À propos
des travaux de F. Griffith (1928)
et d'Avery, McLeod, McCarthy (1944) sur les pneumocoques
Pourquoi un tel
engouement ? À la recherche de la vérité
|
-
|
retour cours
de premièreS,
cours
de terminale spécialité
|
-
|
|
Avertissement
cette page n'est pas un vrai travail d'histoire
des sciences.
C'est une lecture des articles originaux avec un effort
d'analyse du paradigme dominant actuel et de sa
méthode. Je reprend à la source deux des
articles cités par les tenants de ce paradigme afin
d'essayer de voir pourquoi ils leur plaisent tant et si on
ne peut pas essayer d'y voir aussi autre chose. Bref il
s'agit de mieux comprendre les armes de ses "adversaires"
dans la controverse scientifique entre paradigme
moléculariste et génétique et
l'ouverture vers un niveau de compréhension
supérieur. Mais je me place résolument CONTRE
les présentations falsifiées si courantes
(voir ci-dessous).
|
-
|
Sources
article original en anglais http://0-profiles.nlm.nih.gov.portia.nesl.edu/CC/A/A/O/D/_/ccaaod.pdf
E.U. articles "streptocoques", "pneumocoque"
Microbiologie, Prescott, Harvey, Klein, De Boeck, 1995, pp
455-457, 747
(pour la replacer dans son contexte d'histoire de la
génétique voir page
sur la
génétique
où une présentation trop simplifiée et
conforme au paradigme actuel est faite)
|
|
|
|
|
|
Plan
de cette page
|
|
- 1.
Données actuelles sur les pneumonies
à Streptococcus
pneumoniae
- 1.1 Une
infection endogène
- 1.2 La
virulence est due à des polyosides
solubles organisés en capsule qui
diminuent fortement la phagocytose par les
macrophages
- 1.3
Inoculés in vivo en même
temps que des anticorps dirigés contre
eux ou vieillis in vitro en
présence de sérums les souches
S de Streptoccocus pneumoniae peuvent
perdre leur capsule (souches R)
- 1.4 Les
souches de S. pneumoniae peuvent
être différenciés en
types en fonction de substances qu'elles
sécrètent ou de composants de
la capsule ou de la paroi
- 2.
Données historiques
- 3.
Conclusion
|
|
|
|
|
|
-
|
1.
Données actuelles sur les pneumonies à
Streptococcus pneumoniae
|
-
|
|
-
|
1.1 Une infection
endogène
|
|
-
|
Le pneumocoque est
facilement identifié par sa morphologie bien
particulière et par son pouvoir pathogène pour
la souris blanche (septicémie mortelle en 24 heures,
après inoculation
intrapéritonéale).
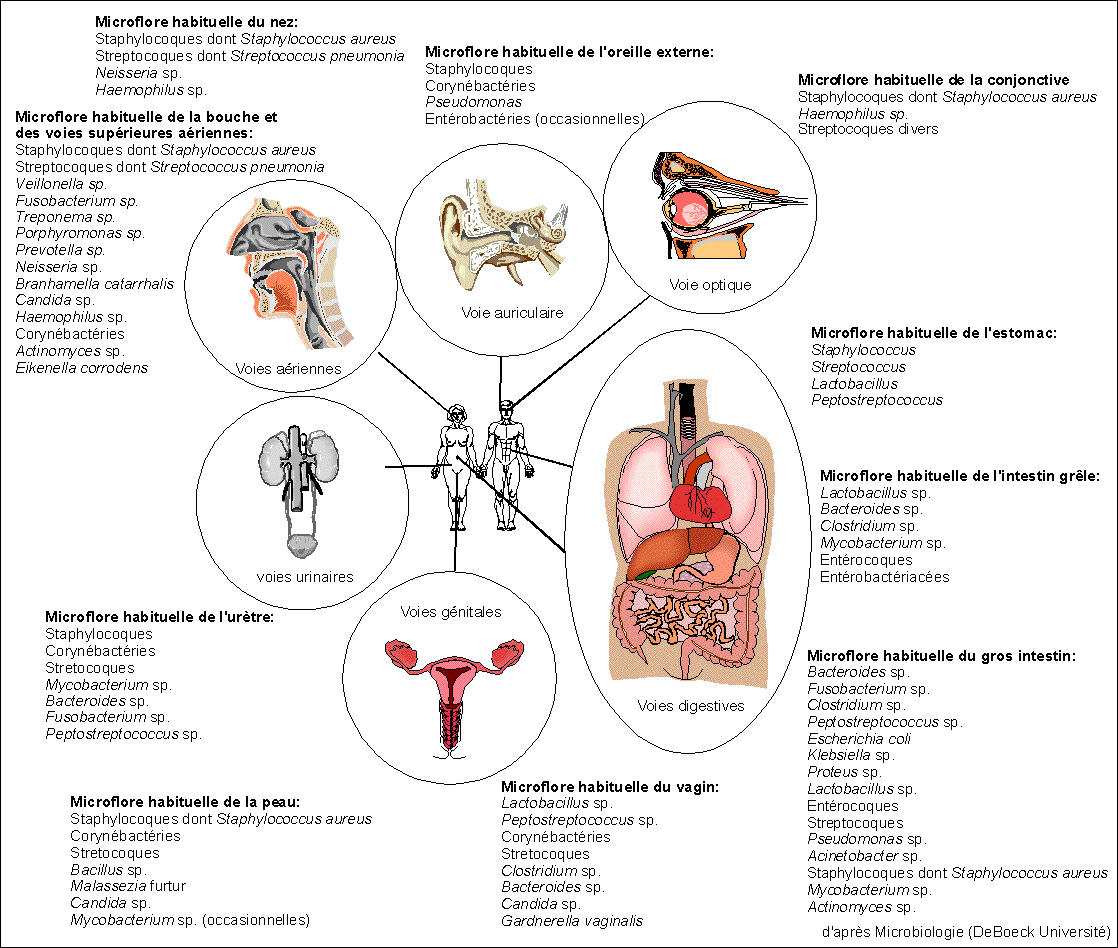
C'est un commensal habituel de la bouche et du rhinopharynx,
mais il peut devenir virulent sous l'influence de divers
facteurs, en particulier une déficience de
l'état général; il donne surtout des
atteintes respiratoires. La maladie habituelle est la
pneumonie franche lobaire aiguë (on le retrouve alors
dans les crachats), mais il est aussi responsable de
sinusites, otites, angines, de pleurésies purulentes,
de méningites purulentes, d'arthrites
suppurées, de péritonites.
Le pneumocoque est très sensible aux sulfamides et
aux antibiotiques, pénicilline surtout. Cette
thérapeutique permet, même lorsqu'il s'agit de
méningites à pneumocoques, autrefois
mortelles, d'obtenir des résultats satisfaisants.
(E.U. article "pneumocoque")
|
La pneumonie streptococcique
est maintenant considérée comme une infection
endogène due au développement anormal dans les
poumons d'un hôte habituel de la bouche et des voies
aériennes (voir
figure de l'ancien
cours
d'immunologie de
terminale). La maladie n'apparaît donc que chez des
individus prédisposés par une fatigue, une
infection virale, une blessure, l'alcoolisme ou le
diabète. 60 à 80% de toutes les maladies
respiratoires reconnues comme des pneumonies sont dues
à S. pneumoniae.
|
La pneumonie lobaire est une
infection des lobes pulmonaires du à
différents agents (bactéries, virus et
même mycoplasmes...). Le Streptococcus
pneumoniae est l'agent principal de la pneumonie lobaire
franche (forte fièvre brutale avec frissons,
oppression respiratoire et ventilation
accélérée, douleurs pectorales,
expectorations sanguinolentes, bruits
caractéristiques entendus au stéthoscope...).
