Sources:
Analyse génétique moderne,
Griffiths et al., 2001, DeBoeck Université (notamment pour la
définition de la norme de réaction)
Il y a deux attitudes à éviter:
* d'une part affirmer que la biologie moléculaire du
gène moléculaire permettra un jour d'expliquer tous les
niveaux du vivant, depuis le niveau cellulaire local jusqu'au niveau
social (cette thèse n'a plus beaucoup de
défenseurs)
* d'autre part condamner les théories chromosomiques de
l'hérédité - qui sont souvent revues à
l'aide des outils de la biologie moléculaire et qui confondent
alors gène moléculaire et gène
héréditaire - (ceci est par contre un danger
réel).
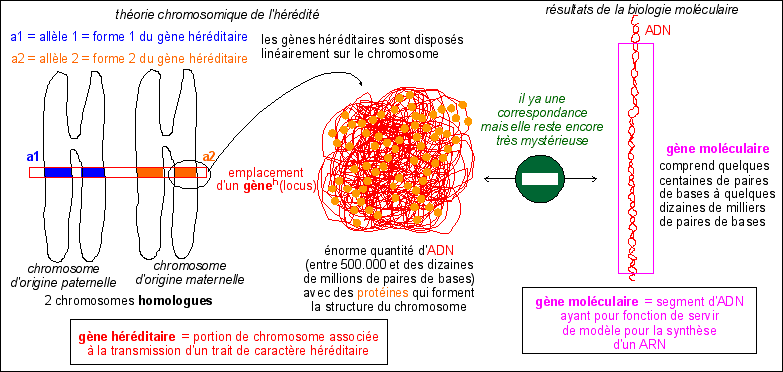
1.1 Le phénotype héréditaire est la forme visible d'un allèle. Le génotype héréditaire est l'ensemble des allèles d'un gène héréditaire.
1.2 Le génotype héréditaire d'un individu eucaryote n'est qu'une liste d'allèles correspondant aux seuls caractères héréditaires associés à des gènes héréditaires
1.3 Le phénotype héréditaire d'un individu eucaryote ne prend en compte que les caractéristiques visibles associées à des allèles et donc à des gènes héréditaires
1.4 Chez les procaryotes il n'y a pas de gènes héréditaires mais uniquement des gènes moléculaires: il n'y a donc ni allèle ni génotype héréditaire ni phénotype héréditaire, sauf par analogie avec les eucaryotes.
2. La notion de gène moléculaire peut être étendue à la cellule mais non aux pluricellulaires
3. Synthèse
|
Un allèle c'est une portion de chromatide. Un gène héréditaire est un segment de chromosome associé à la transmission d'un trait de caractère (voir cours de spécialité). |
|
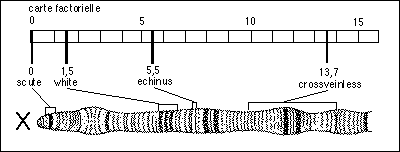
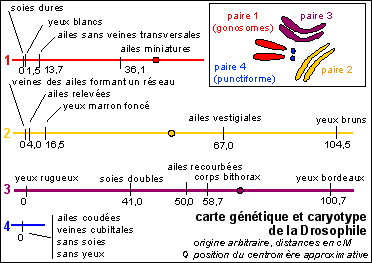
on établit des cartes génétiques ou cartes chromosomiques de ces allèles (ou encore carte factorielle). On établit ensuite des correspondances entre la position des allèles et la carte physique du chromosome, structure colorable du noyau des eucaryotes.
(deux sites de recombinaisons séparent
deux gènes
héréditaires) |
Il semblerait (http://www.esp.org/timeline/) que ce soit Johannsen (en 1909 ?) qui aurait forgé, à la suite de sa définition des gènes, au sens actuel de gènes héréditaires, les termes de génotype et phénotype (du grec phainein = paraître) lors d'études sur les caractères des pois. J'ajoute que génotype est daté de 1937 et phénotype de 1934 par le Petit Robert (ed. de 1984) mais sans autre précision. Je pense que l'on peut affirmer, historiquement du moins, que ces termes font bien référence à des caractères et donc à des gènes héréditaires et à des allèles au sens Morganien (voir cours de spécialité). On est donc clairement dans le cadre d'une hérédité chez des organismes eucaryotes.
