Une (nouvelle) théorie de
l'évolution
(en
travaux)
d'après R. Chandebois
Pour en finir avec le
Darwinisme, Espaces 34, 1993 (cité PFD dans le texte)
Comment les cellules construisent l'animal, Phénix
Éditions, 1999 (cité CCCA dans le texte)
L'essentiel des idées
présentées ici viennent de Mme Rosine Chandebois (voir
les références ci-dessus et d'autres dans la
bibliographie).
Bien évidemment, j'assume les déformations et
incompréhensions que j'ai pu y ajouter, ainsi que ce qui me
paraît être mon propre fond, dont je ne connais pas
toujours l'origine.
1. Essai de formulation de la théorie et de ses lois
L'évolution est à la fois
stabilité
et
variation
de la forme, des structures et des fonctions.
L'évolution c'est le résultat du travail de la vie.
C'est un travail commencé il y a peut-être des milliards
d'années et qui se répète dans chaque cellule.
La plupart du temps le travail est identique à lui-même:
chaque cellule donne naissance à une nouvelle cellule, qui
reproduit les mêmes propriétés que la cellule
mère.
Cette répétition repose sur la théorie
cellulaire (voir par exemple le cours
de seconde, §1.3), qui englobe
une théorie de la stabilité de la forme et des
fonctions que l'on peut appeler automatisme (toute cellule est
issue d'une autre cellule). Le problème de
l'origine de la vie est un problème à part
traité dans l'ancien cours
de terminale S. Mais si l'on veut
que les organismes puissent évoluer il faut un
mécanisme capable d'engendrer la variation et de la
mémoriser. Mais attention, ce dynamisme peut avoir une
origine interne: ce n'est pas la surimposition d'un
élément nouveau, mémorisé, mais ce peut
être une sorte de répertoire naturel de variation, un
futur toujours ouvert, bref une caractéristique profonde du
vivant. Les termes de comportement social (utilisé par
l'auteur) ou de travail sont toujours
préférables à celui de
mémoire.
Le terme de travail englobe et dépasse ceux
d'automatisme et de variation car, tout en évoquant la
répétition d'un même phénomène, il
laisse entendre, philosophiquement, que chaque nouvelle vie, chaque
instant du travail du vivant, est l'expression de quelque chose de
vraiment nouveau car il ne s'est jamais déroulé dans
les mêmes conditions par le passé (voir
par exemple la page sur le
temps du cours de terminale).
Du point de vue scientifique, il existe au moins deux
modèles qui rendent compte de ce travail ou de cette
stabilité et de cette variation.
- Le modèle le plus classique, depuis le milieu du
XXème siècle et jusqu'à maintenant,
était celui d'un programme génétique,
stable, qui transmettait fidèlement à la descendance
les caractères de l'espèce, doublé d'une
variation et mémorisation génotypique, dont
le mécanisme reposait sur l'accumulation de mutations dans
l'ADN. Le sens de l'évolution était donné par
la sélection naturelle.
- Depuis une dizaine d'années il existe un autre
modèle présenté par une embryologiste, R.
Chandebois, qui remplace le programme génétique par
un fonds cytoplasmique de l'espèce (stable car
retrouvé intact pour chaque individu dans l'ovocyte) auquel
s'ajoute l'automatisme de l'évolution (la
lignée comme système autonome) qui est en quelque
sorte une extrapolation de l'automatisme embryonnaire
(l'individu comme système autonome). La variation
reposerait d'abord sur la modification du fonds cytoplasmique
de l'espèce, dans la lignée ovulaire,
responsable de l'évolution directionnelle, mais
aussi sur des mutations qui engendreraient la variation
génotypique, responsable de l'évolution
adventive.
A la notion de programme de développement (pour
lequel aucune expérience n'a jamais pu mettre en
évidence le début d'une preuve) R. Chandebois oppose,
pour les pluricellulaires, l'automatisme embryonnaire
caractérisé par une progression autonome et des
réajustements, phénomènes
réellement observables et dont la validité peut
être testée expérimentalement en embryologie.
L'autonomie du développement embryonnaire, principalement
étudié chez les animaux, repose sur la double
capacité de chaque cellule à répondre
à chaque instant aux signaux qu'elle reçoit des
cellules voisines et à mémoriser son histoire: ce que
R. Chandebois appelle le comportement social
élémentaire. Si ce comportement autonome reste un
mystère, c'est celui de la vie. En effet, l'autonomie d'un
organisme unicellulaire se résume aux trois aspects du travail
du vivant: travail de relation, de nutrition et de reproduction
(voir la
diversité du vivant). Cette
interprétation s'intègre donc aisément dans une
vision "sociale" du travail du vivant et est d'une grande richesse
pour des expériences à mener.
L'automatisme de l'évolution (évolution
directionnelle et évolution adventive) est par contre du
domaine théorique historique mais fournit un cadre pour
l'interprétation des données
paléontologiques.
Pour les pluricellulaires, le système cellule est
coiffé par un système hiérarchiquement
supérieur, le système individu au niveau duquel
sont coordonnées les progressions autonomes des
différentes populations cellulaires composant l'organisme,
notamment par le moyen de substances chimiques.
* Le système lignée ou la complexification des
formes vivantes dans l'espace (de petites formes à de grandes
formes, du milieu aquatique à tous les milieux terrestres... )
et dans le temps (des organismes dont la durée de
développement s'allonge et dont la durée de vie
s'allonge...) a depuis longtemps été
résumée par une loi qui est celle de la division du
travail (la formulation d'Haeckel, pourtant
identique, recouvrait des concepts très différents,
voir ancien cours de
terminale &4.a2).