Mais cette bactérie présente une attirance
particulière pour les tissus nourriciers:
plèvres, méninges, péricarde et cause
aussi des méningites, otites arthrites... Les
pneumonies se traitent fort bien par des antibiotiques
(pénicilline G, céfotaxime, ofloxacine et
ceftriaxone).
Les bactéries pathogènes
pénètrent dans les cellules, s'y multiplient
et provoquent leur éclatement (lyse) et donc une
destruction importante du tissu pulmonaire, notamment au
niveau des plèvres, ce qui empêche une
ventilation correcte.
Les crachats de malades contiennent un grand nombre de
bactéries vivantes.
|
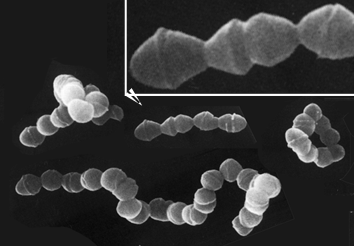
chaînettes de Streptococcus
pneumoniæ (MEB x 33.000, encart x
4,3)
image d'après http://www.uni-tuebingen.de/modeling/images/strep.gif
probablement extraite de Microbiologie, De Boeck,
1995
|
|
Pour des données médicales voir:
http://www.chu-rouen.fr/ssf/pathol/pneumonieapneumocoques.html
voir par exemple http://www.omedit-centre.fr/fichiers/upload/Fiche%20Infections%20a%20S.pneumoniae.pdf
|
|
1.2 La virulence est due
à des polyosides solubles organisés en capsule
qui diminuent fortement la phagocytose par les
macrophages
|
|
-
-
|
Certaines espèces de
streptocoques possèdent à leur surface une
capsule constituée de macromolécules
polyosidiques couvrant la paroi mais ces molécules
solubles sont aussi libérées dans le milieu.
On parle de capsule lorsque la couche de polyosides est
organisée. Elle peut être colorée pour
l'observation au microscope optique et peut être
retirée par attaque ménagée (les
polyosides qui la composent sont dit solubles).
|
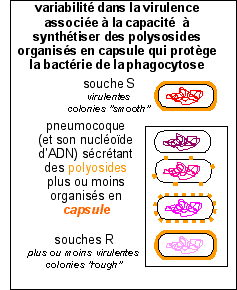
La présence de
capsule confère souvent à la colonie
bactérienne un aspect mucoïde (lisse et
humide: smooth en anglais) sur milieux solides.
Chez S. pneumoniae, la capsule qui
confère également le phénotype
mucoïde aux colonies est le facteur majeur de la
virulence du germe, les souches non capsulées
(colonies rugueuses sur milieux solides) étant
avirulentes. Contrairement à S. pyogenes,
la composition chimique du polyoside capsulaire est
extrêmement polymorphe. Il constitue un marqueur
antigénique et épidémiologique majeur
dont l'identification est à la base de la
sérotypie des souches. On connaît actuellement
quatre-vingt-trois sérotypes distincts
différant par la séquence et
l'enchaînement (polymères linéaires ou
ramifiés) des molécules glucidiques
constitutives (de 2 à 5 oses différents,
rarement plus). Un seul type de polyoside capsulaire est
présent sur une souche donnée. Les polyosides
pneumococciques sont fortement immunogènes et
suscitent des anticorps neutralisants (IgG2 chez l'homme).
Les vaccins acellulaires largement utilisés à
l'heure actuelle sont préparés à partir
d'un mélange de nombreux polyosides différents
en fonction des sérotypes les plus prévalants
dans une aire géographique donnée. Le vaccin
utilisé en Europe et en Amérique du Nord
comporte vingt-trois polyosides.(E.U. article
"Streptocoques")
|
|
|
Le polyoside capsulaire
inhibe la phagocytose (empêche la fixation de la
fraction opsonisante du complément,
voir
cours d'immunologie
cadre sur la réponse à médiation
humorale) en empêchant indirectement la fixation des
anticorps sur les streptocoques, nécessaire à
la phagocytose facilitée
réalisée par les macrophages.
La pathogénèse est due à la
multiplication rapide des bactéries dans les espaces
alvéolaires qui se remplissent de sang et sont le
siège d'une inflammation.
|
|
|
|
|
1.3 Les souches S de
Streptoccocus pneumoniae inoculés in
vivo en même temps que des anticorps
dirigés contre elles ou vieillies in vitro en
présence de sérums peuvent perdre leur capsule
et donner des souches R
|
|
|
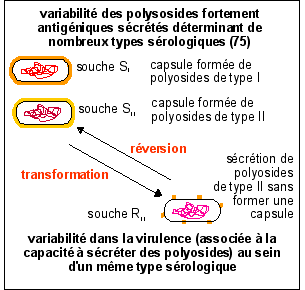
Il ne s'agit pas à
proprement parler de mutation au sens de mutation
génique*
mais simplement de la perte plus ou moins prononcée
de la capacité à synthétiser et/ou
à organiser des polyosides en capsule (qui est
testée par observation microscopique soit par examen
de l'aspect de la colonie en culture). Les résultats
les plus probants obtenus par Griffith ont été
dans un milieu de sang" chocolat" contenant du sérum
de cheval (voir p ).
Les souches R (rough = rugueux en anglais) sont ainsi
désignées à cause de l'aspect de la
colonie au bout de 24 h en culture in
vitro. Les souches R sont habituellement non
virulentes ou moins virulentes. Cependant Griffith obtient
des souches R (groupe IV) directement dans un crachat qui ne
sont ni dépourvues de capsule ni avirulentes. Selon
le type de transformation S -> R obtenu on a plus ou
moins de polyosides sécrétés et une
virulence plus ou moins absente.
La transformation inverse est dite reverse.
|
*depuis
l'époque des travaux historiques du début et
du milieu du XXème siècle et malgré les
innombrables recherches il n'a pas été
isolé de "gène de la capsule" mais de
très nombreux gènes participent à son
élaboration. Il est clair qu'une déficience
structurale de ce type repose probablement sur la
modification de la stabilité d'un réseau
métabolique de bien plus grande dimension que
celui résultant de l'association d'un gène
à une structure.
|
|
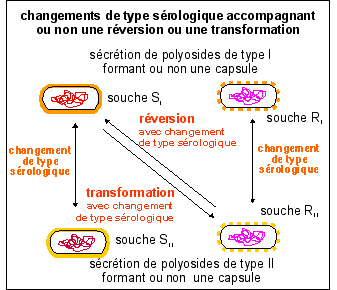
IN
VITRO (en culture) les transformations et
réversions SE FONT (presque) TOUJOURS au sein
d'un MÊME TYPE SÉROLOGIQUE
(sauf
conditions particulières notamment celles
réalisées par Griffith puis
Alloway).
