De la biodiversité à
l'histoire des êtres vivants
(1ère
partie)
Le chapeau introductif du
programme
à cette partie
nous demande d'aborder la biodiversité, qui est un
problème de classification (nommer et comparer les êtres
vivants), et les phylogénies, qui sont autant un
problème de classification mais qui exigent aussi une
théorie de l'évolution. Le détail des parties
proposées ensuite exclut, par le vocabulaire, toute autre
classification que la cladistique et exclue toute étude de
l'évolution, qui apparaît dans les limites
imposées (même si le mot apparaît dans le titre).
Ceci est à mon avis incohérent.
L'homme sera traité à part car on ne peut traiter de
paléo-anthropologie sans faire d'anthropologie, ce qui n'est
pas uniquement un problème de théorie
évolutive.
retour accueil, plan
du cours de terminale S
Plan de cette page:
Introduction
1. Des classifications
évolutives (partie culturelle, pour clarifier les
bases et faire des ouvertures)
- 1ère étape: le
transformisme
- 2ème étape:
l'âge d'or de l'anatomie et de l'embryologie
- 3ème étape:
le triomphe de la biologie moléculaire
- 4ème étape
(pour les curieux uniquement) :
l'évolution téléologique et le retour attendu
de l'embryologie
Introduction
On admet que la zoologie, la botanique et les branches
spécialistes des divers règnes du monde vivant, non
constituées en disciplines, ont été d'abord
descriptives, dans leur démarche si ce n'est dans leur
histoire : analyser, étudier le plus précisément
possible tous les êtres vivants , est la première
tâche. C'est ce que l'on nomme d'une façon moderne
l'étude de la biodiversité.
(Selon les termes de la "Convention sur la
diversité biologique", signée à Rio de Janeiro
en 1992 par 156 nations: «La diversité biologique
signifie la variabilité au sein des organismes vivants de tous
les milieux, aussi bien terrestres, marins que d'autres
écosystèmes aquatiques, ainsi que les complexes
écologiques dont ils font partie; ceci inclut la
diversité des espèces, entre espèces et celle
des écosystèmes » - traduction
personnelle). Vient ensuite, ou au moins dans un
deuxième temps de réflexion, l'étape des
comparaisons, des explications. Toutes ces sciences se
seraient ensuite fédérées, plus ou moins
facilement, en une unique biologie: une science du monde
vivant qui se veut descriptive mais aussi explicative.
L'explication la plus attractive depuis deux siècles
maintenant est celle de l'évolution: les
êtres vivants forment une gigantesque chaîne d'individus
qui transmettent la vie , conquièrent les espaces fluctuant au
cours des temps géologiques qui se chiffrent maintenant en
milliards d'années, résistent à des cataclysmes
et évoluent au point que la diversité des êtres
vivants du passé dépasse probablement celle des
êtres vivants actuels. C'est l'histoire des êtres
vivants.
Nous verrons donc, en replaçant approximativement les
théories dans un contexte historique, les différentes
classifications en usage actuellement et les théories
évolutives qui y sont éventuellement attachées
(juste citées pour ne pas trop déborder du programme).
Nous prendrons ensuite l'exemple des vertébrés, comme
nous y invite le programme.
Les classifications des êtres vivants
et donc la taxinomie qui en est la science constituée
en discipline, reflètent les changements dans les grandes
théories explicatives et donc aussi dans les philosophies
admises par le plus grand nombre à une époque
donnée. Même si certains peuvent regretter que la
position de la taxinomie ne soit plus dominante, comme elle l'a
été pendant au moins deux siècles. Ainsi, comme
l'affirme François Dagognet, « la taxinomie n'est plus
la recherche, elle est seulement son miroir »
(François Dagognet, E.U., article taxinomie). Certains
éléments conformistes (intellectuellement) de notre
Europe de l'ouest de ce début de XXIème siècle,
assez généralement matérialiste et relativiste,
se sont trouvés une classification à la mesure de leur
philosophie: le cladisme
(ce lien renvoie à une page qui s'efforce de
présenter cette méthode de façon critique).
Mais les tenants d'une classification plus ouverte au sens, moins
idéologiquement marquée, bref, plus biologique, sont
prêts à présenter leur travail. L'adoption par
les programmes d'enseignement du secondaire de la classification
cladiste (sans discussion possible) est une étape majeure qui
risque de sonner le glas des autres visions du monde vivant si les
enseignants n'y prennent pas garde. Enseigner plusieurs
classifications me paraît indispensable et fait partie de la
légitime liberté pédagogique, même si le
jeu est souvent faussé par le dogmatisme des sujets d'examen
contre lequel il faudra sans cesse lutter.
