G.
Beadle, E.
Tatum
et le concept
un gène
- une enzyme
|
|
-
|
retour
histoire
de la
génétique
|
-
|
|
Avertissement:
cette page
n'est PAS une
page
d'histoire des
sciences
(discipline
à part
entière,
et je ne suis
pas
historien).
Les
références
et analyses
d'articles
qu'elle
contient ne
servent que de
justification
à mes
propos tenus
dans mon
cours.
|
-Sources
De très
nombreuses
publications
historiques de
Beadle
sont
accessibles
librement sur
PubMed: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&db=PubMed&term=%20Beadle+GW[auth]
notamment
celles
relatives
à ses
travaux avec
Ephrussi ou
Sturtevant sur
la Drosophile.
Beadle,
G.W. & Tatum,
E.L. (1941) Genetic
control of
biochemical
reactions in
Neurospora.
Proc. Natl.
Acad. Sci USA
27, 499-506
(article
complet en pdf
accessible
gratuitement
à
l'adresse
http://www.pubmedcentral.gov/picrender.fcgi?artid=1078370&blobtype=pdf)
Beadle,
G. W.,
1945b Genetics
and metabolism
in Neurospora.
Physiol. Rev.
25:643-663,
non accessible
pour tous sur
internet:
voici le
passage
où il
cite Garrod:  «
These studies
on
tryptophaneless
mutants are an
example of how
genetics can
be of use in
studying
metabolic
processes. It
is almost
exactly
analogous to
the classical
experiments on
alcaptonuria
in man in
which
2,5-dihydroxgphenylacetic
acid is
accumulated
because of a
genetic block
preventing its
further
oxydation
(Garrod, A. E.
Inborn errors
of metabolism.
2nd ed., 216
pp. Oxford
Medicali
Publ., 1923).
This
intermediate
in the
breakdown of
phenylalanine
and tyrosine
would probably
not have been
discovered had
it not been
for
alcaptonuries.
In normal
individuals it
is apparently
a transitory
intermediate
in metabolism
and seldom if
ever
accumulates in
sufficient
amount to
permit, of its
identification.
Similarly in
thryptophane
metabolism ,
ant uranilic
acid is
normally
transitory and
would not be
easily
identified
without using
genetic blocks
to iso1ate it
as a metabolic
step.» «
These studies
on
tryptophaneless
mutants are an
example of how
genetics can
be of use in
studying
metabolic
processes. It
is almost
exactly
analogous to
the classical
experiments on
alcaptonuria
in man in
which
2,5-dihydroxgphenylacetic
acid is
accumulated
because of a
genetic block
preventing its
further
oxydation
(Garrod, A. E.
Inborn errors
of metabolism.
2nd ed., 216
pp. Oxford
Medicali
Publ., 1923).
This
intermediate
in the
breakdown of
phenylalanine
and tyrosine
would probably
not have been
discovered had
it not been
for
alcaptonuries.
In normal
individuals it
is apparently
a transitory
intermediate
in metabolism
and seldom if
ever
accumulates in
sufficient
amount to
permit, of its
identification.
Similarly in
thryptophane
metabolism ,
ant uranilic
acid is
normally
transitory and
would not be
easily
identified
without using
genetic blocks
to iso1ate it
as a metabolic
step.»
L'article
de Garrod
le plus
souvent
cité
(Garrod AE: The
incidence of
alkaptonuria:
a study in
chemical
individuality.
Lancet II
1902; 1616-20)
est disponible
en ligne (http://www.esp.org/foundations/genetics/classical/ag-02.pdf).
Mais on peut
aussi trouver
la
totalité
d'un de ses
ouvrages sur
la librairie
scolaire
virtuelle
(ESP:
Electronic
Scholarly
Publishing: http://www.esp.org/
;
choisir
"digital
books" puis
Garrod,
Archibald.
1923.
Inborn Errors
of Metabolism,
Second
Edition.
London: Henry
Frowde and
Hodder &
Stoughton).
Je
recommande
aussi la
lecture du
discours de
réception
du Prix Nobel
de Beadle en
1958 où
il retrace
toute sa
carrière
précédente:
http://nobelprize.org/nobel_prizes/medicine/laureates/1958/beadle-lecture.pdf
ainsi que
celle de Tatum
(http://nobelprize.org/nobel_prizes/medicine/laureates/1958/tatum-lecture.html)
Pour ceux qui
y ont
accès
il y a des
données
d'histoire des
sciences plus
ou moins
récentes
mais qui sont
en fait des
articles en
hommage
à
Beadle, par
exemple les
interventions
d'Horowitz: le
plus complet
étant
peut
être : A
centennial:
George W.
Beadle,
1903-1989.
Horowitz NH,
Berg P, Singer
M, Lederberg
J, Susman M,
Doebley J,
Crow JF.
Genetics.