Mais les
changements de types (sans
réversion ni
transformation)
sont très fréquents IN VIVO
(lorsque les pneumocoques sont dans un organisme vivant).
|
|
Il est
possible de transformer une souche peu virulente
(qui
provient elle-même d'une souche virulente par
transformation) en
une souche très virulente d'un AUTRE type par
l'intermédiaire d'un facteur inducteur,
l'acide désoxyribonucléique (Avery, MacLeod,
McCarthy, 1944).
|
|
|
|
1.4 Les souches de
Streptococcus pneumoniae peuvent être
regroupées en types en fonction de substances
qu'elles sécrètent ou de composants de leur
capsule ou de leur paroi
|
|
|
Les médecins
cherchent à connaître le germe présent
dans le cas d'une infection afin de le traiter par un
antibiotique efficace. D'où la
nécessité de la démarche de
reconnaissance.
S. pneumoniae fait partie des S. non
groupables car dépourvus d'antigènes de
groupe définis par des marqueurs antigéniques
de la classification de Rebecca Lancefield (1933) et
extraits de la paroi en laboratoire par de
l'acide chlorhydrique du formamide ou des enzymes
protéolytiques.
On connaît un antigène pariétal R commun
aux formes R et S.
L'antigène pariétal C est un polysaccharide
qu'on retrouve dans les formes R et S. Le sérum des
pneumoniques précipite ce polysaccharide ; cette
précipitation n'est pas due à un anticorps,
mais à une protéine, la protéine C
réactive, qui disparaît au décours de
l'infection.
Les autres antigènes somatiques produits par la
bactérie sont sans grand intérêt
classificatoire ou de reconnaissance: pneumolysine,
hyaluronidase, fibrinolysine, etc.
|
Récemment, des tests
rapides de groupage ont été mis au point
faisant appel à l'agglutination sur lame de la
suspension du streptocoque à identifier en
présence d'anticorps
antipolyosides fixés sur des
particules de latex ou sur la protéine A de
surface des staphylocoques.
En effet les
polyosides solubles de la capsule sont très
antigéniques. Ils sont sécrétés
en permanence et sont aussi libérés dans le
milieu en donnant une masse mucilagineuse qui est un
élément de diagnostic chez les malades.
On connaît 75 types sériques différents
de polyosides. Leur structure a été
étudiée et ils comprennent notamment de
l'acide galacturonique et un sucre aminé pour le type
I , du glucose, de l'acide glucuronique et de l'acide
aldobionique pour le type II.
Ces polyosides sont aussi antigéniques chez la souris
et haptèniques chez le lapin (ce qui signifie qu'un
même polyoside peut être reconnu
différemment par différents anticorps au
niveau de sous-unités antigéniques que l'on
nomme des épitopes ou haptènes). Mis en
présence d'anticorps on observe un gonflement de la
capsule qui peut être observé au
microscope.
|
Certains streptocoques ont
la capacité de sécréter des
substances ayant la propriété de
lyser les érythrocytes de l'homme
et de diverses espèces animales. Cette
propriété, utilisée par Brown (1919)
comme première tentative de classification des
streptocoques, est en effet un test présomptif
important d'identification sur milieu gélosé
contenant du sang de mouton ou de cheval, qui permet de
différencier les streptocoques en trois classes selon
l'aspect de la lyse autour des colonies: streptocoques
ß-hémolytiques (zone large de lyse
franche), ±-hémolytiques (zone étroite de
lyse floue et partielle et coloration verdâtre du
milieu) et non hémolytiques (NH). La typologie
hémolytique des streptocoques est utile pour orienter
le diagnostic mais reste insuffisante car une même
espèce peut comporter des variétés ou
même des souches à pouvoir lytique
différent.
|
|
plan
|
|
-
|
2.
Données historiques
|
-
|
|
|
2.1 -
Extraits de l'article de
Griffith
|
|
Publication de Fred
Griffith (1928; Griffith, F. 1928. The significance
of pneumococcal types, J. Hyg. 27,
113-159) relatant des observations sur des malades atteints
de pneumonie et de très
nombreuses expériences
réalisées entre 1922 et 1928 à partir
de prélèvements faits sur les malades (278
patients, plusieurs prélèvements à des
stades successifs sur chaque malade), de cultures de
bactéries et d'inoculations chez des souris.
L'article original est disponible en ligne à
l'adresse: http://0-profiles.nlm.nih.
gov.portia.nesl.edu /CC/A/A/O/D/_/ccaaod.pdf
Fred.
Griffith
extraite de
http://0-profiles.
nlm.nih.gov. portia.nesl.edu/ CC/A/A/O/D/_/
ccaaod.pdf
|
|
|
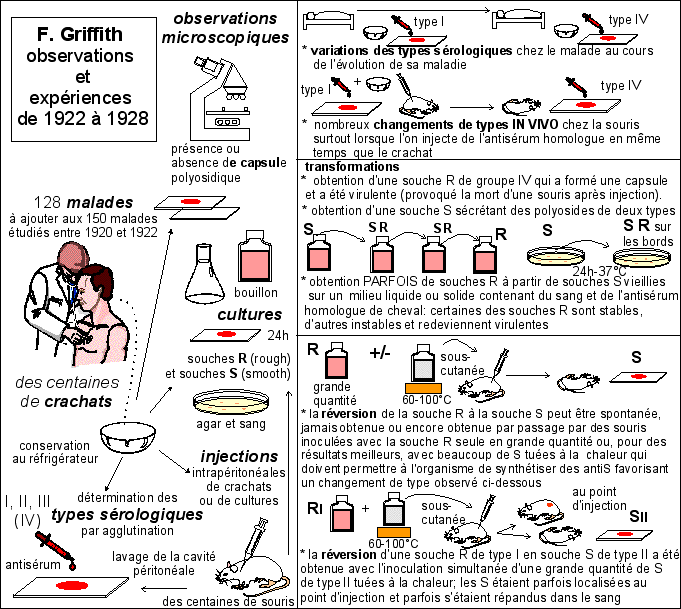
Toutes les
données présentées dans le
résumé de l'article qui est traduit (voir plus
bas) sont résumées dans cette figure
|
|
|
|
|
|
Extraits de l'article
(traduction
personnelle pouvant comporter des erreurs)
|
|
 p 159-160 : The injection of S culture of Type III, heated
at 60" C. for two periods of one hour each, along with
living R strains derived from Type I or Type II results in
the appearance of an S pneumococcus of Type III. This
transformation of type occurs more frequently with the R
form of Type II than with the R form of Type I. The newly
formed strains of Type III sometimes kill the mice from
septicaemia in 7-10 days and at other times are only
discovered at the seat of inoculation when the mouse is
killed, apparently well, 14 days after
inoculation.
p 159-160 : The injection of S culture of Type III, heated
at 60" C. for two periods of one hour each, along with
living R strains derived from Type I or Type II results in
the appearance of an S pneumococcus of Type III. This
transformation of type occurs more frequently with the R
form of Type II than with the R form of Type I. The newly
formed strains of Type III sometimes kill the mice from
septicaemia in 7-10 days and at other times are only
discovered at the seat of inoculation when the mouse is
killed, apparently well, 14 days after
inoculation.
p 162 The
injection of virulent S culture of Type II killed by heat at
60" C together with living R strains of Type I has resulted
in the formation of a virulent S culture of Type II. The
transformation has taken place when the virulent culture has
been heated at 60" C for 2 and 3 hours respectively, but the
positive results were less frequent in the case of the
culture heated for the longer period. Different R strains
appear to vary in their ability to develop into a new S form
under the influence of the heated virulent
culture.
