
Quelques modélisations de couches amphiphiles simples et moins simples....
article "forme", Jean Petitot, Encyclopedia
Universalis; Remarks on the geometry of micelles, bilayers and
cell membranes, Y. BOULIGAND, 1999, Liquid Crystals, vol. 26,
n°4, 501-515; Nouveau regard sur l'origine de l'homme,
Anne DAMBRICOURT-MALASSÉ, La Recherche, 286, avril 1996, 45-51
Remarques:
* J'avais placé ici en 1998 une distinction entre
modèles externes (décrivant le comportement des
observations de l'ensemble d'un système) et modèles
internes (faisant la synthèse des lois décrivant le
comportement de toutes les sous-parties d'un système) à
la suite d'un article de Jacques Oksman dans Pour la Science (258,
avril 1999, 90-95), mais je ne la trouve plus assez pertinente; il
est donc temps de l'abandonner. Même si je l'ai utilisée
notamment dans la description des modèles évolutifs
(ancien cours de terminale) et physiologiques
(pression artérielle). En 2005 je
serai tenté de la reprendre cette fois comme l'expression de
la différence entre les modèles réductionnistes
et structuraux (voir par exemple la page sur
les niveaux d'organisation du vivant).
* 11/2004; j'ai commencé une page
sur la théorie des modèles de René
Thom.... qui s'est
ettofée de deux autres pages (11/2005)...
C'est maintenant la page présente qu'il me faudrait revoir
entièrement mais je remets cela à plus tard.
Les lignes ci-dessous, de Jean Petitot , sont à mon avis le
signe de l'émergence d'un nouveau paradigme (article
"forme" de l'Encyclopedia Universalis; mots signalés,
commentaires et
intertitres ajoutés):
Le paradigme
néodarwinien est insuffisant pour comprendre la genèse
de la forme
(sur ce sujet on peut lire les
deux premiers ouvrages de Rosine
Chandebois et
principalement Le gène et la forme (ou la
démythification de l'ADN), Rosine CHANDEBOIS, 1989,
Ed. Espaces 34 )
« Actuellement, on considère que le
néo-darwinisme - c'est-à-dire la
synthèse de la théorie darwinienne de
l'évolution et de la génétique
moléculaire - fournit un cadre approprié pour la
pensée de la forme en biologie.(...) Or le paradigme
néo-darwinien est un système conceptuel dont
l'apparente «évidence» rend
précisément inintelligibles les
phénomènes morphologiques. Il ne peut que les
attribuer à un hasard évolutif en niant toute
nécessité dans l'ordre des formes, toutes
«lois» de la forme.
Cela est essentiellement dû au fait que, dans ce paradigme, on
identifie subrepticement le concept de contrôle à la
catégorie de cause. Le génome contrôle la forme
et le développement. Son contrôle permet donc de
maîtriser ses effets. Mais cela ne signifie pas pour autant
qu'il n'existe pas de contraintes autonomes et spécifiques
auxquelles doivent satisfaire les formes. En faisant
équivaloir le contrôle génétique à
une cause déterminante, on postule sans plus d'enquête
qu'il n'y a rien à expliquer du côté de ce qui
est contrôlé. Conception historiciste et dualiste, le
néo-darwinisme postule l'existence d'une instance
organisatrice de la matière. C'est un réductionnisme
matérialiste qui admet le primat du fonctionnel,
réduit la connexion structurale et l'organisation
positionnelle des parties à une simple contiguïté
spatiale, et subordonne la «finalité interne»
à la «finalité externe», c'est-à-dire
à l'adaptation et à la sélection. Pour lui, la
structure se réduit à l'hérédité.
Elle est donnée historiquement, et sa seule
nécessité est celle de son évolution. Elle n'est
donc que l'artefact de son contrôle, l'expression
épigénétique de son programme
génétique.
La compréhension de la forme
nécessite une théorie des structures
Le rationalisme structural souligne dans ce point de vue la
difficulté qu'il y a à faire de l'histoire non
seulement la cause de l'évolution, mais également celle
de la stabilité et de l'invariance des espèces.