2004
Jan;166(1):1-10;
One-gene-one-enzyme:
Remembering
biochemical
genetics,
N. H.
HOROWITZ, Protein
Sci. 1995
4: 1017-1019 .
Voir aussi,
accessible
gratuitement:
The
Centenary of
the One-Gene
One-Enzyme
Hypothesis,
Mark Hickman
and John
Cairns, Genetics,
Vol. 163,
839-841, March
2003 (http://www.genetics.org/cgi/content/full/163/3/839),
un article qui
parle des
travaux
antérieurs
à ceux
de Beadle
(Cuénot,
Bateson,
Garrod...,
mais aussi des
réticences
postérieures
à
1945... rien
n'est dit sur
une remise en
question plus
moderne).
Finalement je
ne suis pas
sûr que
l'on puisse
aisément
tirer grand
chose de ce
matériel
sauf à
être
spécialiste
de l'histoire
de la
génétique.
|
|
On
peut aussi
accéder
intégralement
et librement
aux livres de
Bateson,
Morgan et
Sturtevant
notamment, sur
la librairie
scolaire
virtuelle http://www.esp.org/
(
W. Bateson,
1902, Mendel's
Principles of
Heredity: A
Defence,
London:
Cambridge
University
Press ; W.
Bateson, 1908.
The Methods
and Scope of
Genetics,
London:
Cambridge
University
Press; T.
Morgan, 1919,
The Physical
Basis of
Heredity,
Philadelphia:
J. B.
Lippincott
Company; T.
Morgan, 1928,
The Theory of
the Gene,
Revised and
Enlarged
Edition,
New Haven:
Yale
University
Press; T.
Morgan, H.
Sturtevant, A.
H., Muller, H.
J., and C. B.
Bridges 1915,
The Mechanism
of Mendelian
Heredity.
New York:
Henry Holt and
Company; A.
Sturtevant,
1965, A
History of
Genetics)
|
|
|
|
|

http://clendening.kumc.edu/dc/pc/Beadle.jpg
Biographie
de G. Beadle
George Wells
Beadle est
né en
1903 dans le
Nebraska
où il
fit ses
études
supérieures
de biologie.
En 1927 il est
assistant du
Professeur
Keim de
l'Université
de Cornwell
où il
travaille
jusqu'en 1931,
année
où il
obtient son
Ph.D. degree,
avec une
thèse
sur la génétique
du maïs.
À
partir de
cette
même
année
il travaille
au California
Institute of
Technology en
ne cessant de
s'intéresser
au maïs
indien mais en
commençant
une
collaboration
avec
Dobzhansky,
Emerson et
Sturtevant sur
les
crossing-over
chez la
drosophile. En
1935 il passe
6 mois
à Paris
où il
travaille avec
Ephrussi
à
l'Institut de
Biologie
Physico-chimique
sur la couleur
de l'œil
de drosophile
au cours du
développement.
Il travaille
ensuite avec
Tatum sur la
biochimie
génétique
de Neurospora
crassa. En
1936 il
devient
professeur
assistant de
génétique
à
l'université
d'Harvard. Un
an plus tard
il est
nommé
professeur de
génétique
à
l'université
de Stanford
où il
restera 9 ans,
en travaillant
avec Tatum
(ils recevront
conjointement
le prix Nobel
en 1958). En
1946 il
retourne au
California
Institute of
technology
où il
dirige le
département
de biologie.
En 1961 il est
nommé
chancelier de
l'université
de Chicago
puis, la
même
année
président
de cette
université
(d'après
http://nobelprize.org/nobel_prizes/medicine/laureates/1958/beadle-bio.html).
|

http://nobelprize.org/nobel_prizes/medicine/laureates/1958/tatum.gif
Biographie
de E. Tatum:
Edward Lawrie
Tatum est
né en
1909 dans le
Colarado. Il
fit des
études
de chimie et
de
microbiologie
à
l'université
de Chicago. Il
obtint en 1934
son PhD degree
en biochimie
avec une
thèse
sur la
nutrition et
le
métabolisme
des
bactéries.
En 1936 il
isole la
thiamine
(vitamine B1)
comme
substance de
croissance
nécessaire
à de
nombreuses
cultures
bactériennes
(et fongiques)
lors d'un bref
passage
à
l'université
d'Utrecht
(Hollande). De
1937 à
1945 puis de
1948 il
travaille
à
l'université
de Stanford.