1. Des classifications
évolutives (partie culturelle)
|
1ère
étape: le
transformisme
(ou
évolution
dans le sens moderne) découle de l'intuition
géniale de Lamarck (dès 1802) et
énonce que l'ordre de classification naturelle des
espèces, des plus simples aux plus complexes,
reflète l'ordre historique d'apparition des
espèces au cours des temps
géologiques.
|
C'est en 1818, dans son Histoire naturelle des Animaux sans
vertèbres que Lamarck présente ce qui peut sembler
être le premier arbre phylogénétique,
documenté et non abstrait (La naissance
du transformisme, G. Laurent, Vuibert/Adapt, col. inflexions,
2001, p 88-89).
|
«Ainsi, je soumets à la méditation
des zoologistes, l'ordre présumé de la
formation des animaux, tel que je l'exprime dans le tableau
suivant :
ORDRE présumé de la
formation des Animaux,
offrant 2 séries séparées, subrameuses
»
(Lamarck, Histoire naturelle des Animaux sans
vertèbres, t1, 1815, p. 457)
|
|
|
[1] SÉRIE DES ANIMAUX
INARTICULÉS
|
[2] SÉRIE DES ANIMAUX
ARTICULÉS
|
|
Animaux apathiques.
|
|

Infusoires.
|
|
|
l
|
l
|
Polypes.
|
l
|
|
l
|
l
|
|
|
l
|
|
l
|
l
|
l
|
|
l
|
Radiaires.
|
Vers.
|
Ascidiens.
|
|
l
|
|
l
|
|
|
|
l
|
|
l
|
l
|
|
l
|
|
l
|
Epizoaires.
|
|
l
|
|
l
|
l
|
|
.....................................................................
|
|
Idem
sensibles.
|
|
l
|
|
l
|
l
|
Acéphales.
|
|
l
|
Insectes.
|
|
l
|
|
l
|
l
|
l
|
|
l
|
|
Annelides.
|
l
|
Arachnides.
|
Mollusques.
|
|
|
l
|
|
|
|
|
|
Crustacés.
|
|
|
|
|
|
l
|
|
|
|
|
|
Cirrhipèdes.
|
|
|
.....................................................................
|
|
Idem intelligens.
|
|
|
Poissons.
Reptiles.
Oiseaux.
Mammifères.
|
|
|
Les six tomes suivants de son Histoire naturelle sont
consacrés à fournir ce qu'il appelle les
"pièces justificatives" de sa théorie de la
transformation des espèces. Son ordre de classification des
espèces est l'ordre historique d'apparition des
espèces. Lamarck, philosophe matérialiste, trouve
dans cet ordre, l'expression d'une succession causale qui est celle
de la vie organisée: «... pour
faire exister cette force vitale, et lui donner toutes les
propriétés qu'on lui connaît, la nature n'a pas
besoin de lois particulières; celles qui régissent
généralement tous les corps lui suffisent parfaitement
pour cet objet » (Philosophie
zoologique, 1809, t. 2, p. 96 in La
naissance du transformisme, G. Laurent, Vuibert/Adapt, col.
inflexions, 2001, p 113) : il n'y a plus de frontière
entre la connaissance du vivant et du non vivant; les êtres
vivants supérieurs viennent des inférieurs par
génération et les tout premiers se sont
organisés de façon spontanée à partir
du non vivant (complexification naturelle ou mouvement du vivant),
l'homme lui-même étant un produit de cette
complexification (voir La naissance du transformisme, G.
Laurent, Vuibert/Adapt, col. inflexions, 2001, p 116-118 où
Lamarck est reconnu comme le premier scientifique à proposer
l'origine simiesque de l'homme).
Darwin affirmera ainsi plus tard ce qui était devenu un
lieu commun: «Toutes les règles, toutes les
difficultés, tous les moyens de classification qui
précèdent, s'expliquent, à moins que je ne me
trompe étrangement, en admettant que le système naturel
a pour base la descendance avec modifications, et que les
caractères regardés par les naturalistes comme
indiquant des affinités réelles entre deux ou
plusieurs espèces sont ceux qu'elles doivent par
hérédité à un parent commun.
Toute classification vraie est donc
généalogique; la communauté de
descendance est le lien caché que les naturalistes ont, sans
en avoir conscience, toujours recherché, sous prétexte
de découvrir, soit quelque plan inconnu de création,
soit d'énoncer des propositions générales, ou de
réunir des choses semblables et de séparer des choses
différentes» (On the Origin of Species,
1859, traduction de Barbier, 1896, p. 495, in La naissance
du transformisme, G. Laurent, Vuibert/Adapt, col. inflexions,
2001, p 90).