Pour lui, l'organisme n'est pas seulement un système
physico-chimique complexe génétiquement
contrôlé, mais aussi une structure, c'est-à-dire
une totalité organisée par un système de
relations internes satisfaisant à des «lois»
formelles. Son hypothèse est que l'expression du
génotype par le phénotype demeure
incompréhensible tant qu'on n'introduit pas une information
positionnelle contrôlant la différenciation cellulaire.
Il y aurait dans les êtres organisés une
efficacité de la position, la position
sélectionnant certains régimes métaboliques en
déclenchant certains gènes. Et c'est la
compréhension d'une telle information positionnelle qui
constitue le problème théorique central. Les
caractères principaux des structures organisationnelles sont:
la genèse dynamique, l'autorégulation et la
stabilité structurelle; l'équipotentialité,
à savoir le fait que les structures ne se réduisent pas
à des interactions de composants mais incluent une
détermination réciproque de places, de valeurs
positionnelles; l'équifinalité et
l'homéorhèse (l'épigénotype chez
Waddington), à savoir le fait que le développement
est lui-même structurellement stable comme processus, son
état final étant dans une large mesure
indépendant de son état initial; la clôture des
structures élémentaires et l'existence de contraintes,
de «lois» de la forme; la
«générativité» des formes, l'ouverture
de l'ensemble clos des structures élémentaires vers la
complexité.
René Thom propose un
néo-vitalisme géométrique et
morphologique
C'est pour de telles raisons que René Thom n'a pas
hésité à la fin des années 1960 à
proposer une approche proprement morphodynamique et
structurale des processus de morphogenèse
biologique. Cela a suscité une violente controverse car, ce
faisant, il a réactivé de nombreux thèmes tabous
du vitalisme. Mais il faut bien voir que son néo-vitalisme
est géométrique et méthodologique, et non plus
spéculatif et métaphysique. Le défi est
d'arriver à intégrer les mécanismes
métaboliques cellulaires locaux en un tableau cohérent
de la dynamique globale de l'organisme. Il est d'arriver à
traduire en contraintes pour une générativité
des formes les principes a priori de localité
(la structure comme système de connexions
spatiales) et de finalité (le
contrôle des corrélations par la synergie fonctionnelle
globale). Dans le schématisme morphodynamique, l'a priori
de localité se trouve traduit par la contrainte de
stabilité structurelle impliquant la
généricité des bifurcations des régimes
locaux (des «transitions de phases» métaboliques) et
l'a priori de finalité par l'attribution d'une
signification fonctionnelle à la différenciation
cellulaire, autrement dit par une interprétation
fonctionnelle de la topologie des attracteurs des dynamiques
métaboliques locales. Ce dernier principe semble faire
problème. Il devient pourtant assez naturel si l'on remarque
qu'il est impossible, pour des raisons de principe, d'accéder
en biochimie à des descriptions fondamentales explicites et
exactes, et cela pour deux raisons: parce que les systèmes
différentiels de la cinétique chimique
métabolique sont des systèmes non linéaires
énormes présentant certainement tous les subtils
phénomènes de complexité et de chaos
déterministe que présentent déjà les
systèmes non linéaires simples (attracteurs
étranges, sensitivité aux conditions initiales,
ensembles de bifurcation non stratifiés, etc.); et parce que
la catalyse enzymatique dépend de la configuration tertiaire
des protéines.