|
On peut donc parler du phénotype héréditaire d'un allèle, qui est unique mais qui peut varier, chez différents organismes (ayant le même allèle) ou au cours de la vie d'un même organisme (avec son âge ou son état physiologique). |
|
|
Le génotype héréditaire, associé à un seul gène héréditaire, est vite devenu le génotype d'un individu. Pour un trait de caractère, et donc pour un gène héréditaire, le génotype héréditaire d'un individu est donc habituellement composé de deux allèles, l'un venant du père, l'autre de la mère. Si l'on étend la notion de génotype héréditaire à l'ensemble de tous les traits de caractères héréditaires on peut imaginer une liste d'allèles qui serait en quelque sorte la carte d'identité héréditaire d'un individu. Mais cela ne signifie en rien que toutes les caractéristiques anatomiques, physiologiques, éthologiques.... puissent être décrites par des traits de caractères héréditaires qui puissent correspondre à une hérédité chromosomique. Il suffit de voir les cartes chromosomiques actuelles pour être frappé par la faiblesse du modèle chromosomique à expliquer la transmission héréditaire de caractéristiques NORMALES d'une espèce (on attend en vain un gène héréditaire de la pilosité, de la forme d'un membre, du nombre de doigts...). L'explication habituelle étant que l'ordre supérieur est expliqué par la somme des interactions des éléments d'ordre inférieur. Mais il faut avouer qu'à part cette pétition de principe on ne trouve rien à se mettre sus la dent (voir page sur le développement). Il est temps de dire que LA PLUPART des caractéristiques de l'être vivant échouent à être reliés à des segments de chromosomes et donc à des gènes héréditaires. On peut donc sans hésiter affirmer qu'il y a encore de la place pour une théorie de l'hérédité non chromosomique. Personnellement je suis très tenté par des théories de la morphogenèse qui utilisent les outils mathématiques développés à partir des travaux de René Thom.
En tout cas je refuse d'enseigner sans critique l'idée que le génotype héréditaire pourra un jour être décrit comme l'ensemble de toutes les formes des gènes moléculaires de toutes les cellules au sein de leur environnement cellulaire et donc finalement décrire la totalité des caractéristiques de l'individu. Ce qui revient à dire qu'un jour on pourra relier tous les gènes héréditaires à des gènes moléculaires et que le génotype moléculaire décrira l'être vivant par des processus chimiques prévisibles. Ce qui est plus qu'un rêve mais une erreur de méthode (du complexe au simple; du discontinu au continu) et rend stérile la recherche si l'on persiste à garder ce seul modèle explicatif.
De la même manière que pour le génotype
héréditaire et étant donné les
interprétations globalisantes des mécanismes de
l'hérédité chromosomique, on a vite pris
l'habitude de parler non pas du phénotype
héréditaire d'un allèle, chez un individu
donné à un moment donné, mais du
phénotype héréditaire d'un individu,
comme ensemble du phénotype héréditaire de tous
ses allèles, à un moment donné; sorte de carte
d'identité phénotypique.
Phénotype
héréditaire est devenu un équivalent de
l'ensemble des caractères et l'on oublie souvent
d'ajouter "héréditaires" (au sens morganien). Sans me
répéter je pense que l'on pourrait dire que c'est comme
si on avait fait un rapide aller retour depuis l'allèle,
support d'un trait de caractère - placé dans un
chromosome par l'hérédité morganienne -, vers la
cellule et son ADN, pour revenir ensuite à l'organisme comme
somme de l'expression des gène moléculaire de toutes
ses cellules. C'est aller trop vite et trop loin. Tous les traits
de caractère d'un individu ne sont pas des allèles
parce qu'ils ne correspondent pas à des gènes
héréditaires étant donné que l'on ne peut
pas suivre leur transmission héréditaire par
hérédité chromosomique.
Ceux qui utilisent le terme de phénotype pour désigner
toute caractéristique visible (à quelque niveau que ce
soit: moléculaire, cellulaire, organique, individuelle,
sociologique...) font en quelque sorte allégeance à
cette vision d'un pouvoir descriptif pratiquement illimité de
la théorie chromosomique de l'hérédité
alliée à la biologie moléculaire du gène
moléculaire.
Si l'on part de la notion de gène moléculaire on ne
peut pas non plus affirmer que toute caractéristique
cellulaire dépend du fonctionnement d'un gène
moléculaire. On est encore loin de pouvoir décrire les
fonctions globales de la cellule, même procaryote, à
partir des gènes moléculaires. Il y a donc encore un
fossé entre le gène moléculaire et le
gène héréditaire. Fossé qui est peut
être du même ordre qu'entre
l'hérédité et l'hérédité
chromosomique.
