (par rapport à cette
même cellule sans gène modifié), on ne
prouve pas la fonction d'un gène moléculaire
donné. Sauf à supposer que l'ADN est
composé de sous-unités fixes car, on ne voit
que le résultat du travail cellulaire. Proposer un
déterminisme c'est supposer que la seule chose de
changée est l'ADN. Or je crois que c'est tout le
contraire que l'on voit. De petits changements dans l'ADN
provoquent de grands changements dans l'organisme. Dire que
les gènes se chevauchent ou se déplacent est
encore une vision statique. Dire que l'ADN est
organisé en sous-unités fonctionnelles
(probablement circulaires comme l'a proposé Thom) ne
veut pas dire que ces sous-unités ont une place fixe
À CAUSE DE LEUR SÉQUENCE. En affirmant cela
une image vient immédiatement à l'esprit,
c'est la chromosome coloré en bandes
caryotypiques. D'abord je
précise que l'on ne sait toujours pas à quoi
correspondent les bandes, pour ce que j'en sais. Ensuite
comment concilier cette mobilité avec la position et
l'association avec des gènes des bandes qui semblent
fixes ? En plaçant la fonction non dans la
séquence de l'ADN mais dans la structure
chromosomique que l'on peut extraire, c'est-à-dire la
bande colorée. En fin de compte cela reviendrait
à dire que la séquence d'ADN de la bande
n'est pas ce qui compte le plus mais bien les
protéines qui lui sont associées (si les
protéines sont suffisantes à décrire la
forme du chromosome). Dans ce cas, lorsque l'on manipule
l'ADN seul après l'avoir extrait d'une bande,
pourquoi le retrouve-t-on toujours au même endroit
dans le chromosome ? Peut-être tout simplement parce
qu'il s'associe avec des protéines (ou ?)
particulières de cette bande, du fait de sa
séquence. Les marquages in situ (FISH) ne marquent
que des séquences répétitives de l'ADN
mais il n'y a pas, à ma connaissance, de marquage in
situ des gènes sur un chromosome non
digéré et non déroulé
(techniques de peignage par exemple) (voir par exemple
http://www.pasteur.fr/recherche/unites/biophyadn/
).
Ce qui est contestable ce n'est par l'organisation
linéaire des gènes héréditaires
sur le chromosome (voir spécialité
de TS) mais c'est l'image que
certains ont d'un chromosome déroulé
linéairement et qui placent aussi linéairement
les gène moléculaire le long de ce chromosome
monstrueux sans se poser de questions (voir schéma
ci-dessus).
|
Remarques:
* il ne faut jamais oublier que l'ADN est manipulé par la
cellule qui est l'unité irréductible du vivant.
* de plus l'ADN peut être modifié par des
éléments cytoplasmiques (des ARN peuvent être
transcrits en ADN et s'intégrer au génome).
* Actuellement on s'aperçoit que la
quantité d'ADN et que la séquence de
l'ADN changent au cours de la vie d'une cellule: l'ADN
d'un ovocyte n'est pas le même que celui d'une cellule
différenciée; des ADN courts semblent s'intégrer
au génome au cours de l'ovogenèse; des ARN courts
semblent être capables soit de transmettre par voie
cytoplasmique ou nucléoplasmique (par le noyau) leur
séquence qui sera traduite en protéines sans passer par
l'ADN, soit d'être "retrotranscrits" en ADN et
intégrés au génome après le début
du développement. (voir page sur les manipulations
cellulaires)
|
POUR ALLER (beaucoup)
PLUS LOIN
Une analogie classique:
On a souvent comparé les
bases de l'ADN (A,T,C et G) avec les lettres de
l'alphabet, les gènes constituant des
mots, c'est-à-dire des suites de lettres ayant
un sens. On affirme ainsi qu'une série de
gènes activés en même temps
constitue une phrase dont les protéines
sont la traduction en langage utilisé par
la cellule.
Dans cette vision les gènes sont des unités de
fonction, ils ont un sens pour la cellule. Ce qui fait
parler de code, de gouverne, de contrôle... En tant
que mémoire héréditaire, et du point de
vue évolutif, les gènes se combinent entre eux
pour rendre compte de la complexification des structures et
des dynamiques de l'organisme. L'évolution c'est la
complexification, les pertes, les modifications des
gènes.