Plus le travail du vivant est réparti
entre différentes structures, au sein d'un organisme, plus ce
dernier est évolué (ou récent
phylogenétiquement).
Cette division du travail résume une succession
d'innovations qui nous permettent de bâtir la
classification évolutive (phylogénétique) du
vivant. Parmi celles-ci on peut citer: la compartimentation de la
cellule (des procaryotes aux eucaryotes), la spécialisation et
différenciation cellulaire (des unicellulaires aux
pluricellulaires) ou encore la différenciation organique (des
thallophytes aux plantes...). Voir une
présentation de cette classification la
diversité du vivant.
* L'embryologie ajoute un autre élément qui semble
caractériser l'évolution: c'est l'allongement de la
période de développement. Ce que l'on pourrait appeler
le temps de travail ou, avec un clin d'œil à Bergson,
la durée du travail.
Plus le travail du développement qui
donne naissance à un organisme dure, plus ce dernier est
évolué (c'est-à-dire récent,
phylogenétiquement).
Plus la durée du développement augmente,
plus les interactions entre cellules différenciées sont
nombreuses et complexes et conduisent à des organismes
évolués et complexes.
2. Quelques exemples (en cours
d'élaboration)
Étant donné les profonds changements d'ordre de
grandeur entre les étapes évolutives qui font passer
par exemple un ancêtre procaryote à l'état
eucaryote, un eucaryote unicellulaire à un eucaryote
métazoaire, un métazoaire invertébré
à un métazoaire vertébré, ou d'un
ancêtre des félidé au chat actuel,
peut-être serait-il prudent de présenter plusieurs
classifications phylogénétiques. Le premier
problème à mettre en avant, sans pouvoir le
résoudre, du moins dans le cadre d'un tel cours, est celui du
choix, pour toutes les étapes évoquées, entre
une origine monophylétique (une seule lignée ou clade)
et une origine polyphylétique (plusieurs lignées
conduisant à un groupe). Les arguments sont parfois solides
pour les deux hypothèses, mais ne seront bien sûr pas
détaillés ici.
On gardera ici la classification du monde vivant en 5
règnes, comme il a été vu en classe
de seconde (&1.4).
Pour chaque règne, je renvoie aux
généralités présentées dans une
page annexe sur la diversité du vivant,
dont voici cependant les grandes lignes:
- les Procaryotes = un compartiment unique où se
réalisent des travaux multiples; des populations
innombrables de travailleurs aquatiques, cachés et
silencieux
- les Protistes = une cellule compartimentée en
organites réalisant chacun un travail spécialiste ;
des populations parfois regroupées mais qui restent
inféodées à l'eau
- les pluricellulaires: des sociétés cellulaires
orientés vers des travaux différents dans
l'écosystème
- les Mycètes : des sociétés
très diversifiées qui unissent, tissent des liens,
recyclent, transforment...
- les Plantes : des sociétés de cellules
qui se fixent en un lieu et colonisent l'espace; les plantes ont
conquis le milieu aérien
- les Animaux : des sociétés de cellules
qui se déplacent au sein de leur milieu de vie; de nombreux
animaux ont conquis le milieu aérien
- et si l'on ajoute un 6ème règne: les
Hommes (biologiques): des sociétés de
cellules se regroupant en sociétés d'individus et
cherchant à maîtriser la terre
Deux grandes étapes évolutives avec deux exemples de
théories évolutives :
- De l'état procaryote à l'état
eucaryote:
la principale théorie actuelle est celle de
l'endosymbiose (du grec
endo=intérieur, sym =avec; bios=vie):
les eucaryotes résulteraient d'une symbiose (vie commune
avec bénéfices réciproques) de procaryotes
à l'intérieur de la cellule eucaryote donnant
notamment les mitochondries et peut-être les chloroplastes;
un des arguments les plus puissants concerne les ARNr contenus
dans les mitochondries dont certaines sous-unités sont
incontestablement plus proches de celles de certains procaryotes
que ceux du cytoplasme; le polyphylétisme semble le plus
communément admis; ce qui justifieraient les innombrables
modalités reproductrices des Protistes.
Pour un panorama de la diversité de vie des
Protistes je renvoie à la page générale sur
la diversité du
vivant.
- L'origine des animaux:
la théorie actuelle la plus attrayante est celle
proposée par R. Chandebois à partir de Protistes
flagellés (dans Pour en finir avec le darwinisme, R.
Chandebois, Espaces 34, 1993, p 182-186).