Il collabore
avec Beadle de
1937 à
1946 en
prenant en
charge les
aspects
biochimiques
de
l'étude
de la couleur
de l'œil
de la
drosophile
puis des
mutants de
Neurospora. (http://nobelprize.org/nobel_prizes/medicine/laureates/1958/tatum-bio.html)
|
|
|
|
|
-
|
Beadle
et Tatum ont
développé
un protocole
bâti sur
l'hypothèse
de mutations
géniques
chez des
souches
auxotrophes de
Neurospora
sélectionnées
sur milieux
nutritifs
déficients
et construit
le concept "un
gène-une
enzyme".
|
-
|
Beadle
est un
biologiste et
non un
chimiste mais
son
rattachement
à telle
ou telle
université
n'est pas
forcément
le point
essentiel; ses
publications
de 1941, 1942,
1947 sur Neurospora
crassa
font mention
du soutien
financier de
la Fondation
Rockfeller que
l'on retrouve
partout lors
du
développement
de la biologie
moléculaire.
Il a d'abord
travaillé
sur le
maïs
indien puis
sur la
drosophile et
s'est
intéressé
à la
théorie
chromosomique
"mendélienne"
de
l'hérédité
dans la
lignée
de Morgan (voir
pour cette
théorie
le
cours de
terminale
spécialité).
Il cherche
à
comprendre les
mécanismes
des variations
héréditaires
de
caractères
dont on puisse
faire
l'analyse
biochimique
(il n'est pas
le premier
dans ce
domaine).
Pendant
plusieurs
années
c'est la
couleur de
l'œil de
la drosophile
qui retient
son attention
avec des
expériences
de
transplantation
réalisées
avec Ephrussi.
Lorsqu'il
publie ses
résultats
sur Neurospora
crassa il
n'a pas
cessé
de
s'intéresser
au
crossing-over
chez la
drosophile
qu'il
étudie
en
collaboration
avec
Dobzhansky,
Emerson et
Sturtevant.
C'est dans une
publication de
1945 qu'il
associe ses
résultats
à une
"redécouverte"
des travaux de
Garrod.
Tout
comme on verra
en Terminale
spécialité
avec de Vries,
Bateson et
Fisher pour la
"redécouverte"
des travaux de
Mendel (mythe
du père
fondateur,
voir
page avec
notamment les
travaux
d'histoire des
sciences de
Jan Sapp,
partie 4), on
a
peut-être
"redécouvert"
les travaux de
Garrod pour
asseoir le
concept d'un
gène -
une enzyme.
Garrod est
ainsi souvent
nommé
le
"père
fondateur" de
la
génétique
biochimique.
|
-
|
|
|
|
La
publication de
1941 sur la
biochimie
génétique
de Neurospora
donne le
principe de la
méthode,
des
résultats
préliminaires
et un essai de
justification
du
raisonnement
en terme de
génétique
mendélienne.
|
|
Beadle,
G.W. &
Tatum, E.L.
(1941) Genetic
control of
biochemical
reactions in
Neurospora.
Proc. Natl.
Acad. Sci USA
27, 499-506
|
|
 Modèle
:
Modèle
:
Dans leur
introduction
les auteurs
précisent
que pour
étudier
le lien entre
les
gènes
et les
propriétés
physiologiques
et
biochimiques
des organismes
on
procède
habituellement
en
étudiant
l'hérédité
de
caractères
bien connus
(ils citent
les pigments
de type
anthocyanes
des plantes,
la
fermentation
des sucres par
les
levures...).
Leur approche
est
différente
car ils
considèrent
que l'on peut
créer
par
irradiation
aux rayons X
des souches
ayant des
caractères
héréditaires
stables
NOUVEAUX (en
fait des
déficiences
métaboliques)
qu'ils
considèrent
comme des
mutations
géniques.
(Muller
en 1928 a
étudié
les mutations
produites sous
par
différentes
expositions
aux rayons X
de souches de
drosophile;
mais le texte
est assez
confus
(lecture
à
l'académie
des sciences
et non pas
article au
sens moderne
du terme) et
les
données
expérimentales
rares, ce qui
le rend
difficile
d'accès
à un
non historien
des sciences
... la
publication
(Muller, H.J.
1928. The
production of
mutations by
X-rays. Proc.
Natl. Acad.
Sci. USA, 14:
714-726) est
accessible sur
internet: http://www.pubmedcentral.gov/picrender.fcgi?artid=1085688&blobtype=pdf
)
«
The procedure
is based on
the assumption
that x-ray
treatment will
induce
mutations in
genes
concerned with
the control of
known specific
chemical
reactions.»
Dans leur modèle
un organisme
muté
pour un
gène
spécifique
supposé
comme
étant
associé
à une
substance
chimique
pourra
à
nouveau se
développer
s'il est
cultivé
en
présence
de cette
substance (ce
que l'on
pourra mettre
en
évidence
par des
mesures de
croissance en
fonction de la
concentration
en substance
dans le
milieu) et
à
condition que
cette
substance soit
capable de
passser la
paroi et
atteindre les
cellules pour
y être
utilisée.
Cette
vision est
rapportée
ultérieurement
(dans leur
discours de
prix Nobel par
exemple) par
les auteurs
comme venant
notamment de
Lwoff et
Knight qui
avaient
montré
que certaines
souches
auxotrophes
(nécessitant
une substance
de croissance)
avaient perdu
la
capacité
de
synthétiser
une enzyme
(qui
intervenait
dans la
synthèse
de la
substance de
croissance).