Darwin est un des premiers à proposer une théorie
explicative de ce qu'il appelle "l'hérédité
des caractères acquis". Cette formulation, que l'on ne
trouve pas chez Lamarck, désigne une vision courante à
l'époque selon laquelle «les êtres vivants
transmettent à leurs descendants les caractères qu'ils
ont acquis durant leur existence par l'usage ». Darwin
proposa comme hypothèse ce que l'on nomme la
Pangenèse: « les caractères acquis
créaient dans le corps des «gemmules» qui
transmettaient ces modifications dans les cellules
reproductrices». (La naissance du
transformisme, Lamarck, entre Linné et Darwin, Goulven
LAURENT, Collection inflexions, Vuibert/Adapt, 2001, p
131)
Toute classification vraie est donc une
généalogie. Toute classification vraie est donc
évolutive. Même si les mécanismes de
cette évolution restent une question ouverte.
|
2ème
étape: de l'unité du plan
d'organisation , définie pour les animaux et
défendue par Geoffroy Saint-Hilaire (vers 1800), on
passe, au milieu du XIXème siècle, à
une période de domination des études
anatomiques et embryologiques, dont l'intuition
d'Haeckel (vers 1874) exprimée par
"l'ontogenèse récapitule la
phylogenèse" peut être un bon exemple.
Les classifications évolutives sont alors
basées sur des caractères morphologiques,
anatomiques et embryologiques, plus rarement
éthologiques, ou écologiques.
|
La notion de plan d'organisation remonte, d'après
mes sources, à Georges Cuvier. La classification que
Cuvier (1769-1832) propose, dans son ouvrage: le Règne
animal (1817), comprend «quatre plans
généraux» sur lesquels tous les animaux sont
construits; ils correspondent aux embranchements des
Vertébrés, des Mollusques, des Articulés et des
Zoophytes. Le travail de Cuvier repose sur l'anatomie comparée
et l'étude des fossiles est associée à celle des
formes actuelles. Mais les faunes présentées sont
séparées par des hiatus (interruptions ou
catastrophes) pour Cuvier qui était fixiste et ne
concevait pas que les espèces dérivent les unes des
autres par transformation (voir
ancien cours de TS). C'est pourquoi
la vision transformiste de l'unité des plans
d'organisation dans le règne animal revient à
Étienne Geoffroy Saint-Hilaire (1772-1844) qui utilisa
les travaux de Cuvier et qui adopta les idées de Lamarck.
Ernst Haeckel (1834-1919) est probablement le plus connu
des transformistes qui ont développé l'idée
suivante: la mise en place embryonnaire des plans d'organisation des
animaux reflète leur mise en place au cours de
l'évolution. Il désigne ainsi, le premier,
l'évolution de la lignée d'un organisme au cours des
temps géologiques, par le mot de
"phylogenèse", devenu des plus courants.
« Les deux séries de développement
organique, l'ontogenèse de l'individu et la
phylogenèse du groupe, auquel il appartient, sont
étiologiquement liées de la façon la plus
intime. ... Comme je l'ai dit alors (Morphologie
générale, II, 110-147, 371),
l'ontogenèse ou l'évolution
individuelle, est une courte et rapide
récapitulation de la phylogenèse, ou du
développement du groupe correspondant, c'est-à-dire
de la chaîne ancestrale de l'individu, et cette
ontogenèse s'effectue conformément aux lois de
l'hérédité et de l'adaptation» . Ces
deux derniers termes faisant référence à la
sélection naturelle : «action
combinée, générale de
l'hérédité et de la variabilité dans
la lutte pour l'existence ».
Sa "loi
biogénétique fondamentale" est souvent
résumée par "l'ontogenèse
récapitule la phylogenèse", même si ce
raccourci est incomplet (voir quelques autres
éléments dans l'ancien
cours de TS, paragraphe a2).
De nos jours de nombreuses critiques sont dirigées vers cette
loi. La plus importante me paraît être la remarque selon
laquelle cette loi est probablement profondément
altérée par des adaptations secondaires, même si
cette observation ne fait que repousser le problème du
déterminisme de cette loi. Pierre-Paul Grassé, affirme:
« Ne lui en demandons pas trop, car dans un grand nombre de
cas l'ontogenèse est profondément modifiée par
des cénogénèses et des adaptations à des
modes de vie très particuliers ; nous pensons aux larves
pélagiques ou simplement planctoniques dont les
caractères, prétendus archaïques, ne le sont
peut-être pas » (L'évolution du vivant,
P.P. Grassé, 1978, Ed. Albin Michel, Paris, p 58).
Il reste que les similitudes de développement embryonnaire
constituent un caractère majeur dans les filiations,
même si on refuse la loi globalement.
Une nouvelle étape à été franchie
dès 1989 par R. Chandebois, une embryologiste qui
intègre ces ressemblances comme une récapitulation,
recommencée lors de chaque développement, de chaque
individu, des innovations successives réalisées dans le
phylum auquel appartient l'organisme. Nous préciserons, dans
un chapitre suivant, cette nouvelle théorie du vivant qui
englobe l'évolution.