|
 p
159-160 : p
159-160 :
L'injection
de la culture de S du type III, chauffée à
60"C. pendant deux périodes d'une heure chacune, avec
des souches R vivantes dérivées du type I ou
du type II conduit à l'apparition d'un pneumococcus S
de type III. Cette transformation de type se produit plus
fréquemment avec la forme R de type II qu'avec la
forme R de type I. Les nouvelles souches de type III tuent
parfois les souris par septicémie en 7-10 jours et
d'autres fois peuvent uniquement être trouvées
au lieu de l'inoculation quand la souris est tuée,
alors qu'elle est apparemment en bonne santé, 14
jours après l'inoculation.
|
|
p 162
L'injection
de la culture virulente de S de type II tué par la
chaleur à 60"C en même temps que des souches R
de type I a eu comme conséquence la formation d'une
culture virulente de S de type II. La transformation a eu
lieu quand la culture virulente a été
chauffée à 60"C pendant 2 et 3 heures
respectivement, mais les résultats positifs
étaient moins fréquents dans le cas de la
culture chauffée pendant la période la plus
longue. Les différentes souches de R semblent
différer vis-à-vis de leur capacité
à se développer en une nouvelle forme S sous
l'influence de la culture virulente
chauffée.
|
|
|
Résumé présenté en fin d'article
pp 173-175
|
|
IV.
SUMMARY.
1. In the course of the examination of sputum from cases of
lobar pneumonia, observations have been made on the
incidence of the chief types of pneumococci. In the district
from which the material was obtained, there was an apparent
local diminution in the number of cases of lobar pneumonia
due to Type II; the figures were 32.6 per cent of Type II
cases in the period 1920-22: and only 7.4 per cent in the
period 1924-27. The incidence of Type I was approximately
the same in the two periods, the percentages being 30.6 and
34.3.
2. Several different serological varieties of pneumococci
have been obtained from the sputum of each of several cases
of pneumonia examined at various stages of the disease. This
has occurred most frequently in cases of pneumonia due to
Type I, and in two instances four different types of Group
IV were found in addition to the chief types. The recovery
of different types is facilitated by the inoculation of the
sputum (preserved in the refrigerator), together with
protective sera corresponding to the various types in the
order of their appearance.
3. TWO interesting strains of Group IV pneumococci have been
obtained from pneumonic sputum. One was an R strain which
produced typical rough colonies, yet preserved its virulence
for mice and its capacity to form soluble substance. This R
pneumococcus developed a large capsule in the mice, which
died of a chronic type of septicaemia. A strain producing
smooth colonies was obtained from it in the course of a
prolonged series of passage experiments. The second strain,
which was proved not to be a mixture, agglutinated
specifically with the sera of two different types. In the
peritoneal cavity of the mouse the specific soluble
substance of each type was produced.
4. A method of producing the S to R change through ageing of
colonies on chocolate blood medium containing horse serum is
described. After two to three days' incubation small rough
patches appear in the margins of the smooth colonies, and
from these pure R strains can be isolated.
5. It has been shown that the R change is not equally
advanced in the descendants of virulent pneumococci which
have been exposed to the action of homologous immune serum.
Some R strains form traces of soluble substance in the
peritoneal cavity of the mouse; these revert readily to the
virulent S form and, in addition, are able to produce active
immunity. Others show no evidence of S antigen; spontaneous
reversion takes place with difficulty, if at all, and they
are incapable of producing active immunity. The stronger the
immune serum used, the more permanent and complete is the
change to the R form.
6.
Restoration of virulence to an attenuated R strain, with
recovery of the S form of colony and of the original
serological type characters may be obtained by passage
through mice. The change from the R to the S form is
favoured by the inoculation of the R culture in large doses
into the subcutaneous tissues; but the most certain method
of procuring reversion is by the inoculation of the R
culture, subcutaneously into a mouse, together with a large
dose of virulent culture of the same type killed by heat.
Incubation of such a mixture in vitro does not induce
reversion.
7. Reversion of an R strain to its S form may occasionally
be brought about by the simultaneous inoculation of virulent
culture of another type, especially when this has been
heated for only a short period to 60" C., e.g. R Type II to
its S form when inoculated with heated Type I culture.
8. Type I antigen appears to be more sensitive to exposure
to heat than Type II antigen, since the former loses the
power to cause reversion when heated to 80" C, whereas Type
II culture remains effective even after steaming at 100'
C.
9. The antigens of certain Group IV strains appear to be
closely related to that of Type II, and are equally
resistant to heat. Steamed cultures of these Group IV
strains cause the R form derived from Type II to revert to
its S form, while they fail to produce reversion of the R
form derived from Type I.
10. The inoculation into the subcutaneous tissues of mice of
an attenuated R strain derived from one type, together with
a large dose of virulent culture of another type killed by
heating to 60" C., has resulted in the formation of a
virulent S pneumococcus of the same type as that of the
heated culture. The newly formed S strain may remain
localised at the seat of inoculation, or it may disseminate
and cause fatal septicaemia. The S form of Type I has been
produced from the R form of Type II, and the R form of Type
I has been transformed into the S form of Type II. The clear
mutinous colonies of Type III have been derived both from
the R form of Type I and from the R form of Type II, though
they appear to be produced more readily from the latter. The
newly formed strains of Type III have been of relatively low
virulence, and have frequently remained localised at the
subcutaneous seat of inoculation. Virulent strains of Types
I and II have been obtained from an R strain of Group
IV.
11. Heated R cultures injected in large doses, together with
small doses of living R culture have never caused
transformation of type, and only rarely produced a reversion
of the R form of Type II to its virulent S form.
12. The results of the experiments on enhancement of
virulence and on transformation of type are discussed and
their significance in regard to questions of epidemiology is
indicated.
|
IV RÉSUMÉ
1. Au cours
de l'examen du crachat des cas de pneumonie lobaire, des
observations ont été faites sur la
répartition des types principaux de pneumococci. Dans
la zone où le matériel a été
obtenu, il y avait une diminution apparente du nombre de cas
de pneumonies lobaires dues au type II ; les chiffres
étaient de 32,6 pour cent de cas de type II pendant
la période 1920-22 : et seulement 7,4 pour cent pour
la période 1924-27. L'incidence du type I
était approximativement la même pour les deux
périodes, les pourcentages étant 30,6 et
34,3.
2. Plusieurs
variétés sérologiques
différentes de pneumococci ont été
obtenues à partir du crachat de chacun des
différents cas de pneumonie examinés à
diverses étapes de la maladie. Ceci s'est produit le
plus fréquemment dans les cas de pneumonie due au
type I, et, à deux reprises, quatre types
différents de groupe IV ont été
trouvés en plus des types principaux. L'obtention de
types différents est facilitée par
l'inoculation du crachat (préservé dans le
réfrigérateur), en même temps que celle
des sérums protecteurs correspondants aux divers
types , dans l'ordre de leur apparition.
3. DEUX
souches intéressantes de pneumococci du groupe IV ont
été obtenues à partir du crachat
pneumonique.
- L'une était une souche de R qui a produit les
colonies rugueuses typiques, qui pourtant conservaient sa
virulence vis-à-vis des souris et sa capacité
à former la substance soluble. Ce pneumococcus R a
développé une grande capsule chez les souris,
qui sont mortes d'un type chronique de septicémie.
Une souche produisant les colonies lisses a
été obtenue à partir d'elle au cours
d'une série prolongée d'expériences de
repiquage (?).