Mais ils
précisent
que
généralement
on ne faisait
pas le lien
entre cette
perte de
capacité
(vue
principalement
du point de
vue
évolutif)
et la
génétique
(qui se
contentait
d'étudier
l'hérédité
des
caractères
phénotypiques
dans une
perspective
évolutive
d'après
leurs propres
termes). Il
n'est pas
facile de se
rendre compte
de la
nouveauté
de ce
raisonnement :
au lieu
d'étudier
la
transmission
héréditaire
d'une
caractéristique
biochimique
connue (que
l'on associe
à une
portion de
chromosome
nommée
gène);
on
étudie
la biochimie
d'organismes
modifiés
artificiellement;
on est
vraiment au
nœud de
la relation
mécanisme
héréditaire
-
mécanisme
physiologique.
C'est pour
cela qu'il est
indispensable
dans le
protocole
proposé
d'étudier
la biochimie
de l'organisme
supposé
muté.
Il y a lien
supposé
entre une
capacité
biochimique et
un gène
sous un
allèle
et la perte de
cette
capacité
qui
correspondrait
à un
nouvel
allèle.
Mais on notera
que le mot
allèle
n'est pas
alors
utilisé
par ces
auteurs (il
sera
employé
par Beadle
dans sa publication
de 1945).
Expériences
et
résultats
:
Des
périthèces
de Neurospora
crassa et
de Neurospora
sitophila
sont
passés
aux rayons X
avant la
méïose.
Environ 2000
ascospores
recueillies.
Je
précise
que chez Neurospora
les asques
sont
habituellement
ordonnées
et donc que
l'on peut
numéroter
les ascospores
alignées
(voir cours de
Terminale).
Chaque spore
est mise en
culture sur un
milieu
contenant
toutes les
substances
habituellement
nécessaires
: agar, sels
inorganiques,
extrait de
malt, extrait
de levure et
glucose).
Certaines ne
germent pas.
Certaines
poussent
très
mal. D'autres
enfin donnent
des
mycéliums.
Des fragments
de ces
mycéliums
ou des spores
asexuées
issues de ces
mycéliums
peuvent
ensuite
être
repiqués
dans des
milieux de
culture
présentant
telle ou telle
caractéristique
(le milieu
minimum, MM,
contient
facultativement
de l'agar, des
sels
inorganiques,
de la biotine,
un
disaccharide
(dioside) et
un acide gras
ou une autre
source
complexe de
carbone). On
mesure la
croissance des
mycéliums
par leur
avancée,
depuis le
point de
dépôt,
dans un tube
de verre
horizontal
coudé
aux deux
extrémités.
On
peut souligner
que lors de
leurs
premières
expériences
les auteurs
n'étaient
absolument pas
certains
d'obtenir des
résultats
conformes
à leur
attente. C'est
presque avec
surprise
qu'ils ont
obtenu
à la
299ème
spore un
résultat
positif suivi
rapidement par
de très
nombreux
autres
(rapporté
dans la
conférence
de
réception
du prix Nobel
de Beadle ou
par Horowitz).
De
nombreux sites
proposent des
images du cycle
de Neurospora:
voici par
exemple celui
du Fungal
Genetics Stock
Center (Kansas
City) - Centre
de
conservation
de la
génétique
des
Champignons
que je vous
encourage
à
visiter
(http://www.fgsc.net)
à
l'adresse: http://www.fgsc.net/2000compendium/introduction.html.
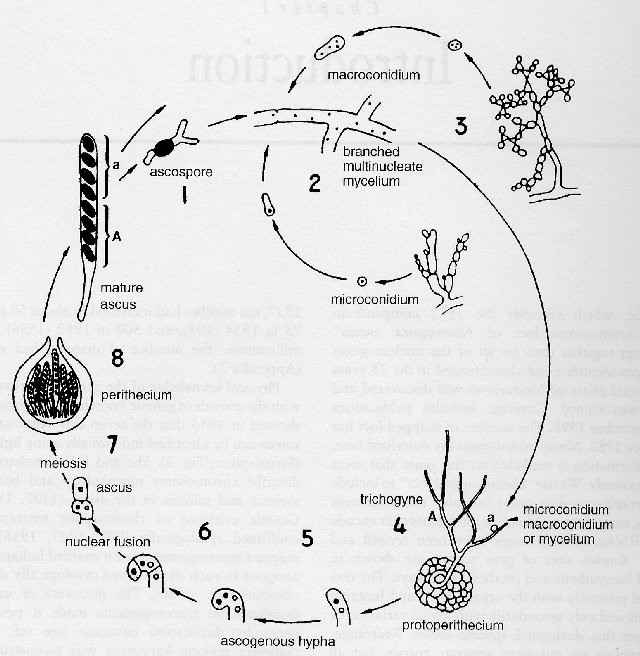
Cycle
de Neurospora
(légendes
en anglais
très
proches du
français)
(lien
permanent
établi
avec l'accord
gracieux de la
FGSC; qu'elle
en soit ici
remerciée)
Trois
souches
mutantes
(métaboliquement
parlant) sont
obtenues et sélectionnées
pour leur capacité
stable [et que
l'on suppose
transmissible
de
façon
stable sur
plusieurs
générations]
à se
développer
sur milieu
minimum (MM)
uniquement en
présence
d'une
substance
particulière.
