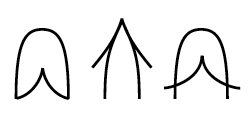retour plan du cours
de 1èreS
Le symbole
![]() , utilisé pour la biologie
théorique,
marque les ouvertures théoriques dans une vision
Thomienne,
ou du moins qui se
croît thomienne.
, utilisé pour la biologie
théorique,
marque les ouvertures théoriques dans une vision
Thomienne,
ou du moins qui se
croît thomienne.
|
Plan
|
... la vie est vraiment l'objet PROPRE de la biologie; toute biologie doit se doter des outils nécessaires pour la comprendre |
||||||||||||||||||||||||||||||||||||||||||||||
|
Compléments
associés (hors de cette page) ;
ces compléments cadrent avec le PROGRAMME DE 1ÈRES : * Le gène moléculaire, unité fonctionnelle synthétique * Les enzymes et la fonction enzymatique * l'ADN lors des cycles cellulaires |
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
Introduction |
Comment j'ai changé mon regard sur le vivant |
||||||||||||||||||||||||||||||||||||||||||||||
|
La vie
n'est pas définie au niveau
moléculaire. |
La vie est vraiment l'objet PROPRE de la biologie; toute biologie doit se doter des outils nécessaires pour la comprendre. Si la biochimie est une voie d'approche de la vie, elle ne peut permettre seule de la comprendre. La vie ne se construit pas à partir du niveau moléculaire mais est un phénomène qui doit être pris comme une donnée. Les mathématiques sont l'outil le plus performant pour comprendre la vie à tous les niveaux. |
Ce cours ne reprend donc pas le plan
classique qui part des molécules pour arriver aux
fonctions car je pense qu'il y a dans cette démarche
une erreur profonde (du discontinu au continu). Ce sont les
dynamiques (les fonctions) qui sont premières. Les
molécules ne sont que des traces de ces
fonctions. |
Cette démarche recoupe la différence entre les observations et expériences in vivo et les expériences in vitro. De très nombreux résultats de la biologie dite moléculaire ont été obtenus in vitro, et les modèles explicatifs proposés ont été, sans état d'âme, extrapolés au vivant. D'où la nécessité, d'une part d'approfondir notre connaissance physique du milieu vivant (si différent du milieu in vitro du tube à essais ; voir page sur la cellule), et d'autre part de toujours laisser la porte ouverte à des explications spécifiques au vivant et donc de proposer de nouveaux outils mathématiques ou physiques pour comprendre la vie. |
||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
La vie s'exprime au niveau des fonctions globales: nutrition, relation, reproduction (voir fiche Qu'est-ce que la vie ?). |
Il est assez conventionnel d'affirmer que les organites et les structures cellulaires (comme une membrane) résultent des dynamiques de cette cellule (sans oublier l'influence de l'environnement). Mais on peut aussi faire cette même remarque au niveau chimique car une molécule n'est pas plus une structure qu'une dynamique (une saillance* au sens de Thom); pour pouvoir réagir avec d'autres substances une substance chimique doit être une forme dynamique et non une forme statique. |
Selon Thom , on ne peut pas faire interagir deux formes statiques sans l'intervention d'une forme dynamique. Le cristal de NaCl se dissous dans l'eau car il présente à son interface avec le solvant une oscillation permanente à l'échelle des liaisons atomiques... |
* le vocabulaire de RenéThom est expliqué dans la page sur les 4 causes. 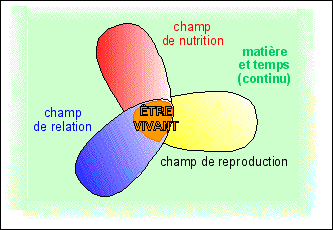
|
||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
 Les 4 causes d'Aristote en SVT |
Toute molécule trouvée naturellement en place dans le vivant ne peut être que la trace ou le support d'une dynamique de vie mais en aucun cas "CAUSER la vie" (on parle de cause au niveau ontologique) ou si vous préférez "DÉTERMINER la vie" (on parle de déterminisme au niveau du phénomène). |
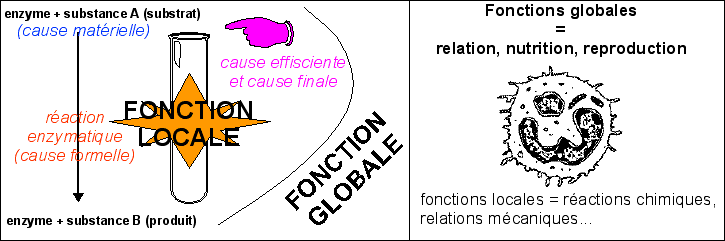