Pour formuler ces notions émergentes de façon simple
et moderne on pourrait dire
* qu'un individu transmet d'abord à ses descendants des
caractères stables de l'espèce (c'est la
définition même de la reproduction). Cette
hérédité ne peut pas actuellement être
décrite par une hérédité chromosomique
ou, autrement dit, les chromosomes ne sont pas le support des
caractères de l'espèce; l'entité minimale
à transmettre est la cellule, recomposée
éventuellement à partir de plusieurs cellules
fournissant chacune une part de l'information nécessaire;
* qu'ensuite certains caractères héréditaires
plus ou moins stables que l'on pourrait qualifier de
caractères héréditaires individuels sont
transmis matériellement par les chromosomes.
Un chromosome (voir cours de
seconde) c'est une particule colorable. Il n'y a pas de
chromosome chez un procaryote.
Il n'y a pas d'hérédité chromosomique chez un
procaryote.
La notion de gène héréditaire n'existe pas
puisqu'elle est définie au niveau d'un chromosome.
Les gènes moléculaires portés par l'ADN
bactérien (de la boucle ou des plasmides) sont
dupliqués avant la division puis répartis dans les
cellules filles. Il y a donc une hérédité
stable, sauf perturbation environnementale.
Des fragments d'ADN peuvent aussi être transmis lors de la
conjugaison bactérienne qui fait suite à un accolement
de deux bactéries et la formation d'un pont cytoplasmique
entre les deux conjuguants dont l'un (donneur) transmet à
l'autre (receveur) la totalité ou une partie de son ADN.
Cependant il ne faut pas oublier que le génome d'une bactérie comme Escherichia coli atteint 4,2 millions de paires de bases pour quelques 2.000 gènes, ce qui est déjà assez complexe (des gènes se chevauchent, certains sont transcrits dans différents sens... bref, la biologie moléculaire des procaryotes a encore du travail). On n'a pas encore fait l'inventaire de tous les types de produits qui interviennent dans le métabolisme d'une telle "machinerie".
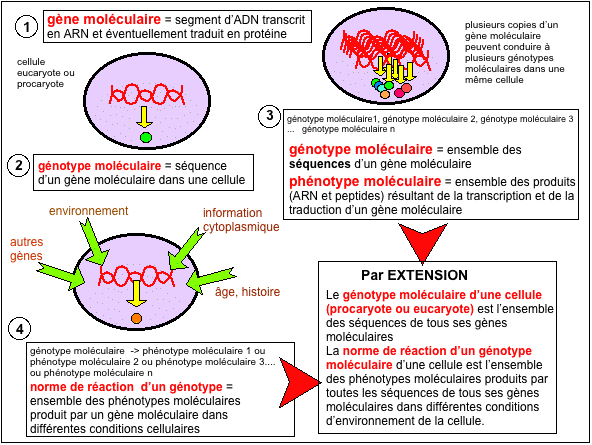
Certains s'essaient à combler le fossé entre
unicellulaires et pluricellulaires en définissant des notions
intermédiaires comme la norme de réaction. On
définit la norme de
réaction d'un génotype moléculaire
comme l'ensemble des phénotypes
moléculaires produits par un génotype
moléculaire donné dans différentes conditions
d'environnement.
Cette idée, séduisante au départ, me semble
finalement n'être qu'un moyen supplémentaire
d'embrouiller les idées. Cette notion n'est pas extrapolable
au pluricellulaire et ne peut surtout pas rendre compte de ce qui
viendrait moduler le génétique
(l'épigénétique). Elle reste toujours dans un
modèle génétique et
l'épigénétique n'est définit que comme
modulation du génétique alors qu'il peut être
tout autre (et ne devrait pas porter ce nom).
Dans un sens héréditaire la norme de réaction
contiendrait par exemple les phénotypes moléculaires
obtenus par phénocopies (voir cours
de seconde).
Toujours dans le cadre de la biologie moléculaire du gène, si l'on s'efforce de trouver des modèles pour rendre compte de la complexité des interactions entre gènes moléculaires on arrive à UNE ILLUSION DU CONTINU. Tout particulièrement chez les pluricellulaires. Si l'on somme tous les phénotypes moléculaires cellulaires dans le temps (embryonnaires, de différenciation, de croissance, de renouvellement cellulaire...) et l'espace (tous les tissus)...on obtient des variations de plus en plus continues. La variation discontinue (à petite échelle) est attribuée aux génotypes moléculaires. La variation continue (à grande échelle) est attribuée à la somme des génotypes moléculaires et à leurs interactions entre eux et entre l'environnement. En fait il y a non seulement perte d'information mais, plus grave, illusion du continu.