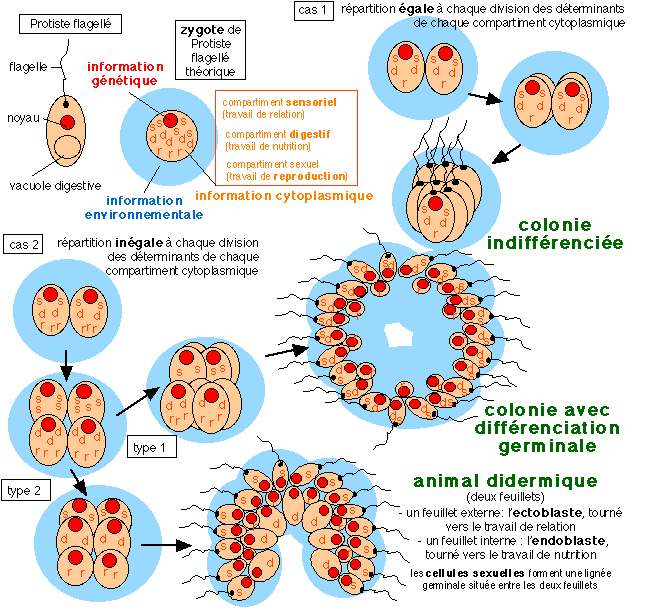
|
Une représentation de l'origine possible de
structures pluricellulaires à partir d'UN
Protiste Flagellé (phagotrophe
c'est-à-dire qui se nourrit de particules solides
en suspension par opposition à osmostrophe, qui se
nourrit de substances dissoutes, comme le font les
plantes notamment) qui donne en UNE unique
génération un descendant
pluricellulaire, capable d'autoreproduction sous la forme
de pluricellulaire. Le polyphylétisme étant
bien sûr possible: plusieurs flagellés
pouvant donner des descendants évolués
pluricellulaires de plusieurs types. C'est en effet le
fonds cytoplasmique qui
porte la mémoire de l'espèce
représentée ici sous forme de
"déterminants" (mais attention il
ne s'agit pas des éléments d'un programme
mais des composants (ARN associés à des
protéines généralement) de chaque
compartiments qui déterminent la
réalisation de tel ou tel travail cellulaire, le
moteur de la différenciation est dans la suite
d'interactions entre cellules engagées dans une
vie autonome, qui passe par des divisions cellulaires
répétées: c'est une progression
autonome) correspondant aux trois types de travail
du vivant (relation
(s, sensoriel),
nutrition (d,
digestif) et reproduction
(r)).
La lignée ovocytaire récupère
ensuite sa totipotence, c'est-à-dire la
totalité de son fonds cytoplasmique. A la
génération suivante les zygotes de
flagellés commenceront une progression autonome
qui réitérera celle qui leur a donné
naissance du fait du fonds cytoplasmique modifié
qu'ils auront reçu (mémoire) et des
interactions qu'ils auront avec leur environnement (leur
comportement social).
L'événement évolutif du passage d'un
unicellulaire à un pluricellulaire est donc une
induction, qui a eu lieu une fois, puis a
été mémorisée et se
répète depuis dans chaque
lignée, pour chaque individu, si son
développement n'est pas perturbé. Cette
évolution est automatique, directionnelle, elle
présente une finalité interne. Les
philosophes disent que l'organisation d'un
pluricellulaire est en puissance dans les
caractères sociaux de l'unicellulaire.
Le sens de cette induction peut être exprimé
comme une division du travail de l'organisme qui
le répartit en plusieurs types cellulaires
spécialisés. C'est une
différenciation cellulaire. Elle peut
s'expliquer par des différences de rendements
entre gènes en supposant, dans un premiers
temps, que toutes les cellules ont la même
information génétique. Ce qui
différencie le comportement puis la structure des
cellules de l'endoblaste par rapport à celles de
l'ectoblaste, c'est leur activité
métabolique. Celle-ci va déterminer
à son tour leur mémoire qui les rendra
capables d'autres déterminations
éventuelles dans la suite du
développement... le développement est ainsi
vu comme une suite d'induction - comportement social -
mémorisation et acquisition d'une nouvelle
compétence ...).
(très modifié
d'après R. Chandebois, Pour en finir avec le
darwinisme, Espaces 34, 1993, p 184, Fig.10; je ne
suis pas à l'abri d'erreurs de
compréhension...)
|
En résumé voici une représentation
simplifiée de l'arbre phylogénétique du vivant
avec les étapes majeures (inductions) de
l'évolution:
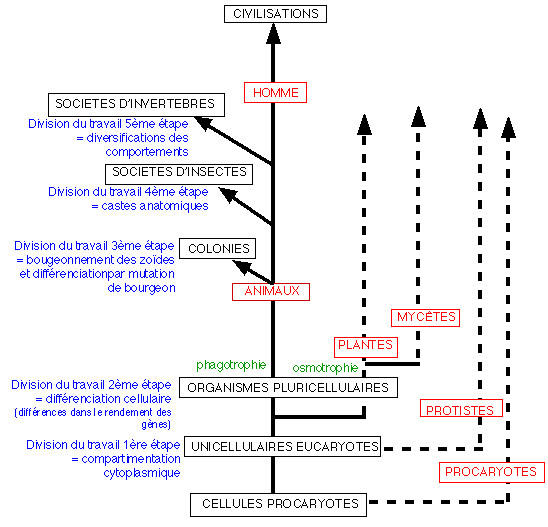
Les étapes majeures de
l'évolution,
une représentation qui n'élude pas la question
biologique de l'homme.
|
En bleu les 5 grandes
étapes (inductions)
isolées par R. Chandebois, pouvant être
regroupées en terme de division
du travail (division reprise pour
chaque règne dans la page annexe sur la
diversité du
vivant).
L'évolution directionnelle
représentée par des flèches
dirigées vers le haut du schéma (temps
actuels) ne signifie pas que l'évolution est
déterminée par une finalité
externe (contrairement à ce que l'auteur peut
faire penser en utilisant le terme progrès
dans son schéma et que nous n'avons bien
sûr pas repris; voir Dominique Letellier, La
question du hasard dans l'évolution, L'harmattan,
2002, p 136-139) mais que c'est cette finalité
interne (voir cours
de TS), qui est un
automatisme que l'on pourrait qualifier - selon cet
auteur - de « faculté des organismes
à s'auto-organiser dès lors qu'ils se sont
donnés à eux-mêmes les moyens de le
faire », qui a conduit à cette histoire
là , que nous essayons de déchiffrer . Cette
histoire repose sur l'étude des archives
paléontologiques (très pauvres pour les
étapes présentées ici) et sa
connaissance est donc perfectible.