Pour désigner un principe (une structure, une substance) qui cause un phénomène (au sens de la cause matérielle mais aussi formelle et efficiente, voire finale), il faudrait mieux parler de principe déterministe expérimental plutôt que de principe causal car seul le déterminisme (concret) peut être exploré au moyen de l'expérience ... en effet la cause n'est pas toujours suivie d'effet (voir page sur les niveaux d'organisation du vivant et page sur les 4 causes d'Aristote). Une molécule peut avoir une fonction locale, une fonction-outil comme toute molécule chimique dans un tube à essai mais si un biologiste à l'ambition de comprendre le vivant il ne peut considérer sans questionnement que cette fonction locale est responsable d'une fonction globale. Tout au plus cette fonction locale peut-elle résulter de la fonction globale. |
Par contre lorsque l'on injecte une molécule (inséparable de sa dynamique), à un être vivant ou à une cellule, on a une toute autre approche. On tente de modifier chimiquement une dynamique ou même parfois on instaure une dynamique nouvelle. Le rôle de la molécule étrangère, même de l'eau, peut aider à comprendre la dynamique vivante mais là encore cela serait bien naïf d'imaginer que la molécule étrangère va avoir un rôle global qui résultera de la somme des actions locales qu'elle aurait eu en solution en dehors du vivant. |
||||||||||||||||||||||||||||||||||||||||||||
|
Il existe un film extraordinaire de manipulation de descendance en 1956 sur le canard par injection d'ADN (fiche du film : http://www.cerimes.education .fr/index.php?page=fiches &op1=view,1287,4,7,417; lien direct avec le film : (http://www.cerimes.education.fr/index. php ?page=affiche_video,http:// sfrs-streamer.sfrs.fr/ramgen/ media-2/sfrs/real/realvideo/ 4390.rm,,1287,4,7,417) Ces liens ne sont plus valables en 2010. Le CERIMES a, une fois encore, modifié son site....très peu de films gratuits et la plupart sans intérêt. |
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
Une illustration: |
|
||||||||||||||||||||||||||||||||||||||||||||||
|
Annexe
1 : un exemple du jeu entre local et
global Je précise bien que cette manière de voir n'est pas un vitalisme qui s'opposerait à un matérialisme. La matière chimique des scientifiques contient bien plus que la matière (materia prima) des philosophes scolastiques. Dans les deux cas il peut y avoir matérialisme, le moderne étant plus "dialectique". Sans vouloir ici proposer de position philosophique tranchée (même si ma position personnelle est celle d'un catholique forcément ouvert au transcendant), il s'agit simplement de décrire en termes scientifiques les dynamiques associées à cette matière si riche ontologiquement. Je ne fais qu'affirmer que la vie n'est pas un mythe ni un phénomène hors du domaine scientifique (c'est l'opinion opposée qui me paraît être obscurantiste). La vie est un phénomène et que fait la biologie si elle ne s'efforce pas de le comprendre ? |
|||||||||||||||||||||||||||||||||||||||||||||||
|
A 1 - De la genèse à l'abandon d'une vision chimique de la vie |
Il est clair qu'historiquement les conceptions du vivant n'ont jamais cessé d'être diversifiées dans la communauté scientifique. Mais je pense que l'on peut affirmer qu'entre 1900 et 1960 la vision chimique de la vie (que l'on peut qualifier aussi de vision moléculaire) s'est petit-à-petit imposée pour devenir un paradigme dans lequel étaient donc éduquées les jeunes générations. Il y a toujours eu des enseignants et des chercheurs qui ont continué à s'opposer à cette vision réductrice du vivant. Mais les jeunes générations ont mis bien longtemps avant de réfléchir par eux-mêmes et de découvrir ces travaux qui avaient été (volontairement ou involontairement) écartés de l'enseignement qu'ils avaient reçu. Nous sommes, en ce début du XXIème siècle à un tournant et il est probable que l'on puisse d'ici quelques années (...dizaines d'années ?) arriver à changer le contenu de l'enseignement. Voici quelques éléments qui vont dans ce sens, même si, en tant qu'enseignant, ma position est forcément inconfortable, puisque ce que j'enseigne n'est pas conforme au paradigme dominant conforté par le pouvoir politique en place. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1.1 Des chromosomes à l'ADN |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
La structure des chromosomes et une ouverture sur leurs rôles a été vue en seconde : voir fiche chromosomes (6) |