- La deuxième souche, que l'on a prouvé ne pas
être un mélange, a été
agglutinée spécifiquement avec les
sérums de deux types différents. Dans la
cavité péritonéale de la souris la
substance soluble spécifique de chaque type a
été produite.
4. Une
méthode pour provoquer le changement de S à R
par le vieillissement des colonies sur un milieu de sang"
chocolat" contenant du sérum de cheval est
décrite. Après une incubation de deux à
trois jours de petites taches rugueuses apparaissent sur les
bords des colonies lisses, à partir desquelles
peuvent être isolées de pures souches
R.
|
|
5. On a
montré que le changement chez R n'est pas
développé de la même façon chez
les descendants de pneumococci virulents qui ont
été exposés à l'action d'un
sérum immunisé homologue. Certaines souches R
produisent des traces de substance soluble dans la
cavité péritonéale de la souris ;
celles-ci retournent aisément à la forme
virulente de S et, en outre, sont capables de provoquer une
réponse immunitaire. D'autres ne montrent aucune
trace d'antigène S ; la réversion
spontanée a lieu avec difficulté, le cas
échéant, et ils sont incapables de provoquer
une réponse immunitaire. Plus le sérum
immunisé utilisé est concentré, plus le
changement en forme R est persistant et complet.
6. La
restauration de la virulence à une souche
atténuée R, avec le retour à une forme
S de colonie et aux caractères du type
sérologique original peut être obtenue par le
passage par des souris. Le changement de la forme R à
la forme S est favorisé par l'inoculation de la
culture R à doses élevées dans les
tissus sous-cutanés ; mais la méthode la plus
certaine pour obtenir la réversion est l'inoculation
sous-cutanée de la culture R à une souris
simultanément à une grande dose de culture
virulente du même type tué par la chaleur.
L'incubation d'un tel mélange in vitro ne provoque
pas la réversion.
7. La
réversion d'une souche R à sa forme S peut
parfois être provoquée par l'inoculation
simultanée d'une culture virulente d'un autre type,
particulièrement quand celle-ci n'a été
chauffée que durant une période courte
à 60"C., par exemple une culture de type II R
retourne à sa forme S une fois inoculée avec
une culture de type I chauffée.
8. L'antigène de type I semble être plus
sensible à l'exposition à la chaleur que
l'antigène de type II, puisque le premier perd la
capacité à provoquer la réversion une
fois chauffé à 80"C, alors que
l'antigène de la culture de type II demeure efficace
même après le passage à la vapeur
à 100'C.
9. Les
antigènes de certaines souches du groupe IV semblent
être étroitement liés à ceux du
type II et sont de la même façon
résistants à la chaleur. Les cultures de ces
souches du groupe IV passées à la vapeur
provoquent la réversion de la forme de R
dérivée du type II à sa forme S, alors
qu'elles ne produisent pas la réversion de la forme R
dérivée du type I.
10.
L'inoculation dans les tissus sous-cutanés des souris
d'une souche atténuée R dérivée
d'un type, ainsi qu'une grande dose de culture virulente
d'un autre type tué par la chauffage à 60"C, a
eu comme conséquence la formation d'un pneumococcus
virulent S du même type que celui de la culture
chauffée. La souche S nouvellement formée peut
demeurer localisée au siège de l'inoculation,
ou elle peut être disséminée et causer
une septicémie mortelle. La forme S de type I a
été produite à partir de la forme de R
de type II, et la forme de R de type I a été
transformée en forme de S de type II. Les colonies
mutées claires de type III ont été
dérivées de la forme R de type I et de la
forme R de type II, bien qu'elles semblent être
produites plus aisément à partir de cette
dernière. Les souches nouvellement formées de
type III ont une virulence relativement basse et sont
fréquemment demeurées localisées au
lieu de l'inoculation sous-cutanée. Des souches
virulentes des types I et II ont été obtenues
à partir d'une souche R de groupe IV.
11. Les
cultures R chauffées injectées à doses
élevées, en même temps que des doses
faibles de culture R vivantes n'ont jamais causé la
transformation du type, et ont seulement parfois
provoqué une réversion de la forme R de type
II à sa forme virulente S.
12. Les
résultats des expériences sur l'augmentation
de la virulence et sur la transformation des types sont
discutés et leur signification est expliquée
dans le cadre des problèmes
épidémiologiques.
|
|
plan
|
|
-
|
|
2.2 -
Comment présenter les résultats de Griffith de
façon vraie ?
|
-
|
|
Aucun
chercheur ne présente ses données de
façon "neutre". La neutralité signifierait en
fait l'absence de but. Or le chercheur veut prouver quelque
chose, même si cette chose lui est apparue au cours
d'une observation ou d'une expérience. C'est à
l'historien des sciences d'essayer de retrouver ces buts qui
souvent se sont dilués lorsque l'on a fait un usage
pédagogique de la relation de
l'expérience.
Ainsi l'usage que font les enseignants actuellement d'une
expérience dite "historique" relève donc de la
compréhension des buts des hommes de science actuels.
C'est cela que je veux démêler. C'est en cela
que l'on peut essayer de chercher une
vérité.
|
Il n'est pas rare de trouver la relation
de ces expériences de façon FAUSSÉE,
parfois sans que le nom de Griffith soit cité. Par
exemple, les figures 2.1 et 2.2 de Biologie
moléculaire du gène, Watson et al., 2009,
Pearson Education. Un gène codant pour la capsule
(cap) est INVENTÉ et présenté
sous deux allèles R et S... ceci est une
mystification grave dans un ouvrage de tel niveau
scientifique. Si la recombinaison est une hypothèse
qui est toujours valable, l'idée qu'il n'y ait qu'un
gène impliqué, pire encore, que ce soit "le
gène de la capsule" (légende fig 2.1) est une
mystification. Même dans le cas de l'hypothèse
génétique, de nombreux gènes peuvent
être impliqués dans un tel phénotype.
Des réseaux génétiques redondants
peuvent être aussi mis à contribution. Une
explication scolaire ne justifie pas l'inexactitude. Cela
prouve, à mon avis, l'intérêt de
présentations vraiment
historiques.
|
|
La capacité à
sécréter des polyosides organisés en
capsule (qui cause la virulence) est
SÉLECTIONNÉE et non comprise. La perte ou le
retour à cette capacité est
réalisée À L'AVEUGLE dans un
organisme vivant tout à fait étranger
à la bactérie (la souris), quoiqu'étant
un hôte habituel, et enfin à une échelle
totalement différente (quels sont les organes et les
cellules immunitaires du mammifère impliqués
?).
La causalité est encore une fois SUPPOSÉE.
Texte proposé
|
Voilà
peut-être ce qui a tant plu à ceux que l'on
pourrait qualifier de "positivistes darwiniens" (sans
vouloir les enfermer dans des positions philosophiques trop
tranchées, certainement inexactes
historiquement).
|
|
Griffith
(1922-1928) a exploré très scientifiquement et
honnêtement la variabilité de la formation de
la capsule protectrice des pneumocoques en culture et in
vivo chez la souris. Parmi d'autres hypothèses et de
très nombreux autres cas, il propose la
présence d'un facteur chimique inducteur
spécifique de certaines réversions (retour
à la capacité à synthétiser une
capsule après la perte de celle-ci) avec changement
de type sérologique.
|
L'hypothèse testée est la
présence d'un facteur chimique inducteur.
Les expériences confirment la validité de
cette hypothèse dans certains cas.