(MODIFIÉ d'après R.
Chandebois, Pour en finir avec le darwinisme, Espaces
34, 1993, p 208, Fig.12); les
5 règnes
ainsi que l'homme ont
été individualisés.
|
3. Quelques questions-réponses autour de cette
théorie
(les réponses n'engagent que ma
compréhension personnelle ).
Quelles différences y-a-t-il entre évolution
orientée et évolution directionnelle ?
Dire que l'évolution est orientée, c'est
affirmer qu'elle à un sens (en mathématique on utilise
ce terme pour les vecteurs et l'analogie est très forte
puisque de nombreuses personnes se représentent le temps comme
une flèche, et donc avec une orientation). Dire que
l'évolution est orientée est donc une évidence
si l'on ne désigne par là que le temps de
l'évolution. Par contre, dire que l'évolution est
directionnelle (toujours cette analogie avec la
flèche du temps) suppose qu'elle est unique, qu'elle emprunte
un chemin unique, déterminé en quelque sorte. Ce
qui est un tout autre problème...non biologique. Pourrait-elle
"reculer" c'est-à-dire aller dans un sens différent ?
Clairement non, le passé est définitivement
passé. De la même manière, pourrait-elle changer
de direction ? Et dans ce cas, par rapport à quelle direction
? D'après la théorie de Mme Chandebois, on pourrait
dire que la direction de l'évolution est unique, car elle
n'est pas déterminée par le futur mais par le
passé. En ce sens, l'évolution n'est pas directionnelle
mais elle est bien orientée.
Quel est le moteur de l'évolution ?
C'est la vie, dans son caractère autonome et
fragile ou adaptable: dans sa stabilité qui
n'empêche pas la variation.
Certains verront dans l'évolution du vivant une constante
adaptation, un réajustement, pourrait-on-dire, mais à
l'échelle écologique et non plus de l'individu.
Du point de vue philosophique, on peut mettre dans la
première cellule - en supposant que tous les êtres
vivants dérivent d'une cellule unique - toute la
capacité d'être (à l'état de
possible) de l'ensemble des êtres vivants de tous les temps (ce
que fera un matérialiste). On peut au contraire penser que la
matière n'est que le lieu de cette capacité
d'être et que le principe vital (l'être en tant
qu'acte) est ailleurs et accompagne sans cesse chaque être
vivant dans sa vie (ce qui est plus ou moins un
vitalisme).
L'évolution est-elle terminée ? Que peut-elle
encore donner ?
Sans jouer les prophètes on peut, à mon avis, adopter
raisonnablement au moins deux attitudes, qui ne sont pas
incompatibles d'ailleurs:
- soit penser avec P.P. Grassé (&3.d.2)
et R. Chandebois que l'évolution actuelle se résume
à la spéciation et que donc l'évolution est en
train de se terminer: «...la marge de manœuvre de
l'évolution n'a cessé de s'amenuiser : à
l'Ordovicien, la genèse des embranchements s'arrête, au
Jurassique, celle des classes, au
Paléocène-Eocène, celle des ordres »
(L'évolution du vivant, P.P. Grassé, Albin
Michel, 1973, p 124); cet amortissement cadre avec l'alternance de
l'évolution directionnelle et adventive, une évolution
directionnelle intervenant maintenant ne peut faire sortir les
individus du cadre de l'espèce (voir PFD, p 115
[38]).
- soit penser qu'un nouveau venu: l'homme, est capable, par sa
technique, de diriger cette évolution comme nous le
suggère par exemple G. Tibon-Cornillot dans son ouvrage
Les corps transfigurés.
Cette dangereuse liberté exige que l'homme agisse avec un
respect total du vivant, à commencer par lui...
L'ontogenèse évolutive repose sur une exploitation des
potentialités de l'œuf (voir PFD, p 115 [39])
mais l'homme, par sa technique peut, à chaque étape
modifier le déroulement de la progression autonome et donc
multiplier les possibles.
L'évolution aurait-elle pu donner autre chose ? Si
l'on rembobinait le film du temps pourrait-on voir se dérouler
un autre film ?
Dans le cadre de la théorie présentée ici, et du
fait du finalisme interne (voir question suivante) de
l'évolution, la profonde similitude entre l'ontogenèse
d'un individu et celle du monde animal montre
l'irréversibilité et l'originalité du
phénomène vivant qui ne saurait être autrement
que celui qui a eu lieu, historiquement (PFD, p 115
[39]).
Mais en fait la question n'est pas scientifique ni
paléontologique mais philosophique et Bergson à mon
avis à un réponse bien plus profonde que l'habituelle
opposition entre un déterminisme aléatoire (hasardeux)
et un déterminisme orienté (finalisme). Pour lui, le
futur n'est même pas un possible car il comprend des actes
libres de l'homme. «... les philosophes qui croient que
l'avenir est donné dans le présent... qui ont cru au
libre arbitre, qui l'ont réduit à un simple "choix"
entre deux ou plusieurs partis, comme si ces partis étaient
des "possibles" dessinés d'avance... D'une action qui serait
entièrement neuve (au moins par le dedans) et qui ne
préexisterait en aucune manière, pas même sous
forme de pur possible, à sa réalisation, ils semblent
ne se faire aucune idée. Telle est pourtant l'action
libre.»... «Radicale est donc la différence entre
une évolution dont les phases continues
s'entrepénètrent par une espèce de croissance
intérieure, et un déroulement dont les parties
distinctes se juxtaposent.» (La Pensée et le
Mouvant, in Œuvres, Bergson, 2ème édition du
centenaire, PUF, 1963).