Les niveaux d'organisation sont expliqués dans la première page du cours de 1èreS |
1.1.1 Dès les années
1930-40, les gènes héréditaires ont
été compris par certains comme
contrôlant -par une voie inconnue- les
réactions chimiques de la vie
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Sous l'impulsion américaine, notamment par le biais de la fondation Rockfeller, même en France, une vision chimique de la vie s'est développée entre les années 1910 et 1960 qui a attiré chimistes et physiciens et a donné des applications pharmaceutiques directes notamment lors des deux guerres mondiales puis des applications technologiques encore actuellement en développement. |
Beadle et Tatum ont développé (1941) un protocole bâti sur l'hypothèse de mutations géniques chez des souches auxotrophes de Neurospora sélectionnées sur milieux nutritifs déficients et construit le concept "un gène-une enzyme" (de 1941 à 1945).  Pour une analyse plus complète voir une page complémentaire sur quelques travaux de Beadle et Tatum au sujet du concept un gène - une enzyme. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Lorsque George Beadle et Edward Tatum en 1941 cherchent un matériel expérimental pour établir un lien entre mutants et métabolisme, il choisissent Neurospora crassa, un champignon, et donc un eucaryote, car, malgré le fait qu'Edward Tatum soit microbiologiste, la notion de gène bactérien n'est pas encore admise (voir page sur G. Beadle, E. Tatum et le concept un gène - une enzyme ). Par contre lorsque Tatum travaillera seul après 1946 il retournera aux bactéries pour lesquelles la notion de gène étaient désormais admise. Dès 1941 et puis d'une façon encore plus nette en 1945 les résultats sont exploités avec l'idée que toutes les exigences biochimiques d'un organisme sont susceptibles d'être contrôlées par des gènes dont on peut faire l'exploration au moyen de mutations dont on sélectionne les représentants sur des milieux appropriés. Toute idée d'une variabilité métabolique individuelle, d'une adaptation ou de transformation au cours du développement, de la redondance de mécanismes biochimiques... est non envisagée. On s'enfonce dans ce que l'on pourrait appeler maintenant le réductionnisme biochimique. |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
je renvoie à une
Annexe: |
|
« La sélection a posteriori d'individus modifiés n'est pas seulement la méthode exclusive en biotechnologie, mais également la voie d'investigation privilégiée du vivant. [...] Or, une telle méthodologie implique un lien déterministe (voir ci-dessous) entre le génotype et le phénotype, alors que ce lien prétend être lui-même démontré par l'expérience » (La raison malmenée, Nissim Amzallag, CNRS Éditions, 2002, p 34) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Solution actuelle |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Le déterminisme concret (qui infère la prédictibilité) n'est plus considéré comme évident et l'on reconnaît maintenant en biologie des phénomènes indéterminés (on utilise souvent le mot de hasard pour définir cet indéterminisme mais ce n'est donc pas le sens classique: il s'agit d'un "hasard expérimental" (un phénomène que l'on ne peut pas prévoir expérimentalement) et non ontologique (qui s'oppose à la causalité) ou même mathématique ; voir page sur la science et page sur les 4 causes en SVT). |
En ce qui concerne le déterminisme
génétique ("tel gène conduit
à tel produit dans tel environnement"), les
positions varient selon la philosophie des auteurs. Mais on
peut considérer deux attitudes principales : |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rq: on utilise parfois le terme de postdarwinisme pour désigner cette attitude en se référant à une confusion entre le hasard darwinien et l'indéterminisme moderne (voir page sur l'évolution pour un essai de compréhension de ce qu'est le postdarwinisme). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
La
première partie de la conférence grand public
de Michel Morange: La biologie, de révolutions en
révolutions...