Actuellement, le terme de
phylogénie ou
phylogenèse (du grec
phulon = race, et
genèse ou génie) est
devenu absolument équivalent à celui de
classification évolutive, sans
référence à la loi biogénétique
fondamentale de Haeckel, lien, que l'on peut s'efforcer de
rétablir dans ce cours.
Toute classification vraie est donc une phylogenèse et,
pour les animaux doit s'efforcer de laisser transparaître
l'ontogenèse.
Le fait que cette loi soit limitée aux animaux s'explique sans
doute historiquement, notamment par le fait que la biologie, comme
science unifiée, n'en était qu'à ses
débuts. On pourra, pour les besoins de notre exposé,
l'étendre à une compréhension globale du vivant,
du moins pour les animaux et les plantes.
Il faut noter qu'Haeckel publie les premiers arbres
généalogiques de quelques groupes de plantes en
1866.
|
3ème
étape: Vers le milieu du XXème siècle,
la biologie moléculaire, d'emblée
unifiée - elle se pose comme science du vivant qui
dépasse les frontières des règnes -,
permet des rapprochements originaux, qui dépassent
ceux établis par comparaison entre les
caractères observables optiquement. Dès lors
trois écoles de classification , dont deux
sont évolutives, peuvent être
distinguées.
|
La biologie moléculaire est une science unificatrice
puisqu'elle repose sur le postulat que tous les êtres vivants
sont composés des mêmes molécules ou
plutôt, qu'il sont composés à partir d'un nombre
fini de molécules pouvant être étudiées de
façon exhaustive par la chimie.
La molécule la plus couramment citée lorsque l'on
cherche une molécule spécifique du vivant est sans
aucun doute l'ADN. Ce n'est bien sûr pas la seule et surtout
pas la plus abondante (cellulose, collagène et kératine
sont de meilleurs candidats). Depuis que l'on en connaît de
nombreuses formes (formes A, B, Z, puis plus récemment formes
courbes et ADN-H avec un fragment de triple hélice...),
l'unité commence à se fissurer car il est sûr que
la séquence de l'ADN est fort insuffisante à
décrire la richesse que peut receler cette molécule
complexe, notamment dans sa forme tridimensionnelle.
De plus, rien de moins apte à une étude
évolutive qu'une molécule comme l'ADN, fragile, non
conservée chez les fossiles (sauf exception et avec une grande
incertitude sur son origine et donc son âge) et qui doit
transmettre de façon stable les informations
nécessaires aux synthèses protéiques des
êtres vivants. Et pourtant, c'est l'ADN qui porte, socialement
maintenant (voir Du gène comme icône
culturelle, Dorothy Nelkin et M. Susan Lindee, La
Recherche, 311, juillet-août 1998, 98-101 et La mystique
de l'ADN, Dorothy NELKIN et Susan LINDEE, 1998, Belin, Coll.
Débats), l'hérédité et donc
à la fois la stabilité et la variabilité; en un
mot: l'évolution, du moins dans la théorie
évolutive la plus couramment admise (voir plus bas,
encadré sur fond vert).
On distingue actuellement trois écoles classificatoires qui
ont été conservées dans différentes
disciplines: phénétique; cladistique
(ou systématique
phylogénétique) et systématique
évolutive (ou classification
évolutionniste) (pour une approche
philosophique voir par exemple la thèse de philosophie de
Richard Blanchette, Le problème de la classification en
zoologie, Université Laval (Québec), PhD, juin
2002 que l'on peut trouver in extenso au format PDF sur internet
à l'adresse: http://www.theses.ulaval.ca/2002/20168/20168.pdf)
:
- La
phénétique
reste, de l'avis des spécialistes, l'outil classificatoire
le plus efficace EN MICROBIOLOGIE car il permet de
prévoir les propriétés d'un nouvel organisme
une fois ses affinités avec tel ou tel groupe mis en
évidence, ce qui est un but essentiel d'une classification.
|
la
phénétique
|
|
Méthode de classification ne prenant en compte
que les caractères visibles d'un organisme
(phénotype: du grec phenos = visible);
elle remonte au XVIIIe siècle avec le
botaniste Michel Adanson. Avec les techniques de biologie
moléculaire elle s'intéresse bien sûr
maintenant aux caractères moléculaires qui
sont en fait cachés. Le terme
phénétique est devenu l'équivalent
de taxinomie numérique.
La phénétique recherche le plus grand
nombre de caractères possibles (c'est pour cela
qu'on parle parfois de "classification naturelle" mais ce
n'est pas une classification évolutive, même
si l'on peut l'utiliser pour classer des fossiles entre
eux ou des fossiles parmi des organismes actuels). Une
classification phénétique se traduit sous
la forme d'un graphe branchu, un dendrogramme
(ou phénogramme), sans
racine, qui traduit les degrés de ressemblance
entre organismes et non une
généalogie.