Cette compréhension de l'évolution n'est-elle
pas finaliste ?
Au sens de finalité externe, non. Cependant, comme
toute interprétation physiologique ou tournée vers la
vie elle présente bien sûr une finalité
interne. Pour une discussion sur ces mots voir le
cours de
TS et une
page du cours des PE ou directement
l'ouvrage de D. Letellier (La question du hasard dans
l'évolution, L'harmattan, 2002, p 27-29). Cette vision
de l'évolution est téléologique (du grec
teleo = la fin, le but) dans le sens où les êtres
vivants sont en quelque sorte les produits de l'automatisme de
l'évolution qui est donc dirigée ou directionnelle si
l'on veut (voir ci-dessus). L'évolution n'obéit pas
à un programme (que l'on a jamais trouvé) mais est le
résultat des innombrables interactions (que l'on
connaît) et mémorisations (que l'on observe et que l'on
peut expérimenter) des processus du vivant
réalisés par la vie au cours du temps. Ce qui est tout
le contraire d'un déterminisme externe.
Quelle différence y-a-t-il entre progrès et
évolution téléologique ?
Le terme de progrès a plusieurs sens. Le sens
biologique de progression ou développement est tout à
fait compatible avec l'évolution. Mais le sens d'augmentation,
qui fait référence à un bien, et en
définitive à une finalité externe, n'est pas
acceptable. Le progrès pourrait alors désigner un
déterminisme de l'évolution qui ne serait pas le seul
fait du vivant. Au contraire, le terme d'évolution
téléologique évacue cette dimension
externe pour ne désigner que la finalité interne, qui
est caractéristique du vivant: le travail du vivant (voir
question précédente).
Cette théorie de l'évolution est-elle
scientifique ?
Dans la mesure où c'est une théorie du vivant en
développement, elle est expérimentale. Mais
à partir du moment où elle cherche à expliquer
comment les formes vivantes ont évolué les unes vers
les autres par le passé, elle devient historique en
intégrant des arguments de la paléontologie (voir
méthode). Cette théorie ne se
rapporte par contre à aucune option philosophique de
façon absolue et peut très bien être
acceptée dans le cadre d'un matérialisme que d'un
vitalisme, ce qui n'était pas vraiment le cas du
néodarwinisme.
La formulation de la théorie de Haeckel selon
laquelle «l'ontogenèse récapitule la
phylogenèse» semble bien cadrer avec cette
théorie? Qu'en est-il réellement ?
Cette phrase de Haeckel cadre fort bien avec la théorie de Mme
Chandebois uniquement si l'on comprend chacun des mots de cette
formule dans le sens proposé ci-dessus: la
phylogenèse, c'est la somme des
innovations par lesquelles doit passer l'embryon
au cours de son ontogenèse pour arriver à
l'étape correspondant à la progression autonome de son
phylum. Mais il est évident qu'au sens d'Haeckel cette
formule recouvrait tout autre-chose: voir à ce sujet
l'ancien cours de terminale (&4a2).
La théorie de la transmission de la
mémoire de l'espèce
par le fonds cytoplasmique ovulaire n'est-il pas une théorie
"maternaliste" ou "féministe" ? Où est passée
l'égalité de la génétique chromosomique
?
C'est une théorie "maternaliste" si l'on considère que
le fonds cytoplasmique est PLUS que l'indispensable
déclenchement de la reprise de l'automatisme
ontogénétique, qui ne peut se faire que par la cellule
sexuelle mâle capacitée. Les nombreuses
parthénogenèse provoquées ou naturelles en sont
l'illustration.
Mais l'information génétique apportée par le
spermatozoïde n'en reste pas moins indispensable pour
l'équilibre génotypique. Ce qui est maternel ou
féminin dans la théorie téléologique
c'est la transmission de l'automatisme du développement. Comme
pour le corps humain où, l'organisme maternel est le seul
capable de mener à bien le développement embryonnaire.
En ce sens, seul le cytoplasme de l'ovocyte, cellule sexuelle
femelle, est capable de démarrer le développement de
l'embryon.
Comment répondre au problème de la
sexualisation ? Notamment lors du passage de l'état
unicellulaire à l'état pluricellulaire.
Chez un unicellulaire Flagellé, par exemple, la cellule
végétative se sexualise progressivement sous l'action
de certains facteurs (on considère notamment que des
conditions défavorables à la survie comme le manque de
nourriture sont inducteurs). La maturation sexuelle, longue, se
termine par la condensation des chromosomes puis par leur
appariement, auquel font suite deux divisions. L'ensemble est
qualifié de méiose. Comme j'ai essayé de le
proposer (voir page
sur la méiose), la
sexualisation peut être décomposée en une
maturation sexuelle, longue, suivie d'une "mitose sexuelle" rapide
sans séparation des chromosomes homologues appariés
puis d'une mitose habituelle, elle aussi rapide. Les 4 cellules
obtenues habituellement sont des gamètes haploïdes. Pour
les Protistes, les anomalies de méiose et de mitoses sont
très nombreuses, en regard du peu de cas où elles ont
été observées et décrites. Les
cytodièrèses sont souvent absentes et de nombreux
noyaux issus des caryodièrèses (imparfaites)
dégénèrent. On peut interpréter ces
grandes variations de modalité de la sexualisation des
Protistes comme étant le résultat d'un fort
polyphylétisme.
Dans le cas des Protistes le fonds cytoplasmique de l'espèce
ne quitte jamais l'unique cellule, même lors de sa
sexualisation. Il n'y a donc pas de problème de transmission
à la génération suivante.