prononcée
en 2006 au congrès IPG (Integrative
Post Genomics)
recoupe
partiellement mes propos: je vous encourage, malgré
la mauvaise qualité de la vidéo, à
suivre celle-ci: |
1.1.2 Du gène chromosomique héréditaire au gène moléculaire : depuis les années 1940, l'ADN, et principalement l'ADN bactérien, a été compris comme un principe causal |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
On est passé progressivement d'un gène eucaryote comme unité de mutation (et de recombinaison) - associé à un emplacement dans un chromosome -, à un gène, unité de fonction, associé à une enzyme, catalysant une (ou deux) réactions chimiques, et présent aussi bien chez les procaryotes que chez les eucaryotes. Ce que l'on peut appeler une vision moléculaire du gène qui a perdu son lien avec le chromosome (et avec la cellule vivante ou même avec l'organisme) pour se limiter à la molécule d'ADN. |
Un exemple paradigmatique: l'ADN comme cause directe de la synthèse des polyosides de la capsule des pneumocoques |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Griffith (1922-1928) a exploré très scientifiquement et avec une grande honnêteté intellectuelle la variabilité de la formation de la capsule protectrice des pneumocoques en culture et in vivo chez la souris. Parmi d'autres hypothèses et de très nombreux autres cas, il propose la présence d'un facteur chimique inducteur spécifique de certaines réversions (retour à la capacité à synthétiser une capsule après la perte de celle-ci) avec changement de type sérologique. |
Parmi les nombreux physiciens et chimistes regroupés notamment à l'Institut Rockfeller aux États-Unis c'est l'équipe Avery, Mc Leod et Mc Carty (1944) qui réussit à sélectionner des réversions avec changement d'un type sérologique (II->III) en bouillon de culture (in vitro), en présence de fluide sérique humain, sous l'action d'un facteur inducteur qui est de l'ADN purifié (le mieux possible).  Les publications originales sont analysées dans une page complémentaire sur la transformation bactérienne des pneumocoques. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Concept historique
de gène moléculaire,
unité
fonctionnelle synthétique de la
cellule. |
La transformation bactérienne, comprise maintenant, de façon quasi générale, comme un échange génétique (qui sera prouvé dans des cas de conjugaison), peut donc avoir une signification bien plus large. Rien ne prouve l'échange d'ADN ici. Il peut s'agir d'une modification d'un réseau métabolique. Ensuite, la transformation, même génétique, peut se faire sans transfert d'ADN. On peut même imaginer que la cellule synthétise alors de nouveaux gènes (à partir d'ARN comme cela est maintenant proposé). |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
La raison malmenée, Nissim Amzallag, CNRS Éditions, 2002, p 35
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Deux exemples pour montrer
l'obsolescence de la liaison
génotype-phénotype enseignée:
l'alcaptonurie,
qui ne peut plus être enseignée comme une
maladie à transmission héréditaire
autosomale récessive et la phénylcétonurie,
plus complexe, mais pour laquelle la liaison
génotype-phénotype reste réfractaire
à l'exploration expérimentale. |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1.2 De la physiologie aux réactions enzymatiques, seules dynamiques du vivant, puis retour |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
1.2.1 Dès 1940, les
protéines, et principalement les enzymes, ont
été comprises comme le principe causal des
dynamiques
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Pour Afinsen la séquence des aa détermine la forme des protéines et la forme détermine la fonction propre |
C'est James Sumner qui en 1926 cristallise l'uréase et émet l'hypothèse que toutes les enzymes sont, comme il vient de le déterminer pour l'uréase, de nature protéique. |
Un quart de siècle plus tard, on considère que les protéines enzymatiques constituent un tout fonctionnel et que le repliement (unfolding en anglais) de la chaîne d'acides aminés qui la compose, stabilisé par les liaisons faibles, est la clé de l'activité catalytique. |
Un article de
Christian B.
Afinsen et col. de 1954 est parfois
cité comme celui où il affirme que la
séquence des acides aminés de la chaîne
polypetidique (pour l'exemple de la ribonucléase
qu'il étudie) détermine
le repliement dans l'espace de la molécule
enzymatique et donc son activité catalytique :
c'est la « native conformation » que prend
spontanément une enzyme dans "le milieu".
Studies on the Gross Structure,
Cross-Linkages, and Terminal Sequences in Ribonuclease,
Anfinsen, Christian B., Redfield, Robert R., Choate, Warren
I., Page, Juanita, Carroll, William R., 1954, Journal of
Biological Chemistry 207, no. 1: 201-210 (http://profiles.nlm.nih.gov/KK/B/B/J/T/_/kkbbjt.pdf).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
En 1954 Afinsen rejoint, grâce à l'aide de la Fondation Rockfeller, le Carlsberg Laboratory à Copenhage où il travaille sur la ribonucléase à l'aide de méthodes physiques modernes. Il publie ses résultats dans un article de 1955 où il considère déjà que seule une petite partie de la protéine est nécessaire à l'activité catalytique (centre réactionnel). Studies on the Structural Basis of Ribonuclease Activity, Anfinsen, Christian, W. F. Harrington, Aase Hvidt, Kaj Linderstrom-Lang, Martin Ottensen, and John Schellman, 1955, Biochimica et Biophysica Acta 17, 141-142 (http://profiles.nlm.nih.gov/KK/B/B/K/W/_/kkbbkw.pdf) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
La mutagenèse dirigée (sélection de mutants présentant des séquences d'enzymes dont des parties sont modifiées à volonté) a permis de voir que les sites supposés actifs n'étaient pas forcément indispensables à l'activité de l'enzyme ou, à l'inverse, que des parties, moins critiques a priori, étaient en fait essentielles à la fonction locale. |
On s'efforce de plus en plus de
comprendre le fonction enzymatique in vivo et non
plus sous forme d'extrait purifié dans des conditions
très éloignées du cytoplasme et les
résultats sont vraiment inespérés
(pour une approche de
l'eau dans la
cellule voir la page
spéciale). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
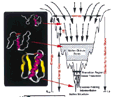 Une illustration d'un paysage énergétique du repliement d'une protéine (Fig 1 de l'article); celui-ci est en deux dimensions. Vous le rapprocherez aisément du paysage phénotypique de Waddington en 3 dimensions (voir cours de seconde sur l'effort pour la première approche sur les fonctions). |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
extrait de René Thom, La notion de préprogramme en biologie, 1984(f5), pp3-4 |
«... on diminuera l'arbitraire de
la construction par une considération
systématique des contraintes qui pèsent
à la fois sur l'organisation spatio-temporelle de
l'être vivant, et sur sa régulation
physiologique. Pratiquement, toutes ces contraintes sont la
conséquence d'un seul principe : le principe de
localité. Il s'exprime ainsi : tout processus interne
à un organisme doit pouvoir se décrire comme
résultant de déterminismes locaux.