La hiérarchisation des caractères repose
sur l'anatomie et l'embryologie et seule l'unité
du monde vivant est prise comme hypothèse de
départ.
|
L'outil informatique est utilisé de façon intensive
par les adeptes de la taxonimie numérique dont les choix
classificatoires sont clairement établis (je renvoie
à des ouvrages spécialisés par exemple
Microbiologie, Prescott, Harley et Klein, De Boeck
Université, 1995, Ch20, p406 et s ). Étant
donné la rapidité des découvertes dans le
domaine des micro-organismes et l'incontestable
supériorité pratique de la méthode
phénétique pour les bactéries, la
classification de référence publiée dans le
Bergey's Manual utilise la phénétique
numérique. Des classifications cladistes ont cependant
été publiées, principalement basées
sur les séquences d'ARNr.
La classification phénétique des procaryotes
propose actuellement 4 embranchements basés sur la
présence ou l'absence de paroi cellulaire et la structure
et la nature chimique de cette paroi (ainsi on a les
Gracilicutes (à la peau fine) dont font partie les
bactéries Gram-négatives, les Firmicutes
(à la peau ferme), dont font partie les bactéries
Gram-postives, les Tenericutes (à la peau tendre),
comprenant les mycoplasmes, et les Mendosicutes, à
la peau présentant des défauts); les
Archéobactéries - voir ci-dessous - font partie des
Mendosicutes.
La plupart des classifications modernes
séparent les Procaryotes en 2 grands groupes:
Eubactéries (du grec eu = bon, vrai et bakterion =
petit bâtonnet), les Archéobactéries
(du grec archaios = ancien),
* Les Eubactéries, ou bactéries vraies,
comprenant la majorité des bactéries,
possèdent habituellement dans leur paroi cellulaire des
peptidoglycanes contenant de l'acide muramique et des lipides
membranaires contenant des acides gras à chaînes
droites estérifiées, comme les eucaryotes, la
plupart de leurs ARNt possèdent de la thymine et l'ARNt
initiateur de la synthèse de la traduction porte la
N-formylméthionine, enfin elles présentent des
séquences nucléotidiques de leur ARN -
sous-unité 16S- caractéristiques (on parle de
signature oligonucléotidique).
* Les Archéobactéries: sont dépourvues
d'acide muramique dans leur paroi (ce qui les rapproche des
eucaryotes), ont des ARNt sans thymine au niveau de certains de
leurs bras (comme les eucaryotes), ont plusieurs ARN
polymérases différentes (les eucaryotes en ont 3),
ont des ribosomes dont les sous-unités sont plus complexe
que ceux des Eubactéries (comme les eucaryotes), ont des
lipides membranaires à chaînes aliphatiques
ramifiées à liaisons éther (à la
différence des Eucaryotes qui se rapprochent des
Eubactéries pour ce caractère), ont une
Méthionine portée par l'ARNt initiateur de la
traduction (comme les eucaryotes) et enfin présentent aussi
une signature oligonucléotidique spécifique. Ce sont
d'incontestables Procaryotes (structuralement,
métaboliquement) mais elles possèdent des
caractères moléculaires qui les rapprochent des
eucaryotes et d'autres qui les séparent
considérablement des Eubactéries.
- Les deux méthodes qui suivent sont
phylogénétiques et reposent donc sur une
l'établissement de groupes apparentés. Elles ne se
réclament pas forcément d'une théorie
évolutive mais supposent toutes deux l'évolution:
- la cladistique
construit, selon une méthode élaborée par
Willi Hennig (Grundzüge einer Theorie der
phylogenetischen Systematik , W. Hennig, 1950), des
cladogrammes qui traduisent graphiquement les
clades ou lignées évolutives construites
à partir de caractères polarisés (dont on
décide s'ils sont primitifs ou évolués
à l'aide d'hypothèses évolutives
paléontologiques et embryologiques - voir encadré
ci-dessous sur fond jaune).
Le cladisme part des
caractères (des organismes actuels et des fossiles) et
s'efforce de les relier phylogénétiquement avec
le plus de rigueur logique possible. Il suppose
l'évolution mais ne favorise aucune théorie
particulière.