Dans le cas des pluricellulaires, la persistance d'une lignée
germinale, a été la réponse habituelle à
la question de la sexualisation des individus. Cette lignée
germinale peut être ici invoquée pour désigner
une lignée de cellules qui conserve le fonds cytoplasmique
puisque celui-ci est altéré par la
différenciation des cellules somatiques. Cependant le soma
semble toujours nécessaire à la
récupération du fond cytoplasmique notamment par le
transfert, vers les cellules germinales de réserves et surtout
d'ARN (et probablement d'ADN par l'intermédiaire d'ARN
transformants, voir page
sur le développement). Les cellules germinales
immatures ne donnent jamais des embryons viables. La distinction
soma-germen n'est pas aussi tranchée qu'on veut parfois
l'affirmer.
Une des tendances évolutives sera l'allongement de la
maturation sexuelle (au sens proposé ci-dessus) qui
dépendra toujours plus de cellules somatiques
différenciées et nombreuses.
Comment concilier des termes
comme l'automatisme ou le comportement social
élémentaire avec les données modernes de la
génétique du développement ?
La théorie de Mme Chandebois, même s'il elle se place
à un niveau de compréhension d'ordre supérieur,
n'empêche pas l'intégration des données de la
génétique du développement, bien au contraire.
Les tenants d'un programme génétique de
développement font appel d'un part au concept de
programme génétique - qui, pour
être universellement employé, n'en est pas moins vague
(voir discussion dans le programme
de la classe de seconde)- et
d'autre part à la notion de libération
différentielle de l'information
génétique. Dans ce cadre, la
différentiation cellulaire se met en place grâce
à l'activation de certains gènes spécifiques et
la répression d'autres gènes. À la place de
cette alternative (activation/répression) R. Chandebois
propose (à la suite d'autres auteurs) la théorie
d'une différenciation cellulaire basée sur des
différences de rendements des gènes: certains
gènes, dont l'activité dépasse un certain
seuil de différenciation, sont fortement activés
et signent le métabolisme principal de la cellule, d'autres
gènes, plus ou moins faiblement activés, sont
responsables des potentialités histogénétiques
de la cellule; d'autres gènes enfin, fortement
réprimés, et habituellement de façon
irréversible, représentent en quelque sorte de
l'histoire de la cellule. Cette théorie a été
notamment développée par Clayton (Stability and
Switching in Cellular Differenciation, R.M. Clayton ed., Plenum
Press, New York, 1982 et autres sources citées in CCCA, p
84).
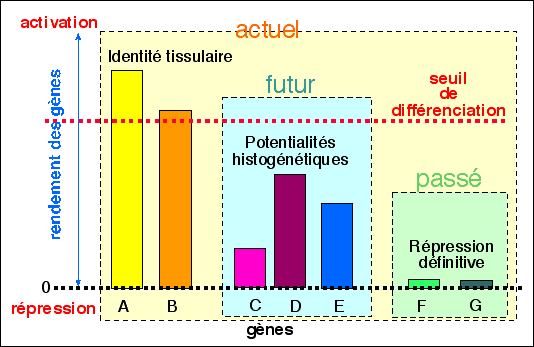
|
|
Une représentation d'une théorie de
la différenciation selon le rendement
des gènes
(d'après R. Chandebois,
CCCA, 1999, p 18),
modifié)
Dans une cellule, au sein d'un organisme en
développement, les gènes A à G ont
différents degré d'activation
répression: les gènes A et B sont fortement
activés et signent l'identité tissulaire,
responsables en grande partie du métabolisme
spécifique de la cellule; les gènes C, D et E
sont plus ou moins activés et réprimés
mais sont susceptibles d'avoir un rendement différent
si la cellule est placée dans des conditions
environnementales différentes, c'est en quelque sorte
la plasticité fonctionnelle de la cellule ou ce sont
les potentialités histogénétiques au
cours du développement (le "futur" de la cellule);
enfin les gènes F et G sont fortement
réprimés et, dans des conditions
physiologiques normales, de façon irréversible
(le "passé" de la cellule).
|
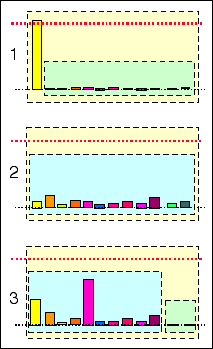
D'autres exemples théoriques sous forme
d'un test
Retrouver les correspondances entre les étapes
décrites ci-dessous (a, b et c) et leur illustration
ci-dessus (1, 2 et 3).
Trois illustrations correspondant à trois cellules
d'une même lignée (issues les unes des autres
par mitoses) ; par exemple :
a - une cellule de l'ectoblaste (stade blastula);
b - une cellule épidermique (stade bourgeon
caudal);
c - un kératinocyte (cellule
différenciée de l'épiderme).
(réponse: 1c; 2a;
3b)
|
Pourriez-vous donner un exemple plus précis de mise
en œuvre de ces théories ou cours de
l'embryogénèse ?
Un exemple mêlant ontogénie et phylogénie
a été donné par R. Chandebois (PFD, p 132 s) et
concerne le cartilage. Du fait de l'âge
déjà avancé (1993) des données
scientifiques présentées, voici une petite
mise-à-jour personnelle.