Autrement dit, toute action à distance se trouve
proscrite, étant considérée comme
magique. Les entités imaginaires introduites seront
elles-mêmes astreintes à des
déterminismes locaux, soit sur l'espace-temps, soit
dans les espaces abstraits de caractère
sémiotique où elles peuvent se trouver
définies. L'hypothèse de localité se
présente en fait sous deux formes assez distinctes
. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1.2.2 . La théorie de
l'information génétique (ou le couple
ADN-protéines comme principe causal de toute la
machinerie morphogénétique de la vie) est
devenu un paradigme
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Il est difficile de situer historiquement la naissance de ce qui va devenir un paradigme, qui est encore utilisé par certains biologistes actuellement (qui prétendent ne pas avoir besoin des nouveaux développements théoriques, voir l'article Qu'est-ce qu'un gène ? , ainsi ce généticien affirmant «Je ne m'occupe pas vraiment de savoir s'il est impliqué ou non dans la fabrication d'une protéine» ), et qui est enseigné dans le secondaire, mais aussi, souvent, dans le supérieur. |
La théorie de l'information génétique n'est pas une théorie héréditaire, c'est une théorie du vivant (une biologie théorique si l'on va dans le même sens que Michel Morange) qui, selon René Thom, fusionne indûment la notion d'information et de cause (voir cours TS spécialité). La causalité étant supposée évidente au regard d'une matière toute puissante. Elle est indissociable d'un matérialisme (voir par exemple l'article "forme" de Jean Petitot dans l'Encyclopédie Universalis ou la page de ce site sur les modèles). |
Il semble que certains physiciens et chimistes soient pour beaucoup dans l'avènement de ce paradigme (il est intéressant de noter que nombre des chercheurs cités dans cette page ont commencé par des études de physique et de chimie et ne sont venus à la biologie moléculaire que plus tard). Max Delbrück (1906-1981) qui fit partie du groupe du phage, et qui proposa notamment une interprétation quantique des mutations qui représenteraient un saut entre deux états stables du gène, semble avoir influencé Erwin Schrödinger (1887-1961) qui dans son livre Qu'est-ce que la vie ?, publié en 1944, parle déjà de programme génétique (?) en ces termes « ces chromosomes (...) qui contiennent sous la forme d'une espèce de code le modèle intégral du développement futur de l'individu et de son fonctionnement dans l'état adulte ». On peut aussi souligner le rôle de Linus Pauling (1901-1994) qui, en plus de ses découvertes fondamentales en chimie, est celui qui a montré le rôle essentiel des liaisons faibles en biochimie (une bonne part de la compréhension moléculaire que l'on a du vivant actuellement, du moins selon la vision la plus courante, est due à ses idées), mais il est aussi le découvreur des hélice alpha et feuillets bêta des protéines avec Corey en 1951, ou encore celui qui proposé, en 1965, avec Emile Zuckerkandl, le concept d'horloge moléculaire. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
une source très
importante de documentation historique (en anglais) :
http://www.dartmouth.edu/
~bio70/papers.html |
C'est en embryologie que, paradoxalement (Morgan était d'abord un embryologiste avant de se rallier aux théories chromosomiques de l'hérédité), la théorie de l'information génétique va se développer en s'appuyant sur la notion de programme génétique de développement: idée selon laquelle les gènes, et tout particulièrement des gènes majeurs qualifiés de gènes du développement, sont activés séquentiellement dans les différents tissus et organes mis en place au cours de l'embryogenèse (on parle de derépression séquentielle des gènes). |
Scott F. Gilbert, un embryologiste prolifique, auteur du monumental Biologie du développement (De Boeck Université, dernière édition française 2004) - site internet de mise à jour en anglais ; http://8e.devbio.com/), s'est efforcé de mettre à la portée de tous ces données historiques. On peut voir par exemple, en plus du lien ci-dessus (en anglais) :
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