Pour une approche critique de
la méthode cladiste voir la page "cladisme".
|
La recherche de parentés
Si l'on remonte suffisamment loin, étant
donné que beaucoup pensent que tous les
êtres vivants dérivent d'un ancêtre
unique et que tous les groupes se sont
diversifiés progressivement, tous les
êtres vivants sont apparentés et tous ont
donc un ancêtre commun. La ressemblance entre
deux individus A et B plus proches entre eux que d'un
autre C (similitude relative de A et B par
rapport à C) s'exprime alors par l'idée
que leur ancêtre commun le plus ancien est plus
récent que l'ancêtre commun qu'ils ont
chacun ou à deux avec cet individu C. C'est
cette notion de ressemblance relative qui me
paraît fondamentalement différente d'une
classification qui cherche à établir des
parentés absolues, entre deux groupes. La
question pour un cladiste n'est pas de savoir combien
A et B se ressemblent mais quels sont les
caractères pertinents qui font qu'ils se
ressemblent plus qu'ils ne ressemblent à C,
autrement dit, en supposant qu'une plus grande
ressemblance s'exprime par une origine commune plus
récente, la question est de savoir quels sont
les caractères pertinents qui permettent de les
établir comme un groupe dérivé
d'un ancêtre commun avec C. Aux recherches de
similitudes, immédiatement associées
à une certaine filiation, non fixée dans
le temps mais supposée, on substitue une notion
de ressemblance relative que l'on chiffre à
l'aide de matrices de caractères, toujours
établie, non pas de façon absolue, mais
relativement à un groupe de comparaison. La
polarisation des caractères d'un groupe se fait
toujours en référence à un autre
groupe et non d'une façon absolue. Si je
compare le groupe mammifères au groupe oiseaux,
je n'obtiens pas la même matrice qu'avec le
groupe amphibiens. Petit-à-petit s'est
éloignée de l'esprit de nombreux
systématiciens, l'idée que l'on puisse
avoir une classification définitive, absolue.
Non pas dans le sens qu'elle puisse résister au
temps, car tout le monde s'accorde à dire que
toute classification est dépendante des
connaissances de l'époque, mais dans le sens
où elle explique l'ordre naturel,
extérieur à l'homme et qui
dépasse en soi la connaissance, qui restera
imparfaite. La classification de Lamarck
reflétait cette soif d'absolu, même s'il
savait son travail perfectible.
|
- la systématique
évolutive construit des phylogrammes
qui traduisent des parentés déduites
principalement à partir d'homologies (ressemblance entre
deux structures qui ont la même position anatomique et
ont la même origine embryonnaire).
Les systématiques
évolutives partent aussi des caractères mais
utilisent différentes théories évolutives
pour les hiérarchiser, non pas dans leur ensemble, mais
en les regroupant (par exemple une théorie
évolutive de la sexualité, une théorie
évolutive de la forme...). Il n'y a pas de
théorie évolutive unifiée pour tout le
monde vivant qui soit utilisée par ces
systématiciens.
C'est la classification évolutive qu'utilisent
majoritairement les botanistes.
En effet, les premières classifications connues de
plantes sont être très anciennes
(Théophraste vers 370-285 avant Jésus-Christ)
mais les grandes classifications modernes
phylogénétiques intégrant des
caractères moléculaires, organiques,
embryologiques, fonctionnels et écologiques dans des
phylogénies sans cesse en discussion restent très
ouvertes.
Pour donner un exemple voici une classification
phylogénétique très explicite due à
Dahgren (1975) qui représente dans un plan les plantes
actuelles qui sont le résultat de l'évolution de
rameaux issus d'une souche (elle-même scindée en
plusieurs rameaux, ce qui représente le caractère
polyphylétique des plantes actuelles, comme on le
perçoit de nos jours). Le temps est donc
représenté verticalement et la diversité
des familles dans le plan horizontal.
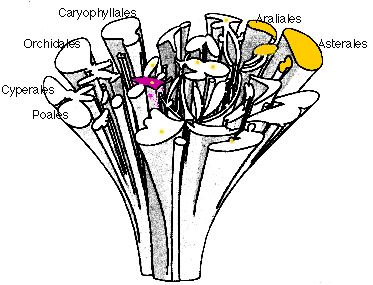
Arbuste phylogénétique de Dahlgren (1975)
d'après La botanique redécouverte, A.
Raynal-Roques, Belin-INRA, 1994, p 92)
présentant les plantes actuelles comme résultat
des différenciations progressives le long de l'axe
vertical du temps
(l'actuel étant représenté par le plan
horizontal supérieur).