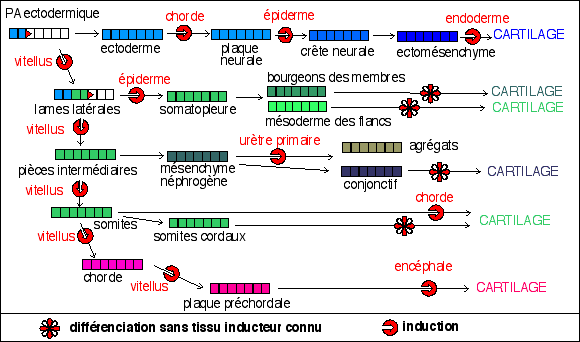
Origines diverses du cartilage à la suite du
développement in vivo et lors d'expériences in
vitro chez différents
Vertébrés (?) (d'après RC, PFD, p 133,
fig 7)
|
Le cartilage provient aussi bien de
l'ectoderme que du
mésoderme. Les
inducteurs sont
différents selon les moments et les lieux où
le cartilage est induit. Les inducteurs ne sont pas non plus
interchangeables. Dans certains cas le cartilage peut
émerger spontanément : dans les somites
cordaux isolés; dans le mésenchyme des
bourgeons des membres qui abouti à des îlots de
précartilage; et enfin dans le mésoderme des
flancs dans le cas où celui-ci est lésé
expérimentalement par l'introduction d'un corps
étranger ou isolé en culture. Enfin, le
cartilage peut être induit in vitro dans des tissus
qui normalement ne forment pas de cartilage comme le
mésenchyme néphrogène non soumis
à l'induction de l'urètre primaire.
|
|
La chondroïtine sulfate, un marqueur de la
formation du cartilage
Le tissu cartilagineux est composé de cellules
conjonctives sécrétant une matrice
fondamentale particulière. Cette matrice comprend
des substances protéiques comme le collagène
et l'élastine mais aussi des composés
glycoprotéiques nommés
protéoglycannes (anciens
mucopolysaccharides; en anglais proteoglycans
[PGs]).
Ce sont de gigantesques molécules ramifiées
formées d'un long filament de hyaluronate (un
hétéropolysaccharide) auquel se rattachent de
très nombreuses chaînes formées chacune
d'un filament protéique sur lequel sont
attachés d'innombrables chaînes glucidiques de
type kératane sulfate et chondroïtine
sulfate (des hétéropolysaccharides).
Les chondroïtine sulfates ("chondroitin sulfate"
(CS) ou "CS proteoglycans" (CSPGs) en anglais) sont
aussi associées à certaines molécules
de collagène (soutien de la structure fibrillaire de
la molécule). La synthèse extrêmement
abondante de chondroïtine sulfate est ainsi un bon
marqueur du métabolisme des cellules cartilagineuses
et osseuses.
Sans pouvoir entrer dans le domaine florissant de la
recherche des gènes impliqués dans la
synthèse des chondroïtine sulfates et avoir une
vue exhaustive des fonctions de ces dernières, on
peut juste citer que l'on a récemment cloné le
gène de la chondroïtine synthase. Les
chondroïtine sulfates sont reconnues dans les
articulations comme des marqueurs de l'effort ou des
pathologies au sens ou leur degré de sulfatation est
notamment fonction des efforts mécaniques
réalisés au cours la croissance du cartilage.
Elles sont reconnues par de nombreux récepteurs de
micro-organismes. Elles interviennent dans la croissance
neuronale et dans les interactions entre cellules gliales et
neurones. On connaît des chondroïtines plus ou
moins sulfatées chez de nombreux
invertébrés dont des Echinodermes, des
Crustacés et l'incontournable Cœnorhabditis
elegans. Chez ce dernier on a suivi par
l'intermédiaire des ARN de la chondroïtine
synthase le rôle majeur des chondroïtines non
sulfatées lors de la morphogenèse; celles-ci
s'étant révélées indispensables
aux divisions cellulaires. D'une manière encore plus
nette une chondroïtine sulfate a été
isolée à partir de vésicules de
sécrétion de la couche endodermique des
blastula de l'oursin. Après exocytose, c'est cette
chondroïtine sulfate, très hydrophile, qui
serait responsable du gonflement d'une fraction de la
matrice accolée aux cellules endoblastiques et qui
provoquerait leur poussée vers le centre de la
cavité embryonnaire (voir texte et
figures (35,36 et 37) sur le site de jussieu:
http://www.snv.jussieu.fr/bmedia/oursinMDC/p4gastrula.html).
Pour pousser le raisonnement plus loin on peut aussi faire
appel à des liens qui commencent à être
établis entre stimulus mécanique (ici le
gonflement d'une couche hydrophile de par sa nature
polysaccharidique particulière) et l'activation de
certains gènes (voir par exemple
Emmanuel Farge: «Certains gènes embryonnaires
sont mécano-sensibles», brève de La
Recherche, 369, novembre 2003, p 16-17 d'après E.
Farge, Current Biology, 13, 1365, 2003).
Dans les colorations histologiques de préparations de
cartilage utilisant un colorant (HE) basique (chargés
positivement), ce dernier se fixe
préférentiellement aux molécules
chargées négativement (protéines et
acides notamment). L'intensité de la coloration
reflète assez bien la teneur de la matrice en
protéoglycannes, fortement anioniques. Cette teneur
est plus forte dans la matrice entourant les chondrocytes
adultes que dans celle des cellules souches cartilagineuses
ou chondroblastes. Avec des colorants plus
spécifiques, on peut suivre encore plus finement la
teneur en chondroïtine sulfate des tissus.
De la synthèse de chondroïtine sulfate,
précoce et largement répandue dans le jeune
embryon de la lignée animale à un
métabolisme principal dans des tissus à
vocation structurelle chez les
vertébrés.