De nombreux mathématiciens et philosophes se sont élevés contre l'emploi abusif des mots information et programme par les biologistes : un texte particulièrement profond présenté par Guiseppe Longo : L'alphabet, la Machine et l'ADN : l'incomplétude causale de la théorie de la programmation en biologie moléculaire, lors du Colloque de Philosophie des sciences : LE LOGIQUE ET LE BIOLOGIQUE, 22 avril 2005, Université Paris 1 (Panthéon-Sorbonne). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Les embryologistes qui ont travaillé en dehors du paradigme moléculaire ont été peu nombreux (et peu médiatisés)
Voir aussi quelques données sur la théorie de R. Chandebois présentées sur une ancienne page. |
L'embryologiste Rosine
Chandebois a présenté une
théorie du
développement
autonome dans plusieurs
ouvrages : |
Plus récemment Vincent
Fleury, un physicien
d'origine, à
présenté des travaux plus
médiatisés (site
personnel): |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Je propose une piste de réflexion et de travail dans la lignée d'une conception du vivant qui place le temps en première ligne avant l'énergie. Avec mes mots, il s'agit encore une fois de s'appuyer sur le temps comme CONTINU. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 (extrait vidéo (.mov) sur le temps biologique -1Mo) Guiseppe LONGO, présentation de son livre avec Francis Bailly, Mathématiques et sciences de la nature. La singularité physique du vivant, Hermann, Paris, juillet 2006 |
Guiseppe LONGO :
complément sur le temps continu dans le cours de terminale sur le temps |
En sciences expérimentales, on
cherche à s'appuyer sur les
causes (page
sur les
sciences).
Si l'on accepte la richesse de la pensée d'Aristote
(voir page
sur les 4 causes en
SVT), on
peut séparer le problème de la
causalité de façon pertinente: |
Dans le mouvement des synthèses de
la cellule vivante comment concilier une dynamique qui va de
l'ADN à la protéine et qui a été
décrite par l'image de l'information (d'une
façon qui semble désormais si
inappropriée) et une dynamique qui va de la cellule
à l'ADN ? La réponse
pourrait résider dans une différence de temps
qui engendrerait deux types de dynamiques
différentes. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 René Thom Photo extraite du CDRom des Œuvres complètes de René Thom publié par l'IHES:http://www.ihes.fr/jsp /site/ Portal.jsp?page_id=217) *Pour le vocabulaire et des généralités sur les modèles thomiens voir une page spéciale. ** chréode, emprunté à Waddington, n'a pas été retenu par l'usage, iI signifie "route obligatoire" suivi par un processus en développement. Le terme de paysage épigénétique, aussi du à Waddington, est maintenant employé pour signifier l'ensemble des trajectoires (qui ne sont pas toutes des chréodes car non structurellement stables). |
1.3 Un essai de formulation des dynamiques cellulaires (partie théorique) Résumé des thèses (René Thom): - plutôt que de chercher des dynamiques extrêmement complexes dans un espace de faible dimension (typiquement euclidien), il est préférable (mais pas du point de vue mathématique où la solution est plus difficile) de chercher des dynamiques simples dans des espaces de grandes dimensions. - René Thom s'est appliqué à décrire les morphologies et les dynamiques les plus simples à partir du vocabulaire ordinaire du fait de la profonde analogie entre le langage et les phénomènes. Avec les mots de René Thom: « 1. Tout objet, ou toute forme physique, peut être représentée par un attracteur* C d'un système dynamique dans un espace M de variables internes*. 2. Un tel objet ne présente de stabilité*, et de ce fait ne peut être aperçu, que si l'attracteur correspondant est structurellement stable*. 3. Toute création ou destruction de formes, toute morphogenèse*, peut être décrite par la disparition des attracteurs représentant les formes initiales et leur remplacement par capture par les attracteurs représentant les formes finales. Ce processus, appelé catastrophe*, peut être décrit sur un espace P de variables externes. 4. Tout processus morphologique structurellement stable est décrit, par une (ou un système de) catastrophe(s) structurellement stable(s) sur P. 5. Tout processus naturel se décompose en îlots structurellement stables les chréodes**. L'ensemble des chréodes et la syntaxe multidimensionnelle qui régit leurs positions respectives constitue un modèle sémantique. 6. Si l'on considère une chréode C comme un mot de ce langage multidimensionnel, la signification de ce mot n'est autre que la topologie globale du (ou des) attracteur(s) associé(s) et celle des catastrophes qu'ils subissent. En particulier, pour un attracteur donné, la signification est définie par la géométrie de son domaine d'existence sur P et la topologie des catastrophes de régulation qui limitent ce domaine. » (stabilité, p 463-464). |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