Quelques noms d'ordres (suffixe en -ales) sont signalés;
l'ordre des Poales par exemple comprend la famille des
Poacées ou Graminées, et l'ordre
des Asterales, la famille des Asteracées
ou Composées, une des plus évoluées
des familles de Dicotylédones; en
violet
l'ordre des
Magnoliales,
probablement le plus ancien, en
jaune
les plantes chez qui on a
retrouvé des
polyacétylènes
dérivés d'acide gras,
illustrant le fait que cette classification
phylogénétique a été établie
à l'aide de nombreux caractères et permet ainsi
des rapprochements originaux.
|
4ème
étape à l'attention des curieux
(pour
les autres, la suite du cours se trouve à la
page
suivante):
En ce début de
XXIème siècle, les grands traits de l'histoire
des êtres vivants qui émergent de nos
connaissances biologiques (notamment en embryologie) et
paléontologiques convergent vers l'idée d'une
finalité interne (téléologie) de
l'évolution: une évolution
directionnelle dont les étapes majeures auraient
été brusques. Cette nouvelle théorie
évolutive unifiée reste à construire,
dans le détail.
|
Selon les actuels défenseurs de ces idées en
gestation, à qui je souhaite faire une place dans ce cours
(voir bibliographie), la théorie
néodarwinienne de l'évolution s'est
révélée incapable de justifier d'un
mécanisme évolutif responsable d'autre chose que de la
spéciation -mécanisme encore discutable. Il y a du
travail - ce qui n'est pas qu'un jeu de mots - pour fonder une
nouvelle théorie évolutive. La théorie
évolutive nouvelle et unifiée partirait du fait de
l'évolution, et d'une théorie biologique de sa
dynamique, pour faire dériver logiquement les espèces
les unes des autres. Dans une telle phylogénie les
caractères ne sont plus primordiaux, ce qui réduit
l'importance du cladisme comme méthode de classification.
Cette phylogénie repose plutôt sur une histoire de
chaque groupe. Cette théorie est téléologique:
c'est-à-dire qu'elle part de la finalité interne du
vivant. Elle est soutenue par une réflexion philosophique
intense (voir la page
d'accueil et la page
sur les modèles).
finalité
(en philosophie finalité = cause finale: ce pour quoi
un acte est fait)
ou
téléologie
(du grec téléo = fin, but et
logos = parler)
peu importe, mais...
il faut cesser de
traquer le finalisme en biologie.
Le
cours des PE présentait
déjà une discussion autour du finalisme et du
fonctionnalisme à partie d'un article de
l'encyclopedia universalis.
Suite à la parution de l'ouvrage de D. Letellier (La
question du hasard dans l'évolution, L'harmattan,
2002), j'utilise sa mise au point (p 27-29) dans les lignes
qui suivent.
|
|
Une machine à laver possède une
finalité externe dont l'intention doit être
imputée à celui qui a conçu
l'objet.
Un organisme vivant n'a, POUR UN BIOLOGISTE, comme
finalité que lui-même, en tant qu'être
vivant, son auto-organisation, son propre travail de vivant:
c'est une finalité interne. L'adaptation de
l'organisme à son milieu, l'équilibre
dynamique de ses systèmes physiologiques... sont
autant de manifestations de cette finalité interne
qui m'a toujours fait dire qu'il était hautement
préférable d'enseigner aux enfants «
que les incisives étaient faites pour
couper» (ce n'est pas Dieu qui est
supposé ici avoir "fait" les dents, c'est
l'être vivant qui s'est construit au cours de
l'embryogenèse avec des incisives) plutôt que
de dire «que les incisives étaient
utilisées par l'animal pour
couper ses aliments», ce qui est
à mon avis hypocrite car c'est bien l'homme qui nomme
les dents et leur utilisation. L'animal, quant à lui,
vit.
Le terme téléologique sera
employé ici pour désigner une
finalité interne non plus des êtres
vivants, en tant que vie au travail, mais de
l'évolution, en tant qu'histoire des êtres
vivants, résultat du travail du vivant.
|
|
Pour creuser davantage la question, car
tout n'est peut-être pas si simple : Citations et
idées extraites de l'article "Du code
génétique aux codes culturels" de Henri
Atlan, in Encyclopédie Philosophique
Universelle, L'univers philosophique, PUF,
1991):
- sur la définition d'un
programme (puisque la finalité est
appelée à remplacer le programme
génétique moribond) :
« Si programme il y a véritablement, c'est
celui que le biologiste est désormais capable de
se donner à lui-même pour orienter le
développement d'un organisme dans le sens qu'il
souhaite et qu'il peut, dans une certaine mesure,
prédire».
« Les métaphores du langage de la nature sont
trompeuses (...) "le livre de l'univers [est
écrit] dans la langue mathématique"
..."la notion d'information et le programme
génétique..."... il ne s'agit pas dans tout
cela de langages tels que nous les parlons et entendons,
mais de symbolismes opérationnels qui nous
permettent d'agir sur le monde». (voir aussi
le
commentaire sur le programme de
seconde qui rejoint ces
idées en cherchant à remplacer programme
génétique par programme
génétique de travail...).