On a ainsi pu mettre en évidence par des
méthodes histochimiques que la synthèse de
chondroïtine sulfate n'est pas limitée aux
cellules cartilagineuses mais qu'elle est présente
aux premiers stades du développement, même dans
les ébauches épidermiques qui n'engendrent
jamais de cartilage. Sa synthèse est très
fortement augmentée dans les tissus qui
évolueront en cartilage alors que sa synthèse
est diminuée dans les tissus qui ne donneront pas de
cartilage. On peut aussi relier son niveau de
synthèse aux autres rôles que l'on
découvre progressivement.
R. Chandebois proposait en 1993 (PFD, p134-135) une
audacieuse hypothèse concernant l'évolution
de la formation du cartilage. Elle notait ainsi que tous
les éléments inducteurs (chorde, somites,
endoderme, épiderme...) que l'on retrouve chez les
Vertébrés à squelette osseux et
cartilagineux, sont présents chez l'Amphioxus (qui
n'a cependant ni tissu osseux ni tissu cartilagineux) et
donc susceptibles de déclencher des inductions
nécessaires à la lignée cartilagineuse.
Le vitellus est l'élément mésodermisant
principal chez les vertébrés. Mais chez
l'Amphioxus, dont l'œuf possède cependant une
grande charge en vitellus, c'est une zone en forme de
croissant, riche en mitochondries associées à
du reticulum lisse qui possède les
potentialités mésodermiques. Dans le cadre
d'un hypothèse phylogénétique qui
ferait dériver les vertébrés d'un
ancêtre de l'Amphioxus, on peut imaginer que le
changement radical d'un organisme non cartilagineux à
un organisme cartilagineux ai pu s'opérer en une
unique et brutale modification de la compétence
chondrogénétique des premières
ébauches à l'induction du vitellus. De
même, on a longtemps considéré certains
invertébrés comme très
évolués, notamment du fait de
métabolismes particuliers. C'est le cas des
Céphalopodes parmi les Mollusques, dont certains
membres synthétisent du cartilage. Ce changement
serait aussi à rapprocher d'une évolution
brutale ayant touché les ancêtres des
Céphalopodes au sein d'une lignée
déjà âgée de Mollusques. Dans les
deux cas c'est une évolution brutale sur un terrain
ontogénétique déjà prêt
à recevoir ce changement de compétence. La
synthèse d'une molécule spécifique
à un tissu ne se situe pas en bout de chaîne
d'inductions embryonnaires mais bien au début du
développement, au moment où son
métabolisme est larvé, selon la théorie
de la différenciation par changement dans le
rendement des gènes.
Cette hypothèse, revue à la lumière de
connaissances modernes (voir ci-dessus les différents
rôles des CS), reste cependant intéressante,
malgré une indispensable reformulation. En effet, les
résultats actuels ont mis en évidence que les
très nombreuses chondroïtine sulfates des
animaux - invertébrés compris -
synthétisées dès les premières
étapes embryonnaires ont un rôle majeur dans
les mouvements cellulaires et donc dans la
morphogenèse, comme il fallait s'y attendre de la
part de molécules essentielles de la matrice
cellulaire dont un rôle est, en quelque sorte, de
guider les cellules. Le cartilage est bien un tissu
très spécialisé dans lequel la
synthèse de chondroïtine sulfate devient
prédominante à tel point que cette substance
confère sa principale propriété au
tissu - sa souplesse - de par sa forte hydrophilie. Mais
l'hypothèse phylogénétique du passage
d'une position mineure d'un gène à la position
de gène majeur, garde toute son actualité. Sa
large distribution vient au contraire renforcer
l'idée que c'est bien avec un nombre restreint de
gènes polymorphes et polyfonctionnels que
l'évolution progresse et non par ajout de
gènes de plus en plus nombreux.
|
Peut-on dire, pour simplifier, que si l'évolution
néodarwinienne résulte de l'accumulation de
gènes nouveaux, l'évolution téléologique
résulte, elle, de l'accumulation d'automatismes embryonnaires
?
En acceptant cette formulation, on risque de retomber dans une vision
matérialiste qui réduit l'automatisme à un
programme qui doit bien se trouver écrit quelque part. Or, une
théorie téléologique est fondamentalement
différente de cette vision, car, même s'il elle
n'interdit pas une interprétation matérialiste, elle
est ouverte à un vitalisme. Comme il est écrit plus
haut, les termes de comportement social ou de travail
sont toujours préférables à celui de
mémoire. L'ovocyte
ne mémorise pas les comportements déterminés. On
peut dire qu'il contient la matière, l'information, les
formes et les structures qui peuvent permettre, à sa
réactivation (lors de la fécondation) de conduire les
populations qui sont issues de son développement à
donner naissance à un organisme qui atteigne au moins
l'état évolutif de son ancêtre. Mais surtout
l'ovocyte est lui-même la cellule sexuelle qui, pour un
non matérialiste, dépasse en tant qu'unité
fonctionnelle du vivant, la simple description de son contenu. Pour
un vitaliste, l'évolution est en cela une
continuité d'êtres vivants et non pas une
succession d'êtres vivants qui se transmettent des
éléments héréditaires (ADN,
protéines, structures...). Du point de vue métaphysique
ce passage d'une cellule sexuelle à un nouvel être
vivant est une création nouvelle.
Ajout 2008 : cette
question de la mémoire (qui occulte l'histoire) me
paraît être clairement expliquée par André
Pichot qui l'attribue à un héritage
weismannien (repris dans le darwinisme moderne): voir par
exemple les extraits de l'article:
Hérédité et évolution (l'inné et
l'acquis en biologie),André Pichot, Revue Esprit, juin
1996
A suivre....