MAIS
de nombreux problèmes naissent de cette signification
(ce sens) donnée aux gènes
Quelques arguments pour un lecteur qui
ne serait pas élève de seconde...
car l'ADN peut être "lu" dans plusieurs sens, les
gènes se chevauchent; un gène peut donner deux
produits différents dans des conditions
cytoplasmiques différentes et donc ne pas avoir la
même fonction cellulaire; deux gènes
différents en retour ont souvent la même
fonction... de plus l'ADN est fortement redondant (de
très nombreux gènes présentent de
multiples copies dont seules quelques-unes sont
utilisées...)...
On cherche de nos jours à poser clairement la
question de la signification des gènes (voir par
exemple pour quelques questions timides le petit
dossier de La Recherche: qu'est-ce qu'un gène ?). De
façon plus profonde, c'est Jean Petitot qui a
parlé semble-t-il en premier de
sémiophysique (une science du
signe, du grec seméion = signe et
phusikos = naturel) et qui continue avec d'autres
à développer cette notion à la suite de
René Thom (voir R. Thom: Esquisse d'une
sémiophysique). Le projet est de «
préciser le rapport entre une situation dynamique
globale (le « signifié »), et la
morphologie locale en laquelle elle se manifeste (le «
signifiant »)(Thom, 1968f4). »
En biologie le sens d'une molécule c'est sa
FONCTION, ce à quoi elle sert. Il est donc essentiel
de se poser la question : A quoi servent les gènes
?
Et la réponse n'est pas aussi
compliquée que l'imaginaire de certains veut nous le
faire croire.
Avec du recul on peut dire que les gènes n'ont
que quelques fonctions toujours les mêmes qui peuvent
se résumer en une seule: servir de
modèle pour des molécules
polymériques (ARN ou ARN puis protéines). Les
gènes ne sont pas "lus" (au sens d'une lecture des
mots signifiants) mais sont "recopiés" dans un
matériau plus stable (l'ARN) puis
éventuellement convertis en molécules
fonctionnelles (les protéines). Les ARN non
convertis en protéines sont des molécules
fonctionnelles comme les protéines (ou plutôt
les complexes protéiques).
De façon simpliste et en
gardant la même analogie de l'alphabet, on peut dire
que les ARN sont parfois des mots (lorsqu'ils
ont une autre fonction que de servir de modèle stable
pour la synthèse des protéines) mais ce sont
surtout les protéines qui sont les
mots. Les phrases ne sont pas dans les
associations de gènes mais bien dans les
associations protéiques ou dans les
associations ARN-protéines.
Certes, ces protéines et ARN viennent du
"fonctionnement" des gènes mais les associations ne
se font pas au hasard. Dire qu'il y a
nécessité ou auto-organisation revient
à dire qu'il y a une finalité dans la
matière et donc, in fine, dans les gènes et
qu'il est inutile d'aller la chercher ailleurs. Ce à
quoi ne peut se résoudre le philosophe : si
finalité il y a il est urgent d'en rechercher les
contours (de rechercher les attracteurs ou les
prégnances au sens de Thom).
Considère-t-on que le poète n'est que le jouet
de l'alphabet ou même du dictionnaire des mots qu'il
emploie ? Son œuvre de création poétique
est bien de jongler avec les mots dont il dispose
culturellement (par mémoire)? On peut penser que la
cellule fait de même: avec le stock de gènes et
d'ARN qu'elle possède par héritage, elle
s'exprime en vivant par ses fonctions (protéiques
notamment). Ses finalités sont inscrites dans des
formes (les saillances au sens de Thom) qui
représentent les trajectoires possibles des
dynamiques.
Les partisans d'un programme
génétique, souvent
matérialistes, affirment que l'expression de tel et
tel gène à tel instant est le résultat
d'une finalité (aveugle ou nécessité
comme l'on voudra) due aux conditions dans lesquelles une
cellule vivante se trouve (l'environnement agit sur
l'expression de l'information génétique). Au
contraire, les partisans d'une finalité globale de
l'être vivant, pas nécessairement
matérialistes ni vitalistes si cette opposition
signifie encore quelque chose, cherchent à formaliser
de façon mathématique ce pour quoi une cellule
vit. Voici un citation de René Thom à ce
sujet.