René Thom, Dynamique globale et morphologie locale chez les êtres vivants, 1981, 11. Preprint IHES, Bures-sur-Yvette, cf. ULP, Sém. Fond. Sc. RCP 443, Strasbourg, 13 novembre 1981, cité dans le CDRom des œuvres complètes :1981f11.pdf ; pp 6-8 C'est moi qui souligne Pour d'autres explications voir la page sur les modèles thomiens. |
« 3. Vers un nouveau paradigme en
Biologie
|
Les variations de type d'attracteur s'appellent en mathématique des bifurcations. Or on démontre, en Dynamique Qualitatitive, qu'au voisinage de chaque attracteur, il existe une sorte d'entropie locale qui croît sur les trajectoires voisines de l'attracteur (ce qu'on appelle une fonction de Lyapunov) ; et lorsqu'un attracteur entre en bifurcation, sa fonction de Lyapunov subit elle aussi une bifurcation dont on pourra supposer qu'elle est analytique. Le modèle de la théorie des catastrophes consiste à distinguer deux types de variables ; des variables internes x, à variation rapide loin de l'équilibre (et des points singuliers du système) ; et des variables u (dites variables externes ou de contrôle) à variation lente. Cette différence de vitesse entre les dynamiques sert en fait à exprimer le rapport entre deux niveaux d'observation : un niveau local fin, paramétré par les x, et un niveau plus grossier - moyenné - paramétré par les u ; la dynamique totale* étant représentée par un potentiel V(x; u) dont on prend le gradient en x. Le même formalisme intervient en Physique dans la théorie des changements de phase (théorie dite de Landau, ou du champ moyen). Ce modèle semble bien adapté à la description de l'épigénèse de l'embryon : les différenciations cellulaires sont alors - comme l'avait suggéré en 1949 Max Delbrück - représentées par des attracteurs du métabolisme cellulaire local, en raison du changement de niveau réalisé de la cellule (niveau fin) au tissu (niveau grossier). On suppose alors que ces potentiels locaux V(x; u), où les variables u contiennent les coordonnées d'espace-temps, ont une signification fonctionnelle liée à la régulation globale de l'organisme ; et que, dans les bifurcations qui les affectent ces significations subissent elles-mêmes des variations compatibles avec la contiguïté sémantique de leurs significations. Tel est le principe, par exemple, d'une théorisation de l'embryologie, dont j'ai esquissé autrefois les premières étapes (pour la gastrulation et la neurulation chez les Vertébrés, notamment). |
Je ne me dissimule pas le
caractère extrêmement incomplet de ces
tentatives : en effet, j'ai été amené
à faire sur les bifurcations des hypothèses
très restrictives (hypothèses de
généricité, lesquelles conduisent
à la stabilité vis-à-vis des
perturbations) ; par ailleurs, le formalisme - qui est
purement qualitatif - ne permet par lui-même aucun
contrôle métrique de la croissance des tissus.
A cet égard, il y aurait lieu d'introduire de
manière plus systématique la notion de «
phase »** : le tissu osseux est -
à peu près - une réalisation de la
phase solide ; le tissu sanguin réalise une phase
liquide ; or chaque phase a son propre « groupe de
symétrie » qui lui aussi agit dans l'espace u
des coordonnées spatio-temporelles ; d'où des
contraintes sur l'application de l'espace (u) dans le «
déploiement » de la bifurcation, application qui
définit la croissance et le mouvement de la phase
correspondante. Les phases locales définies par des
minima des potentiels V(x; u) seraient à la fois en
équilibre « sémantique » - dans un
champ sémantique approprié - et globalement,
des contraintes de conservation des espèces chimiques
et de leur ordre moléculaire imposeraient des
relations entre les modes de croissance de ces phases (noter
que le problème analogue de la croissance d'une phase
en milieu inanimé - par exemple, une cristallisation
- n'est pas résolu).