- la différence entre le
terme de téléologie (intentionnelle)
et téléonomie (non intentionnelle)
établie par Pittendrigh en 1958 (Adaptation,
Natural Selection and Behavior, in Behavior and
Evolution, New Haven, Yale Univ. Press., 390-416) n'a
pas été reprise ici, car le terme
téléonomie a tendance à
désigner la théorie du vivant
proposée dans Le Hasard et la
Nécessité de Jacques Monod (le vivant est
système téléonomique, capable de
morphogenèse autonome et d'invariance
reproductrice). Téléologie, dans cette
page, est pris au sens de téléonomie de
Pittendrigh mais peut aussi dépasser ce
sens...
* «...la seule façon qu'a trouvé
l'éthologie jusqu'à présent de se
développer en discipline scientifique qui ne soit
pas que descriptive, est de s'engager dans la
construction de modèles mathématiques du
comportement. Et tout naturellement la finalité y
apparaît sous la forme de processus à
optimiser, c'est-à-dire de fonctions
mathématiques à maximiser et à
minimiser. Comme en physique depuis toujours, cette
finalité là est acceptable à cause
de son caractère impersonnel et
calculable».
La rationalité causale dans la nature peut
s'exprimer sous la forme du "principe de la raison
suffisante" (c'est la logique fondée sur la
causalité qui régit les
phénomènes naturels), qui se traduit d'une
part par des règles expérimentales
et théoriques (règle
d'objectivité: l'objet doit être
séparé du sujet dans les protocoles
expérimentaux ; et règle de
reproductibilité: les
phénomènes doivent être
reproductibles) et d'autre part par la
nécessité d'un consensus intersubjectif
d'experts sur le rapport des déductions
logico-mathématiques avec les principes
d'identité et de non contradiction.
Mais il existe une voie actuellement empruntée par
certains scientifiques qui acceptent une
intentionnalité comme propriété
émergente des systèmes
auto-organisés, ce qui ajoute des règles
supplémentaires qui sont l'optimisation et
l'adaptation des moyens aux fins (par analogie avec
nos machines) ce qui fait référence
à une science de l'artificiel au sens de H.
Simon.
|
|
« Dans les systèmes
naturels, [...], la notion même
d'auto-organisation implique une origine interne, non
seulement pour les structures mais pour les significations
fonctionnelles des comportements produites par ces
structures. C'est ce qu'avait vu Kant
dans la Critique du jugement, quand il
définissait les organismes vivants par leur
« capacité de finalités
internes ». Dans le contexte vitaliste
de son siècle, il attribuait cette capacité
à une intelligence supérieure orientant les
phénomènes de la vie, de façon
téléologique, c'est-à-dire à
l'aide de causes finales, à la façon d'une
intelligence humaine. Au contraire, les théories
modernes de l'auto-organisation
s'attachent à montrer comment des
structures et des fonctions complexes peuvent être
produites mécaniquement, de façon causale,
à partir de contraintes physico-chimiques locales et
dans certaines conditions d'observation et de mesure. (Henri
Atlan, in EU v12, 2007, article
"auto-organisation)
|
|
Deux
théories évolutives
|
|
La théorie évolutive -couramment
appelée "théorie synthétique"
ou néodarwinisme
- la plus couramment admise en de début du
XXIème siècle repose sur l'hypothèse
d'un programme génétique, stable, qui
transmettait fidèlement à la descendance les
caractères de l'espèce, doublé d'une
variation et une mémorisation
génotypique, dont le mécanisme serait une
accumulation de mutations dans l'ADN. Le sens de
l'évolution serait donné par la
sélection naturelle.
Ces deux premiers points vous ont
certainement déjà été
présentés dans le cours de la classe de
seconde - sans que le nom de la théorie
enseignée ait été cité.
Je me suis
efforcé d'y substituer une vision moins
théorique, plus proche du vivant (voir
cours
de
seconde).
La sélection naturelle n'a par contre pas
été traitée et ne le sera pas,
n'étant pas au programme.
Pour les
élèves que la question intéresse, je
vous renvoie à l'ancien
cours de
terminale.
|
Il existe au moins une autre théorie
évolutive, qualifiée de
téléologique
(du grec téléo = fin, but) car elle
exprime une finalité (ce qui est très
différent d'un programme, voir
ci-dessus). Cette finalité est interne
au vivant. Elle est un automatisme
(que l'on peut encore résumer par un
verbe: survivre). Cette théorie est
issue de l'embryologie animale (notamment
R.
Chandebois), et repose sur
une mémoire cytoplasmique de l'espèce
transmise par l'œuf ou la cellule sexuelle femelle
dans la reproduction sexuée, doublée d'un
automatisme de développement et d'évolution
qui donnerait le sens de celle-ci (évolution
directionnelle). La variation résulterait de
modifications automatiques du fonds cytoplasmique de
l'espèce (responsables des caractères de
l'évolution directionnelle) et de modifications
génotypiques qui permettraient une
évolution adventive.
Cette théorie est
présentée dans une page
annexe. A ma
connaissance, elle n'a pas été encore
enseignée.
|