Autrement dit
des souches
auxotrophes
sont
sélectionnées.
On fait
l'hypothèse
que l'origine
de cette
auxotrophie
est
génétique.
Seules trois
souches sont
rapportées
dans la
publication
alors que
Beadle affirme
en 1958 en
avoir obtenu
rapidement
plus d'une
dizaine...
Une souche de
N.
sitophila
(la n°
299) exigeant
la vitamine B6
additionnelle
(pyridoxine).
Une autre
souche de N.
sitophila
(n°1085)
ne se
développe
pas sans
vitamine B1
additionnelle
(thyamine,
isolée
par Tatum en
1936) mais des
tests
complémentaires
prouvèrent
que seule la
partie
thiazole de la
molécule
était
nécessaire.
Et une souche
de N.
crassa
exigeant
l'acide
para-amino-benzoïque.
Les
auteurs
s'efforcent
pour chaque
souche, et
selon leur
modèle,
de mettre en
relation la
vitesse de
croissance
avec la
quantité
de substance
ajoutée
au milieu de
culture. Pour
le premier
mutant la
relation est
assez nette
mais nettement
moins pour les
deux autres.
Pour ce qui
est de
l'interprétation
génétique,
le
raisonnement
est
très
simple. Ils
considèrent
comme
hypothèse
que la
déficience
est due
à un
gène si
par
fécondation
avec une spore
de la souche
sauvage (non
mutée)
puis
méïose
on
récupère
la
moitié
de spores de
type mutant et
la
moitié
de spores de
type sauvage.
Attention,
il faut bien
comprendre ici
que la notion
de gène
est celle
d'une portion
de chromosome
(ce dernier
étant
considéré
comme un
groupe de
liaison, voir
cours
de terminale
spécialité),
c'est la
notion de
gène
héréditaire
au sens
mendélien
ou morganien.
L'obtention de
myceliums
présentant
la même
déficience
que la souche
parentale
mutée
montre
l'héréditabilité
du
caractère.
On
notera combien
cette vision
reste sommaire
(stabilité
biochimique
à
suivre sur
plusieurs
générations)
et non
comprise
(quels sont
les
mécanismes
biochimiques
et
physiologiques
impliqués
dans cette
déficience
?).
Voici le
tableau du
seul
résultat
publié.
L'apparente
répartition
du
phénotype
des ascospores
en 50%
"muté",
50% "non
muté"
est l'argument
présenté
pour affirmer
que l'on a un
gène
(on dirait
maintenant
sous deux
allèles:
un
allèle
sauvage
correspondant
à la
capacité
à
synthétiser
sa propre
vitamine B6
considérée
comme
nécessaire;
et un
allèle
muté
correspondant
à la
perte de cette
capacité
et à
l'apparition
d'une nouvelle
capacité:
celle
d'utiliser une
vitamine B6
exogène).
TABLE 2
RESULTS OF
CLASSIFYING
SINGLE
ASCOSPORE
CULTURES FROM
THE CROSS OF
PYRIDOXINLESS
AND NORMAL N.
sitophila
Résultats
de la culture
de chacune des
ascospores
issues du
croisement
d'un
mycélium
de la souche
mutée
n°299
(exigeant de
la vitamine B6
dans le
milieu) avec
un
mycélium
non
muté de
Neurospora
sitophila
|
|
N°
de l'asque
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
|
1-16,
21
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
17
|
-
|
pdx
|
pdx
|
pdx
|
N
|
N
|
N
|
-
|
|
18
|
-
|
-
|
N
|
N
|
-
|
-
|
pdx
|
pdx
|
|
19
|
-
|
pdx
|
-
|
-
|
-
|
-
|
-
|
N
|
|
20
|
-
|
-
|
N
|
-
|
-
|
-
|
-
|
pdx
|
|
22
|
-
|
-
|
N
|
-
|
-
|
-
|
-
|
-
|
|
23
|
-
|
*
|
*
|
*
|
N
|
N
|
pdx
|
pdx
|
|
24
|
N
|
N
|
N
|
N
|
pdx
|
pdx
|
pdx
|
pdx
|
|
N
:
ascospore
germant et
ayant une
croissance
normale sur le
milieu sans B6
pdx :
ascospore
germant et
mycélium
ayant une
croissance
faible sur le
milieu sans B6
- : ascospore
non
germée
* :
incertitude
sur la
position de
l'ascospore (2
ont
germé
et
étaient
mutants ??)