|
|
extrait de "La notion de
programme en biologie" (René Thom,
1984f5)
I Introduction : La théorie en biologie
Depuis qu'à partir du 16e siècle, on a
commencé à ouvrir les cadavres afin d'en
scruter les organes internes, l'explication biologique a
pris un tour résolument techniciste. On assimile
l'organe à un outil, lequel, construit par l'homme en
vue d'une fin connue, ne présente aucun obstacle
à l'intelligibilité. Dès Harvey, ce
point de vue a obtenu d'indiscutables succès. Il est
indéniable que le coeur est une pompe qui refoule le
sang dans ces canalisations que sont nos vaisseaux. De
même, les poumons se comportent (sur le plan
mécanique) comme un soufflet (mais ici
l'interprétation mécaniste laisse de
côté la fonction physiologiquement essentielle
d'échange gazeux entre l'air et le sang). Le
squelette (os + articulations) permet une
interprétation mécanique immédiate.
Toutes ces descriptions ont abouti à la
théorie cartésienne de l'Homme animal-machine,
et ce n'est guère qu'avec le plus noble de nos
organes (le siège de l'âme), à savoir le
cerveau, que l'imagerie techniciste est quelque peu
restée courte. Mais, avec la naissance
quasi-simultanée, vers 1950, des ordinateurs et de la
Biologie Moléculaire, cette imagerie a connu son plus
récent et peut-être son plus glorieux avatar.
On a fait de l'ADN des chromosomes, du génome,
l'analogue du programme qui régit le fonctionnement
d'un ordinateur. Cette dernière interprétation
offrait, sur le plan conceptuel, un nouvel, énorme
avantage. En effet, l'imagerie techniciste des organes
soulève, de manière inévitable, le
problème de la finalité. Comment tous ces
mécanismes d'une haute efficience, d'une parfaite
efficacité, comment toute cette horlogerie
pouvait-elle se construire apparemment sans horloger ?
À partir du moment où l'on pouvait supposer
« codée » dans l'ADN du génome toute
la structure organique, le mystère s'effaçait,
puisqu'il suffisait d'imaginer que l'ADN, promu au rang de
démiurge, dirigeait toute l'épigenèse
de l'être vivant, à la manière d'un
ingénieur dictant ses ordres à ses
subordonnés. Moyennant quoi, on a pu rendre
admissible la finalité, rebaptisée pour la
circonstance téléonomie. Et cela d'autant plus
que la traduction ADN ? Protéines, via l'ARN messager
était un code au sens étroit, technique, du
terme, puisque cette opération associe à tout
triplet de nucléotides un acide aminé bien
défini. Or, dans le premier cas la situation est
toute différente : il s'agit de comprendre comment
« l'information » supposée incluse dans
l'ADN génique, peut spécifier la structure
tridimensionnelle de l'organisation embryonnaire, puis
adulte. (Je parle ici des Pluricellulaires, je reviendrai
sur le cas des Unicellulaires en fin d'exposé.)
Pratiquement toute la théorie biologique moderne
s'est trouvée enfermée dans l'homonymie
abusive de ces deux emplois du mot code, et cet abus de
langage l'a condamnée à une
stérilité conceptuelle dont elle n'est pas
encore sortie...
Comment sortir de cette impasse ? Il ne fait guère de
doute que seule une certaine audace théorique peut
permettre de faire avancer la question. Il faut songer
à réintroduire en Biologie l'imaginaire, cet
imaginaire sans lequel il n'est pas de théorie.
Déjà, dans l'interprétation techniciste
des organes, ne peut-on songer comme autrefois le
suggéra Bergson à renverser l'ordre des termes
? Plutôt que d'expliquer l'organe par l'outil, ne
faudrait-il pas plutôt expliquer celui-ci par
celui-là ? Autrement dit, l'imagination intuitive qui
a permis à l'Homo Faber de construire, bien avant
toute science constituée, des outils doués
souvent d'une remarquable efficience, cette intuition ne
trouvait- elle pas son origine dans un « inconscient
biologique » légué par l'évolution
phylogénétique de l'espèce ? Et ceci
d'autant plus que la plupart des outils ne sont en fait que
le prolongement d'actions, motrices ou physiologiques, et
que l'action dans son organisation spatio-temporelle, nous
est organiquement léguée. Il faut reprendre
ici l'axiome lamarckien : la fonction crée l'organe
non pas au sens banal qui voudrait qu'une structure
organique se crée ou se développe à la
suite de l'usage qui en est fait, mais de manière
plus abstraite, plus « platonicienne ». Toute la
régulation épigénétique et
comportementale d'une espèce repose sur une structure
formelle de caractère géométrique
(topologique) qui se réalise dans l'espace des
activités métaboliques de l'organisme ; une
« fonction » apparaît alors comme le
dispositif régulatoire partiel afférent
à l'homéostasie d'un paramètre
physiologique essentiel (teneur en réserves
énergétiques, en oxygène, en
déchets organiques... par exemple), et la
réalisation organique d'une telle fonction peut
comporter les agents physico-chimiques les plus
variés, comme les organes les plus divers. (Par
exemple, la communication entre animaux d'un même
groupe social peut employer des signaux olfactifs, sonores,
visuels... souvent dans un même but). Toutes ces
« fonctions » concourent ainsi à la
canalisation de la dynamique dans l'espace des
activités métaboliques, formant ainsi un
« attracteur » global (que j'ai proposé
d'appeler la figure de régulation de l'espèce
considérée). »
Je précise que l'article de Thom
représente 25 pages de théorie
détaillée sur cette idée.