Avec le recul pris en ce début de
XXIème siècle vis-à-vis de ces
résultats, les développements de ces
expériences devraient porter principalement sur
l'évaluation d'une autre méthode que la
sélection qui a dominé le
XXème siècle (et domine encore la biologie
moléculaire) ; l'exploration de réseaux
métaboliques supposés plus ou moins stables en
fonction de l'environnement implique une méthode qui
tienne compte de tous les individus et notamment de la
variabilité individuelle
(voir Amzallag qui vulgarise ces
notions essentielles mais combien complexes: Gérard
Nissim Amzallag , Du sens de la variabilité in
Génétiquement
indéterminé - le vivant
auto-organisé,
éditions Quæ (Cemagref, Cirad, Ifremer, Inra),
2007).
|
|
plan
|
|
-
|
|
2.3 -
Étapes suivantes: induction de la transformation
reverse R -> S avec changement de type sérologique
in
vitro
|
-
|
|
D'une façon plus
générale voici une source appréciable
de documents:
http://osulibrary.
oregonstate.edu/ specialcollections/ coll/pauling/dna/
chronolist.html
http://osulibrary.
oregonstate.edu/ specialcollections/ coll/pauling/dna/
papers/avery.html
http://profiles.nlm.
nih.gov/CC/Views/ Exhibit/documents/
shifting.html
puis http://profiles.
nlm.nih.gov/CC/ Views/Exhibit/
documents/dna.html(
|
Les travaux de
Griffith vont donner lieu à de très nombreuses
recherches notamment dans le centre de Rockfeller où
travaillent les auteurs ci-après.
Les chercheurs sont à la recherche d'un principe
transformant CHIMIQUE qui aurait agit dans la
dernière série d'expériences de
Griffith.. Alors que dans sa publication Griffith affirme
clairement qu'il pense que les antigènes de S
injectés en même temps servent avant tout
à stimuler le système immunitaire de la souris
qui va produire des anticorps antiS qui vont favoriser la
conversion de type comme il l'avait déjà
maintes fois observé dans les expériences
précédentes (voir
ci-contre ->).
Mais ici la voie suivie est donc celle d'une transformation
à partir d'un facteur externe en mettant de
côté nombre d'autres hypothèses comme
par exemple des interactions directes entre les cellules
bactériennes VIVANTES. Une vision de chimistes et non
de biologiste.
|
|
pp163-164: « The heated R culture, although the
doses were very large, viz. the deposit of 170 c.c. of
broth, exerted no effect on the living R strains either in
the direction of reversion or transformation of type. This
is consistent with the view that in both cases the result
depends on the presence of S antigen of which there are only
traces in an R strain.».
" les cultures R chauffées, même avec en grande
quantité et additionnées de 170 cc de
bouillon, n'ont aucun effet sur les souches R vivantes, que
ce soit pour la réversion ou pour le changement de
type. Ceci est en accord avec l'idée selon laquelle,
ces deux cas nécessitent la présence de
l'antigène S, qui n'est présent qu'à
l'état de traces dans la souche R".
|
|
a. En présence de
cellules sanguines
|
|
|
|
|
The transformation of
pneumococcal types in
vitro, Dawson,
M.H., and Sia, R.H.P., J. Exp. Med., 1931, 54,
681
Ce papier peut
être obtenu en texte intégral à
l'adresse: http://profiles.nlm.nih.
gov/CC/A/A/B/Q/_/ccaabq.pdf
|
|
|
|
|
Mais ce
n'est qu'un petit papier de 3 pages qui ne décrit
absolument pas les résultats avec la minutie
de Griffith. L'origine des souches, leur âge, les
conditions de culture, le nombre d'essais... bref ce n'est
pas une publication scientifique; en tout cas elle est
inexploitable.
|
"Le procédé consiste dans l'ensemencement
d'une culture de R en quantité minimale dans une
suspension d'organismes S de type hétérologue,
tuées par chauffage. Certaines conditions, bien que
non absolument nécessaires, facilitent apparemment le
processus de transformation. Quelques-unes de ces conditions
sont : (1) la quantité de l'inoculum de R mis en
présence, (2) l'incubation des cultures pendant un
plus long temps que la période conventionnelle, (3)
l'addition d'un peu (10%) de sérum anti-R, (4)
l'addition d'un peu de bouillon de sang. Le degré de
chaleur auquel les organismes ont été
exposés affecte également
matériellement les résultats. Les suspensions
d'organismes chauffés pendant des périodes
aussi longues que quatre heures à 60°C. ont
été efficaces. De même les suspensions
d'organismes chauffés pendant pendant 15 minutes
à 80°C. ont été efficaces.
Cependant, les organismes chauffés pendant pendant 15
minutes à 100°C. ont perdu la capacité
d'induire la transformation."
|
|
« The procedure consists in seeding minimal amounts of
an R culture into a suspension of S organisms of
heterologous type, killed by heating. Certain conditions,
while not absolutely necessary, apparently facilitate the
transformation process. Some of these conditions are : ( 1)
The amount of the R inoculum introduced, (2) the incubation
of the cultures for a longer time than the conventional
period, (3) the addition of a small amount (10%) of anti-R
serum, (4) the addition of a small amount of blood-broth.
The degree of heat to which the organisms have been exposed
also materially affects the results. Suspensions of
organisms heated for periods as long as four hours at
60°C. have been effective. Likewise suspensions of
organisms heated for 15 minutes at 80°C. have been
effective. However, organisms heated for 15 minutes at
100°C. have lost the capacity for inducing the
transformation.»
|
|
plan
b. En absence de cellules
sanguines
|
|
|
|
Alloway Lionel.J. The
Transformation In Vitro of R Pneumococci into S Forms of
Different Specific Types by the Use of Filtered Pneumococcus
Extracts. Journal of Experimental Medicine
55, 1 (1 January 1932): 91-99
http://profiles.
nlm.nih.gov/ CC/A/A/A/I/_/ccaaai.pdf
L'élément
marquant est la nécessité absolu de
sérum anti-R.
|
|
|
|
c. Avec des
extraits les plus purs possibles ne contenant ni
protéines actives, ni polyosides antigéniques,
ni acides ribonucléiques actifs mais uniquement des
acides désoxyribonucléiques actifs
(article
de Avery, McLeod et
McCarthy)
|
|
|
|
Avery Oswald T.,
MacLeod Colin M., and McCarty Maclyn.
Studies on the Chemical Nature of the Substance
Inducing Transformation of Pneumococcal Types.
Journal of Experimental Medicine 79, 2 (1
February 1944): 137-158
L'article réimprimé à l'occasion
d'un anniversaire est disponible en pdf à l'adresse:
http://profiles.nlm.
nih.gov/CC/A/A/ B/Y/_/ccaaby.pdf
La
présentation que j'en ai fait dans ma
page
de
génétique
était fausse, je l'ai
supprimée.
|
Je rappelle
qu'il faut toujours garder à l'esprit que ce que l'on
obtient est la transformation réverse
d'une souche R (provenant elle-même d'une
souche S) en S avec changement de type
sérologique (II -> III). Il ne s'agit pas
à proprement parler d'une transformation
génétique mais bien d'une réversion
biologique induite. L'interprétation
génétique ne prend en compte qu'un des aspects
de la transformation.
|
|
Le papier présente
une description méticuleuse des protocoles.
La souche R (R36A) est obtenue à partir d'une souche
SII repiquée 36 fois dans des bouillons contenant du
sérum de lapin anti-pneumocoque, et qui donne 3
autres souches R mais seule la souche nommée R36A est
capable de transformation.