La
ligne 1-16,21
a
été
ajoutée
à
partir des
données
du texte. Le
tableau ne
donnait que
les spores
ayant
germé,
avec cette
ligne elles y
sont toutes
!!!
|
|
|
Quelle
est la cause
de la non
germination de
certaines
spores ?
Comment ne pas
s'étonner
que la plupart
des ascospores
des 24 asques
étudiées
ne germent pas
(29
germinations
sur 24x8=192
spores soit un
taux de
germination de
15%)
La croissance
ralentie est
reliée
à la
mutation
parentale
n° 299
mais comment
le prouver ?
Le même
phénotype
n'est
peut-être
pas du tout
relié
à la
même
cause
biochimique ou
physiologique...
La croissance
normale est
reliée
à
l'autre souche
parentale,
mais là
aussi comment
le prouver ?
Il peut s'agir
d'une
transformation
ou d'un retour
à une
physiologie
différente...
On
ne peut pas
vraiment dire
que le concept
"un
gène-une
enzyme" soit
proposé
dans cette
publication,
même si
ce propos est
souvent tenu.
Les mots
exprimant
peut-être
cette
idée
sont les
premières
phrases de
l'article :
«
From the
standpoint of
physiological
genetics the
development
and
functioning of
an organism
consist
essentially of
an integrated
system of
chemical
reactions
controlled in
some manner by
genes. It is
entirely
tenable to
suppose that
these genes
which are
themselves a
part of the
system,
control or
regulate
specific
reactions in
the system
either by
acting
directly as
enzymes or by
determining
the
specificities
of enzymes....
Furthermore,
investigations
of this type
tend to
support the
assumption
that gene and
enzyme
specificities
are of the
same order
». Le
point de vue
actuel de la
physiologie
génétique
est que le
développement
et le
fonctionnement
d'un organisme
résultent
essentiellement
de celui d'un
système
intégré
contrôlé
d'une certaine
manière
par les
gènes.
Il est
raisonnable de
supposer que
ces
gènes,
qui sont
eux-mêmes
une partie du
système,
contrôlent
ou
régulent
des
réactions
spécifiques
de ce
système
soit en
agissant
directement
comme enzymes,
soit en
déterminant
la
spécificité
d'enzymes...
De plus, des
recherches de
ce type
tendent
à
prouver que
les
spécificités
du gène
et de l'enzyme
sont de
même
type
(traduction
"personnelle").
Horowitz qui
travaille avec
Beadle et
Tatum au
Caltech entre
1941 et 1946
affirme que ce
concept a mis
de nombreuses
années
avant
d'être
plus ou moins
conforté
par les
résultats
expérimentaux.
Peu
de temps
après
la publication
de cet article
Beadle
reçut
une lettre des
Laboratoires
Merck lui
demandant un
échantillon
de la souche
299 afin de
développer
un test pour
la pyridoxine.
Il envoya la
souche.
Quelques mois
plus tard les
chercheurs du
laboratoire
Merck (Stokes,
Foster, and
Woodward)
l'informèrent
que la souche
299 pouvait se
développer
sur MM si le
pH du milieu,
normalement de
5,
était
ajusté
à 6 (in
Horowitz,
1995). Le
laboratoire de
Beadle
découvrit
alors le
rôle
essentiel de
la
température
dans
l'expression
de leurs
"mutations"
(en cultivant
les souches
sur MM
à
35°C ou
à
25°C, ils
n'obtenaient
pas les
mêmes
résultats
:
généralement
le
phénotype
mutant
n'apparaît
pas à
25°C).
Plus
que le rôle
de
l'environnement
sur les
gènes,
comme ils ne
manquèrent
pas de le
proposer comme
interprétation
la plus
simple, c'est
une remise en
cause de la
méthode
sélective
qui
s'imposait...
mais ne
fût pas
faite. Horowitz
(1995)
rapporte
qu'ils
considérèrent
que la faible
fraction des
"mutants"
"sensibles
à la
température"
suffisait
à
conforter la
généralité
de
l'interprétation
génétique,
les cas de
thermosensibilité
étant
expliqués
par des
propriétés
moléculaires...
hypothétiques.
Des critiques
furent faites
du temps de
Beadle et
rapportées
ensuite (notamment
celle de
Delbrück
rapportée
par Horowitz
(1995) sur le
fait que les
mutants
sélectionnés
doivent se
développer
sur milieu
"complet" et
les spores ne
germant pas
sont
peut-être
des mutants
que l'on perd
avec cette
méthode
de la
sélection).