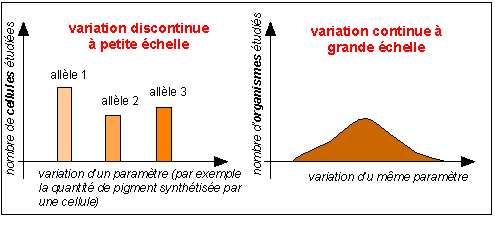
Remarque:
Voici une citation de Thom (in stabilité (SSM, 1968) p 213)
qui, à mon avis, précise pourquoi la notion de
phénotype moléculaire n'est même pas utilisable
pour un procaryote.
« Chez les Bactéries, le comportement
enzymatique ou la résistance aux antibiotiques sont des
composantes importantes du phénotype (peut-être plus
pour des raisons de commodité expérimentale que pour
des raisons intrinsèques) ; il importe de se convaincre que,
même en un sens aussi étendu, la définition du
phénotype comme une classe de formes modulo un groupe
d'équivalences G garde toute sa valeur. Par exemple, une
bactérie donnée, placée dans un milieu
donné contenant de la streptomycine à concentration c,
donne naissance à une colonie dont le développement
sera donné par une fonction N = F(c, t) ; il est clair
dès lors, que le caractère streptomycine
dépendant ou résistant est donné par le type
topologique de la fonction F(c, t). D'un point de vue abstrait, le
phénotype pourrait ainsi se définir comme l'ensemble
des réponses de l'organisme à toutes les perturbations
possibles du milieu et à la classification qualitative de ces
réponses. C'est dire qu'il est pratiquement impossible de
donner une définition formelle du phénotype ; la
Génétique dite formelle repose ainsi en entier, sur un
concept, celui de phénotype normal ou sauvage qui se
révèle impossible à formaliser. Mais c'est
là une situation qu'on rencontre communément même
en des disciplines plus exactes et un mathématicien ne saurait
s'en formaliser.»
>>> ou <<< désignent des extensions ACCEPTABLES de concepts en précisant bien à chaque fois le sens du mot
|
|
|
||||||
|
|
|
||||||
|
pas de chromosomes, pas d'hérédité chromosomique donc pas de gènes héréditaires |
|
|
|||||
|
|
|
||||||
|
|
|
||||||
|
V V |
|||||||
|
le génotype d'un individu unicellulaire reste une notion théorique (dans le sens d'inaccessible expérimentalement) même si on peut caresser l'espoir de relier de très nombreux caractères héréditaires à des gènes héréditaires |
le génotype d'un individu est une notion théorique (dans le sens d'inaccessible, expérimentalement) car un gène héréditaire n'est défini que par rapport à un caractère héréditaire observable et relié à un segment de chromosome; un gène héréditaire n'est pas défini au niveau de chaque cellule mais est le résultat du fonctionnement d'un très grand nombre de cellules |
||||||
|
|
|
||||||
|
V V |
|||||||
|
|
|
||||||
|
allèles, génotypes et phénotypes sont des notions HISTORIQUEMENT héréditaires mais il existe un sens analogique à ces termes en biologie moléculaire; les liens entre les deux sens ne sont pas encore intégrés dans une théorie globale et restent donc INCOMPATIBLES |
|||||||
|
|||||||
|
Vocabulaire ANALOGIQUE : gène moléculaire n'est pas égal à gène héréditaire allèle d'un gène moléculaire= séquence d'un gène moléculaire (il est préférable de parler de séquence allèlique d'un gène moléculaire) >>> |
|||||||
|
|
|
|
|
Même si l'on peut définir le génotype d'un pluricellulaire comme la l'ensemble de toutes ses paires de séquences allèliques pour tous ses gènes moléculaires on ne peut imaginer que toutes les cellules possèdent les mêmes allèles, les mêmes normes de réactions et que par là on puisse définir un phénotype moléculaire au niveau de l'organisme. La dichotomie génétique + épigénétique* n'est pas non plus satisfaisante puisqu'elle ne fait pas sortir du cadre de l'interprétation de la biologie moléculaire. |
|||
|
|
|
|
|
||||
|
|
|
|
|||||
|
|
|||||||