Pour être transformées les souches R36A sont
ensuite cultivées pendant 16 à 18h à
température ordinaire dans de grands volume (50-75L)
dans un système réactif composé
de bouillons de culture (bouillon de foie de bœuf
passé sur charbon de bois, additionné de 1% de
neopeptone et dont le pH est ajusté à 7,6-1,8)
additionnés NÉCESSAIREMENT de fluide
sérique (human pleural or ascitic fluid), en
présence du principe transformant +/- purifié.
La composition en anticorps antiR du sérum ne semble
pas être le seul point déterminant son
activité; de plus il a été
trouvé que de nombreux sérums tout comme les
lysats de colonies bactériennes possèdaient
des enzymes capable d'inactiver le principe transformant.
|
|
On notera qu'aucune
réversion de la souche R36A vers un type SII n'a
jamais été observé spontanément
et les auteurs considèrent que le mécanisme en
est très différent que la transformation sous
l'action du principe transformant doit probablement se faire
nécessairement pendant une phase de croissance
(divisions cellulaires) dans le bouillon de culture et en
présence du sérum.
|
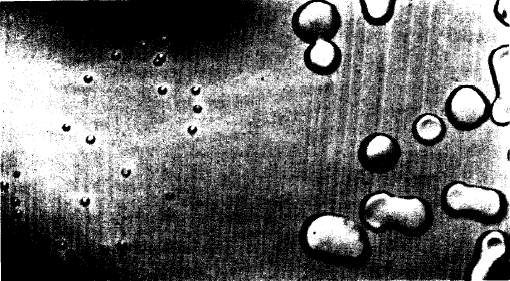
The photograph was made by Mr. Joseph B. Haulenbeek. (in
ttp://profiles.nlm.nih.
gov/CC/A/ A/B/Y/_/
ccaaby.pdf)
|
|
À
gauche: colonies (R36A) de la souche R
dérivée de pneumocoque de type II.
Cultivées sur agar et sang à partir d'une
culture ayant poussé sur du bouillon
additionné de sérum et en absence de la
susbtance transformante (grossissement x
3,5)
FIG. 1. Colonies of the R variant (R36A) derived from
Pneumococcus Type II. Plated on blood agar from a culture
grown in serum broth in the absence of the transforming
substance. X 3.5.
FIG. 2.
Colonies on blood agar of the same cells after induction of
transformation during growth in the same medium with the
addition of active transforming principle isolated from Type
III pneumococci. The smooth, glistening, mucoid colonies
shown are characteristic of Pneumococcus Type III and
readily distinguishable from the small, rough colonies of
the parent R strain illustrated in Fig. 1. X3.5.
|
|
À
droite : colonies sur agar et sang des mêmes
cellules après induction de la transformation lors de
leur culture en bouillon et sérum à l'aide du
principe actif transformant isolé des pneumocoques de
type III. Les colonies lisses, luisantes et d'aspect
"mucoïde" sont caractéristiques des pneumocoques
de type III et aisément différentiables des
petites et rugeuses colonies de la souche parentale R
illustrée à gauche (grossissement x
3,5).
|
|
|
Quelques points de la
discussion montrant combien l'explication
génétique n'est qu'un des aspects bien
abusivement considéré comme unique dans bien
des présentations actuelles de ces
résultats:
Various hypotheses have been advanced in explanation of the
nature of the changes induced.
- In his original description of the phenomenon Griffith (1)
suggested that the dead bacteria in the inoculum might
furnish some specific protein that serves as a "pabulum" and
enables the R form to manufacture a capsular
carbohydrate.
- More recently the phenomenon has been interpreted from a
genetic point of view (26, 27). The inducing substance has
been likened to a gene, and the capsular antigen which is
produced in response to it has been regarded as a gene
product. In discussing the phenomenon of transformation
Dobzhansky (27) has stated that "If this transformation is
described as a genetic mutation-and it is difficult to avoid
so describing it-we are dealing with authentic cases of
induction of specific mutations by specific treatments. . .
."
- Another interpretation of the phenomenon has been
suggested by Stanley (28) who has drawn the analogy between
the activity of the transforming agent and that of a virus.
On the other hand, Murphy (29) has compared the causative
agents of fowl tumors with the transforming principle of
Pneumococcus. He has suggested that both these groups of
agents be termed "transmissible mutagens" in order to
differentiate them from the virus group.
Whatever
may prove to be the correct interpretation, these
differences in viewpoint indicate the implications of the
phenomenon of transformation in relation to similar problems
in the fields of genetics, virology, and cancer research. It
is, of course, possible that the biological activity of the
substance described is not an inherent property of the
nucleic acid but is due to minute amounts of some other
substance adsorbed to it or so intimately associated with it
as to escape detection. If, however, the biologically active
substance isolated in highly purified form as the sodium
salt of desoxyribonucleic acid actually proves to be the
transforming principle, as the available evidence strongly
suggests, then nucleic acids of this type must be regarded
not merely as structurally important but as functionally
active in determining the biochemical activities and
specific characteristics of pneumococcal cells. Assuming
that the sodium desoxyribonucleate and the active principle
are one and the same substance, then the transformation
described represents a change that is chemically induced and
specifically directed by a known chemical compound. If the
results of the present study on the chemical nature of the
transforming principle are confirmed, then nucleic acids
must be regarded as possessing biological specificity the
chemical basis of which is as yet
undetermined.
|
|
Sommaire
1. Une fraction biologiquement active a été
extraite et hautement purifiée à partir de
pneumocoques de type III; une quantité infime de cet
extrait est capable, dans des conditions de culture
appropriées, d'induire la transformation de variants
R sans capsule de pneumocoques de type II en cellules
complètement encapsulées du même type
que celui des micro-organismes tués à la
chaleur dont le matériel inducteur a
été extrait.
2. Les méthodes d'isolement et de purification du
principe transformant sont décrites.
3. Les résultats obtenus par les analyses chimiques,
enzymatiques et sérologiques, conjointement aux
résultats des études préliminaires par
électrophorèse, ultracentrifugation et
spectroscopie UV, indiquent, dans les limites des
méthodes utilisées, que la fraction active ne
contient aucune protéine détectable, ni lipide
libre, ni polysaccharide sérologiquement actif et
consiste principalement, si ce n'est seulement, en une forme
visqueuse, hautement polymérisée, d'acide
désoxyribonucléique.
4. La preuve est apportée que les altérations
chimiquement induites au niveau de la structure cellulaire
et des fonctions sont reproductibles, spécifiques du
type sérologique et transmissible en série.
Les diverses hypothèses qui ont été
proposées concernant la nature de ces changements
sont étudiées.
|
|
SUMMARY
1. From Type III pneumococci a biologically active fraction
has been isolated in highly purified form which in
exceedingly minute amounts is capable under appropriate
cultural conditions of inducing the transformation of
unencapsulated R variants of Pneumococcus Type II into fully
encapsulated cells of the same specific type as that of the
heat-killed microorganisms from which the inducing material
was recovered.
2. Methods for the isolation and purification of the active
transforming material are described.
3. The data obtained by chemical, enzymatic, and serological
analyses together with the results of preliminary studies by
electrophoresis, ultracentrifugation, and ultraviolet
spectroscopy indicate that, within the limits of the
methods, the active fraction contains no demonstrable
protein, unbound lipid, or serologically reactive
polysaccharide and consists principally, if not solely, of a
highly polymerized, viscous form of desoxyribonucleic
acid.