Mais les
réponses
apportées
(dans les
années
1951)
restèrent
dans le cercle
interprétatif
de la
biochimie
génétique.
C'est pourtant
la
méthode
qui pose un
problème
et non
l'interprétation
des
résultats.
Le
lien
déterministe
entre le
gène
supposé
muté et
la fonction
(synthèse
de vitamine
B6) est une
hypothèse
qui
préexiste
dans le
modèle
proposé.
Comment
peut-il
être
démontré
par
l'expérimentation
? Il y a une
profonde
erreur de
raisonnement
ici (voir
Nissim
Amzallag
ci-dessous).
|
|
Une
idée me
paraît
aussi
insuffisamment
soulignée
(elle
vient de
l'article de
Horowitz:One-gene-one-enzyme:
Remembering
biochemical
genetics,
N. H.
HOROWITZ, Protein
Sci. 1995
4: 1017-1019):
lorsqu'il
collabore avec
Tatum, qui est
un biochimiste
et un
microbiologiste,
Beadle ne peut
pas prendre
comme
modèle
une
bactérie,
dont pourtant
la biochimie
est en plein
développement
et pour
lesquels le
rôle des
mutations est
clairement
associé
à des
variations
biochimiques
car la
GÉNÉTIQUE
DES
BACTÉRIES
N'EXISTE PAS
ALORS*. Il
faudra
attendre de
nombreuses
années
avant que l'on
parle de
gènes
chez les
bactéries,
avec la
confusion que
cela a
entraîné
et
entraîne
encore: le
gène
étant
alors
défini
en
génétique
moléculaire
comme une
portion d'ADN
associée
à la
synthèse
d'un produit.
C'est pour
cela que
malgré
la
compétence
de Tatum pour
les
bactéries,
ils prennent Neurospora,
grâce
aux
études
réalisées
par B.O. Dodge
au Jardin
Botanique de
New York
dès
1928.
*Cette
affirmation
est inexacte
au sens
où
certains
chercheurs
étaient
persuadés
de l'existence
de
gènes
chez les
bactéries.
Il serait donc
plus exact de
dire que la
génétique
bactérienne
n'existait pas
alors au sens
de science
constituée.
Voici un
extrait de Gene
Recombination
in the
Bacterium
Escherichia
coli,
Tatum, Edward
L., and Joshua
Lederberg.,
1947, Journal
of
Bacteriology
53, 6, 673-684
(http://profiles.nlm.nih.gov/BB/A/B/E/P/_/bbabep.pdf)
où
Tatum
lui-même
explique ce
que l'on
entend par gène
chez les
bactéries
:
«
On the basis
of mutation
studies many
investigators
have concluded
that the
hereditary
properties of
bacteria are
based on the
existence of
genes (Luria
and
Delbrück,
1943; Roepke
et al., 1944;
Lwoff, 1941;
Demerec and
Fano, 1945;
Gray and
Tatum, 1944),
although it is
not clear
whether these
genes should
be homologized
with the
Mendelian
factors of
higher
organisms, or
with the
extranuclear
factors which
have been
demonstrated
in some
microorganisms
and higher
plants »
(Sur
la base
d'études
des mutations,
de nombreux
auteurs ont
affirmé
que les
caractéristiques
héréditaire
des
bactéries
sont
basées
sur
l'existence de
gènes
(Luria and
Delbrück,
1943; Roepke
et al., 1944;
Lwoff, 1941;
Demerec and
Fano, 1945;
Gray and
Tatum, 1944),
même
s'il reste
cependant
à
savoir si ces
gènes
sont les
homologues des
facteurs
Mendéliens
des organismes
supérieurs,
ou si ce sont
plutôt
des facteurs
extranucléaires
qui ont
été
découverts
chez certains
micro-organismes
et chez les
plantes
supérieures)
.
|
|
|
|
La
seconde
publication
signée
de Beadle seul
et
datée
de 1945 est
déjà
une revue du concept
un gène
- une enzyme
qui s'est
répandu
|
|
Beadle,
G. W.,
1945b Genetics
and metabolism
in Neurospora.
Physiol. Rev.
25:643-663
|
|
Ce
n'est que dans
le papier de
1945 (je
n'ai pas pu
accéder
à
toutes les
publications
mais seules
celles en
libre
accès
dans PubMed)
qu'apparaît
l'exemple de
la
chaîne
métabolique
du tryptophane
et
l'expression "one-gene-one-reaction"
qui sera
repris plus
tard comme le
"concept" "one-gene-one-enzyme".
On peut
trouver une
relation de
ces
idées
dans son
discours de
réception
du prix Nobel
en 1958 (http://nobelprize.org/nobel_prizes/medicine/laureates/1958/beadle-lecture.pdf).
L'analyse
de la liaison
génotype-phénotype
est cependant
plus riche que
dans la
publication
préliminaire
de 1941. Sans
remettre en
question la
méthode
qui leur donne
des
résultats
- plus de 100
mutants
isolés
(HOROWITZ,
N. H., D.
BONNER, H. K.