On pourrait donc qualifier cette
méthode de structuralisme
dynamique ; elle réconcilie
Geoffroy-Saint-Hilaire et Cuvier : Geoffroy, parce que la
succession des différenciations successives dans
l'embryon se réalise de manière sensiblement
isomorphe entre diverses espèces de
Vertébrés ; Cuvier, parce que les potentiels
locaux V(x; u) (ou fonctions de Lyapunov) ont une
signification fonctionnelle liée à la
régulation globale de l'organisme. On a affaire
là à une sorte de vitalisme
méthodologique , nullement incompatible, d'ailleurs,
avec le réductionnisme
physico-chimique.» *pour
des exemples de dynamiques voir une page
complémentaire (la
dernière
partie est en gestation :
toute contribution serait bienvenue) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Un aperçu des
dynamiques enzymatiques a été
réalisé par R. Thom |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Exemples de morphologies dans un espace R4 obtenues à partir de conflits d'attracteurs René Thom, SSM, Fig 13-18 p
451 S'effondrer Ouvrir |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1.4 L'investigation des dynamiques L'expérience ne prouve jamais les dynamiques théoriques proposées comme modèle (a priori): elle permet simplement de voir si le modèle peut ou non s'appliquer dans un cas précis. C'est une justification a posteriori du modèle (voir page sur les modèles thomiens). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
en travaux |
Je recommande ici particulièrement
la réflexion de Gérard Nissim Amzallag;
basée sur ses recherches en botanique elle est
diffusée dans des ouvrages et articles de
vulgarisation accessibles à tous : |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Annexe 1 Un exemple du jeu entre local et global |
|
|||||
|
Le blanc d'œuf (de Poule) composé principalement d'une protéine (l'ovalbumine) soluble dans l'eau et d'eau n'est pas un contenu cellulaire vivant mais un milieu stable, riche en vitamines, équilibré en aa et comportant une fraction glucidique. On la considère comme un aliment (et une boisson nutritive) de réserve pour le développement postembryonnaire. La conformation spatiale de la protéine albumine est celle de pelotes assez lâches emprisonnant de nombreux domaines d'eau liquide. Chauffé le blanc d'œuf coagule ce que l'on interprète comme un déroulement des pelotes d'ovalbumine avec départ d'eau. On considère que cette dénaturation irréversible par chauffage, que l'on observe d'ailleurs pour la plupart des protéines globulaires, est la preuve de ce que la conformation de la protéine est critique pour sa fonction. Bref on fait reposer la fonction sur la structure tridimensionnelle de la protéine. On peut inverser aisément ce raisonnement : la fonction ne vient pas de la molécule mais bien du milieu ; ici la fonction est une fonction physique d'emprisonnement de l'eau. La forme de la protéine reflète la fonction et non l'inverse. Si la protéine est déroulée et a établi de nombreuses liaisons qui conduisent à un état solide du blanc d'œuf cuit c'est bien la trace de la dynamique de chauffage imposée. Lorsque le blanc d'œuf liquide est limpide, c'est bien la trace d'un milieu stable à température ordinaire. Tout le monde est d'accord avec ce raisonnement pour le monde inanimé. |
Mais dès qu'il s'agit d'un
être vivant les choses changent. On considère
que les molécules de la matière vivante
déterminent les fonctions (par exemple les enzymes
participant aux réactions chimiques sont donc les
agents déterminants de ces réactions;
où encore, l'ADN, nécessaire à la
synthèse d'une protéine est donc l'agent
déterminant de la présence de cette
protéine...). De plus, si l'on accorde volontiers une
part importante à l'environnement (telle enzyme a
telle conformation en fonction du pH, de la
température, des interactions avec les autres
molécules...) on fait cependant à tort reposer
les propriétés générales de cet
environnement sur les molécules qui le composent.
Bref on dilue la causalité en multipliant les agents
à l'infini, chacun finalement n'étant plus
cause de rien. |
|||||
|
Les molécules sont
les traces des fonctions locales. Les fonctions locales sont
les traces de la dynamique du vivant (fonctions globales).
|
||||||
|
Remarque supplémentaire pour enfoncer le clou: |
pour les enzymes l'imaginaire est plus fort et l'idée de fonction locale (activité enzymatique) est souvent inconsidérément associée à la fonction globale (synthèse de telle molécule par exemple). Lorsque l'on met dans un tube à essai une enzyme et son substrat la fonction globale est celle que réalise l'expérimentateur en mettant en présence les deux molécules (il est nettement plus clair de parler en termes de causalité efficiente et finale... mais cela demande un bonne connaissance philosophique : voir page sur les 4 causes en SVT). |
C'est en cela que la vision d'une cellule comme un tube a essai est particulièrement fausse. Ce n'est pas en recherchant les interactions entre molécules supposées en place que l'on peut comprendre la cellule vivante. On doit s'intéresser à la mise en place des composants cellulaires et donc des dynamiques, soit héritées soit nouvelles (c'est la dimension temporelle ou évolutive incontournable). On a tendance à oublier qu'in vitro la fonction enzymatique locale dépend de la fonction globale qui prend en compte le travail réalisé par l'expérimentateur lorsqu'il mélange les réactifs dans le milieu dans certaines conditions. |
Pour les 4 cause d'Aristote, voir page annexe). Il n'est pas rare que l'on invoque le hasard pour le vivant en refusant ainsi les notions de causes efficiente et finale. Pour moi c'est une reculade. Cependant il est clair que les fonctions globales ne peuvent pas être explorées expérimentalement; seules les mathématiques pour une formalisation géométrique, semblent pouvoir aujourd'hui y accéder. |
|||
 |
||||||
|
|
||||||