4. Evidence is presented that the chemically induced
alterations in cellular structure and function are
predictable, type-specific, and transmissible in series. The
various hypotheses that have been advanced concerning the
nature of these changes are reviewed.
|
|
Conclusion
Les preuves apportées confortent l'idée que
l'acide nucléique, du type désoxyribose, est
l'élément fondamental du principe transformant
des pneumocoques de type III.
|
|
CONCLUSION
The evidence presented supports the belief that a nucleic
acid of the desoxyribose type is the fundamental unit of the
transforming principle of Pneumococcus Type III.
|
|
|
|
Ce que l'on peut dire
|
|
Ce que l'on ne peut PAS
dire
|
|
Un extrait
purifié d'ADN d'une souche III a été
inducteur, nécessairement en présence de
sérum, d'une transformation (de type II-> III)
dans une culture.
|
|
L'ADN
étranger pénètre dans le
pneumocoque
|
|
|
L'ADN
étranger est utilisé pour synthétiser
un type de polyosides différent de celui
synthétisé par le pneumocoque
transformé
|
|
|
L'ADN
étranger ne provoque la transformation de
pneumocoques que dans le type sérologique des
cellules dont il provient
|
|
|
La
transformation ne peut pas être induite par d'autres
substances
|
|
Discussion
|
|
* On ne peut vraiment que
faire confiance à des chimistes pour ce qui est de la
rigueur des protocoles d'extraction et de dilution. Vous
admirerez la prudence avec laquelle ils affirment que l'on a
uniquement de l'ADN dans l'extrait de souches SIII
|
The data obtained by chemical, enzymatic, and
serological analyses together with the results of
preliminary studies by electrophoresis,
ultracentrifugation, and ultraviolet spectroscopy
indicate that, within the limits of the
methods, the active fraction contains no
demonstrable protein, unbound lipid, or
serologically reactive polysaccharide and
consists principally, if not solely, of a
highly polymerized, viscous form of
desoxyribonucleic acid.
Les résultats obtenus par les analyses
chimiques, enzymatiques et sérologiques,
conjointement aux résultats des
études préliminaires par
électrophorèse, ultracentrifugation
et spectroscopie UV, indiquent, dans les limites
des méthodes utilisées, que la
fraction active ne contient aucune protéine
détectable, ni lipide libre, ni
polysaccharide sérologiquement actif et
consiste principalement, si ce n'est
seulement, en une forme visqueuse, hautement
polymérisée, d'acide
désoxyribonucléique.
|
|
* une souche R de type II
(possédant donc un métabolisme particulier) a
modifié son métabolisme (une de ses voies
métaboliques que l'on sait maintenant former un
réseau, très souvent redondant avec plusieurs
voies pour des produits similaires...) sous l'action
inductrice d'ADN. Qu'est-ce qui prouve que l'ADN est
utilisé dans cette "nouvelle" voie métabolique
? Quelles sont les voies inductives du métabolisme
?
* sous quelle forme
pénètre l'ADN? (pénètre-t-il
même ?). Quel est le lien métabolique entre
l'ADN et les polyosides de la capsule ? (quelles sont les
enzymes qui interviennent ? Sont-elles différentes
selon les polyosides capsulaires ?.... je précise que
ces questions n'ont ACTUELLEMENT pas de réponse
claire, voir ci-dessus les données
actuelles).
* pourquoi n'a-t-on pas fait
des réversions pour d'autres types
sérologiques I -> III ou IV -> III et ainsi de
suite avec d'autres ADN ?
|
|
* seules les souches
révertantes SIII ont été
recherchées. Des souches SII ont probablement
été obtenues d'après le
texte
|
The capacity of this R strain (R36A) to respond to
a variety of different transforming agents is shown
by the readiness with which it can be transformed
to Types I, III, VI, or XIV, as well as to its
original type (Type II), to which, as pointed out,
it has never spontaneously reverted , p 140
La
capacité de la souche R (R36A) à
répondre à une gamme d'agents
transformants variée est montrée par
la facilité avec laquelle elle peut
être transformée en types I, III, IV
ou XIV, ainsi qu'à l'intérieur de son
type original (II), alors qu'il faut noter qu'elle
n'a jamais opéré de réversion
spontanée vers ce type.
|
C'est en cela que le protocole n'est pas clair. On a
systématiquement recherché les tubes où
la transformation souhaitée avait eu lieu mais on n'a
pas listé les autres transformations ni leurs
conditions d'obtention. Cela provient probablement d'une
certitude exprimée dans le paragraphe suivant:
|
The reversible conversion of S <=> R within
the limits of a single type is quite different from
the transformation of one specific type of
Pneumococcus into another specific type through the
R form. Transformation of types has never
been observed to occur spontaneously and
has been induced experimentally only by the special
techniques outlined earlier in this paper. Under
these conditions, the enzymatic synthesis of a
chemically and immunologically difIerent capsular
polysaccharide is specifically oriented and
selectively determined by the specific type of S
cells used as source of the transforming
agent.
La conversion réversible entre S et R au
sein d'un même type est nettement
différente de la transformation d'un type
spécifique de Pneumococcus en un autre type
spécifique à travers la forme R. Le
changement de type n'a jamais été
observé comme se produisant
spontanément (voir
ci-dessous)
et
n'a été induit
expérimentalement qu'à l'aide des
techniques décrites auparavant dans cet
article. Dans ces conditions, la synthèse
enzymatique d'une capsule de polysaccharide
chimiquement et immunologiquement différente
est spécifiquement induite et
sélectivement causée par le type
sérologique particulier des cellules S
utilisées comme source d'agents
transformants.
|
MAIS Griffith (voir-ci-dessus
) rapporte de
très nombreuses transformations de type in
vivo à moins que l'on puisse considérer
que plusieurs souches coexistent chez un même malade
et viennent successivement à devenir
dominantes.
|
|
plan
|
|
-
|
3.
Conclusion
Ces affirmations, peu
fracassantes, sont très éloignées de ce
que l'on peut trouver habituellement sur internet ou dans
les encyclopédies. J'ai préféré
présenter des affirmations neutres, qui me semblent
exactes, car il est hors de question de traquer toutes les
affirmations erronées. Ce travail est encore une
fois, malgré ses évidentes limitations, la
démonstration de l'intérêt de l'histoire
des sciences.
Ce que l'on peut
affirmer:
|
-
|
|
|
|
Griffith (1922-1928) a
exploré très scientifiquement et
honnêtement la variabilité de la formation de
la capsule protectrice des pneumocoques en culture et in
vivo chez la souris. Parmi d'autres hypothèses et
de très nombreux autres cas, il propose la
présence d'un facteur chimique inducteur
spécifique de certaines réversions (retour
à la capacité à synthétiser une
capsule après la perte de celle-ci) avec changement
de type sérologique.
|
|
Parmi les nombreux
physiciens et chimistes financés notamment par
l'Institut Rockfeller aux États-Unis c'est
l'équipe Avery, Mc Leod et Mc Carty (1944) qui
réussit à sélectionner des
réversions avec changement d'un type
sérologique (II->III) en bouillon de culture
(in vitro), en présence de fluide
sérique humain, sous l'action d'un facteur inducteur
qui est de l'ADN purifié le mieux
possible.
|
|
plan
|

{kind=link}