MITCHELL, E.
L. TATUM AND
G. W. BEADLE;
Am.
Naturalist.
79, 304, 1945,
publication
non
trouvée
sur le web)
-, Beadle
souligne que
l'on est
encore
à une
première
étape
de
l'étude
de
l'auxotrophie
par l'analyse
génétique
de mutants.
Notamment des
extraits de
levure
présents
dans le milieu
minimum ont
été
prouvés
contenir des
substances
inhibitrices
au
développement
de certains
mutants.
Ensuite le
rôle
sélectif
du pH a
été
aussi mis en
évidence.
L'ensemble de
ces
résultats
est cependant
exploité
biochimiquement
dans le
même
sens que
précédemment
: toutes les
exigences
biochimiques
d'un organisme
sont
susceptibles
d'être
contrôlées
par des
gènes
dont on peut
faire
l'exploration
au moyen de
mutations dont
on
sélectionne
les
représentants
sur des
milieux
appropriés.
Toute
idée
d'une
variabilité
métabolique
individuelle,
d'une
adaptation ou
de
transformation
au cours du
développement,
de la
redondance de
mécanismes
biochimiques...
est non
envisagée.
On s'enfonce
dans ce que
l'on pourrait
appeler
maintenant le
réductionnisme
biochimique.
|
|
|
En
laissant aux
historiens des
sciences le
soin de
démêler
les parts
respectives de
chacun (et
notamment
celle de la
fondation
Rockfeller, y
compris en
France, voir
par exemple
L'histoire de
la
génétique
à Gif
sur Yvette
racontée
par ses
acteurs: http://picardp1.ivry.cnrs.fr/histoire_de_la_genetique.html
)
dans
l'établissement
de la
génétique
moléculaire
comme
discipline
dominante, on
peut se
focaliser sur
la critique de
la
démarche
scientifique
de Beadle et
Tatum (et de
bien d'autres)
comme le fait
Amzallag dans
son ouvrage :
La raison
malmenée
(Nissim
Amzallag, CNRS
Éditions,
2002, p 34).
Il y affirme
que pour ces
travaux, et
pour une
grande part
des travaux
ultérieurs,
la
sélection
remplace la
compréhension
(ce
que me semble
avoir
été
clairement mis
en
lumière
dans les
analyses des
deux articles
ci-dessus):
|
La
sélection
a
posteriori d'individus
modifiés
n'est pas
seulement la
méthode
exclusive en
biotechnologie,
mais
également
la voie
d'investigation
privilégiée
du vivant.
C'est par
l'analyse de
mutants,
individus
reconnus comme
déficients
pour une
fonction, que
les
biologistes se
proposent de
comprendre le
fonctionnement
d'un organisme
normal. Il est
possible, au
moyen
d'irradiations
ou d'autres
techniques,
d'altérer
la structure
ou
l'expression
de petites
régions
du
génome.
L'analyse des
modifications
observées
en
parallèle
sur
l'organisme et
sur les
gènes
affectés
permet
d'établir
un lien de
causalité
entre
gènes
et fonctions,
génotype
et
phénotype.
Le cas
idéal
est bien
entendu celui
où il
est possible
d'établir
un lien direct
entre une
déficience
fonctionnelle
et
l'altération
d'un seul
gène,
comme c'est le
cas dans de
nombreuses
maladies dites
génétiques.
C'est la
première
mise en
évidence
d'une telle
relation qui
valut, en
1958, le prix
Nobel à
Georges Beadle
et Edward
Tatum. Depuis
lors, cette
approche est
devenue la
méthode
privilégiée
d'investigation,
celle qui
ouvrit la voie
à la
biologie
moléculaire
et à la
correspondance
étroite
entre science
et technologie
dans le
domaine du
vivant.
Cependant, la
méthode
en question
recèle
un très
sérieux
travers. La
technique de
sélection
mène
à
l'identification
de mutants
modifiés
pour une
fonction, sur
lesquels
l'analyse
génétique
se focalise
ensuite. Le
phénotype
est donc le
révélateur
des
modifications
génétiques.
Or, une telle
méthodologie
implique un
lien déterministe
entre le
génotype
et le
phénotype,
alors que
ce lien
prétend
être
lui-même
démontré
par
l'expérience.
En effet, elle
suppose que le
gène
modifié
au point de
perturber la
fonction de
son produit
d'expression
implique
nécessairement
une carence au
niveau
phénotypique.
Cependant, il
est impossible
de
déterminer
si, au sein de
l'immense
majorité
des individus
non
sélectionnables
parce que
n'exhibant
aucune
anomalie
visible, ne se
trouvaient pas
également
des individus
dont le
gène en
question
était
lui aussi
modifié.
(retour)
|
|
|