|
La question de la
définition de l'information sera encore une fois posée
dans le cours de terminale S (voir spécialité,
thème 3). Ici notre
propos est de présenter une voie alternative basée sur
un emploi rigoureux du mot fonction (dans la mesure des
capacités mathématiques d'un professeur de SVT qui n'a
pas poursuivi de formation mathématique depuis la classe
préparatoire de maths sup biologie (actuellement BCPST); il
est évident que j'appelle de mes vœux une formation
continue en mathématique pour biologistes qui soit plus
orientée vers la biologie théorique que vers les
statistiques comme elle l'est actuellement, à ma
connaissance.
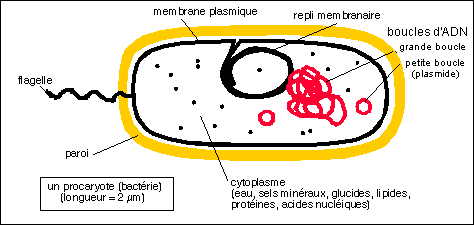
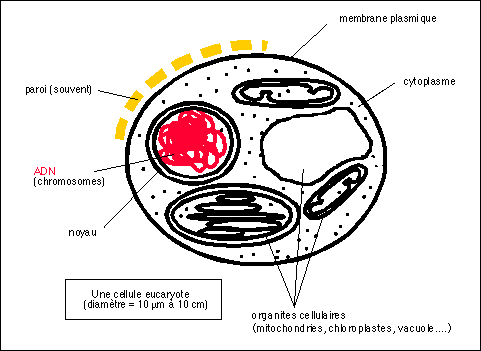
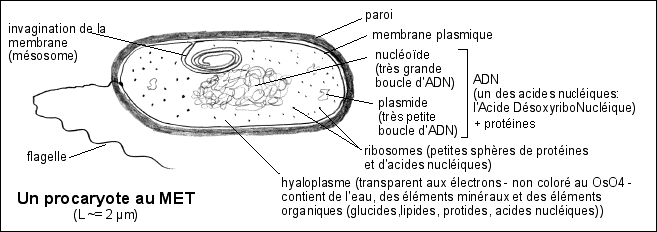{kind=link}
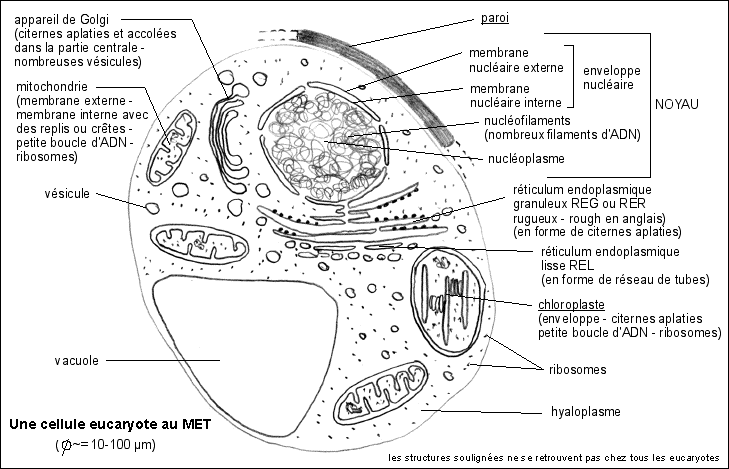{kind=link}