Cette impossibilité impose d'en revenir à une
description structurale du métabolisme, par exemple, comme
l'admettent tous les biologistes, en termes de
«cybernétique» et de théorie des
systèmes. Mais on rencontre là un obstacle
épistémologique incontournable. Car on ne saurait se
borner à postuler que, par une sorte d'harmonie
préétablie, la matière vivante est toute
prête à «incarner» une cybernétique
formelle. Au contraire, tout le problème est d'arriver
à comprendre comment peuvent «émerger» du
substrat biochimique et de ses dynamiques métaboliques locales
des structures qui, une fois stabilisées dans leur
activité fonctionnelle, deviennent susceptibles de la
description schématique qu'est la modélisation
systémique. Thom a toujours été
catégorique sur ce point: «L'approche purement
technologique de la cybernétique laisse intact [...]
le mystère de la genèse de l'être vivant et de
son développement au stade embryonnaire et
juvénile.» Ce genre de théories
«soulève de très graves difficultés
dès qu'on veut passer du schématisme abstrait à
une réalisation matérielle dans l'espace-temps»
(R.Thom, Modèles mathématiques de la
morphogenèse, Christian Bourgois, Paris ,1980,
p. 178). Il n'est valable «que pour des
mécanismes partiels, tout montés, et en pleine
activité fonctionnelle». «[Il] ne saurait en
aucun cas s'appliquer à la structure globale des êtres
vivants, à leur épigenèse et à leur
maturation physiologique» (R.Thom, Stabilité
structurelle et morphogenèse, Benjamin, New York,
Edisciences, Paris, 1972, p. 207 ). Il faut donc arriver
à engendrer les cinématiques formelles des descriptions
systémiques à partir des dynamiques métaboliques
sous-jacentes et, pour cela, comprendre de façon
générale comment des descriptions structurales et
systémiques peuvent être associées à des
bifurcations de systèmes dynamiques, à des
singularités de processus, à des
phénomènes critiques.
Le «vitalisme» géométrique et
méthodologique n'a donc rien d'un holisme métaphysique.
Son propos est de ramener à une racine morphologique commune
le biochimique et le cybernétique.
«Écartelée entre ces deux modèles, le
modèle atomique ou réductionniste d'un
côté, le modèle cybernétique de l'autre,
tous deux visiblement insuffisants, la biologie théorique
pourra-t-elle sortir de l'impasse? Le seul espoir d'en sortir est de
reconnaître qu'il n'y a pas de hiatus entre les deux types de
systèmes, et qu'on peut les plonger dans une famille continue
qui les relie tous les deux. Cela obligera à renoncer - au
moins provisoirement - à ce qui fait l'attrait des deux
modèles: l'aspect quantitatif et calculable du premier,
l'aspect diagramme-cybernétique du second. Il faut revenir
à cela seul qui reste commun aux deux types de
systèmes, c'est-à-dire leur extension spatiale, leur
morphologie» (R.Thom, Modèles mathématiques
de la morphogenèse, Christian Bourgois, Paris ,1980,
p. 145). Il faut pouvoir interpréter en termes de
théorie qualitative des systèmes dynamiques et de leurs
bifurcations les concepts fondamentaux de la théorie des
systèmes. L'enjeu est clair, et clairement fondamental.
Il s'agit de dépasser le
conflit entre le physicalisme (primat de la physico-chimie des
substrats) et le fonctionnalisme structural (primat des
schèmes abstraits d'organisation). L'idée
directrice en est que cela est possible à partir de la
mathématisation, en termes de dynamique
générale, du niveau morphologique conçu à
la fois comme tiers terme et comme niveau autonome. Des
phénomènes comme ceux d'induction embryologique ou de
régulation, des problèmes comme ceux des rapports entre
structure et fonction ou de la classification des plans
d'organisation, des concepts comme ceux de champ
morphogénétique ou d'épigénotype exigent
une conception à la fois biochimique et topologique du
formalisme structural qui régit les mécanismes de
l'embryogenèse. C'est l'exigence d'un tel
«mixte» de biochimie et de topologie qui explique
que le prolongement du rationalisme physique à la biologie ne
puisse pas être direct et passe par la schématisation
morphodynamique des catégories structuralistes.»
Même si on fait reposer sa compréhension du vivant
sur une ontologie qui dépasse la logique, les
mathématiques peuvent, comme la philosophie aider à
comprendre le vivant, en profondeur. Les mathématiques par
leur objet (la «quantité des êtres matériels
» ou «la mesure des choses», si l'on utilise un
vocabulaire plus moderne) et leur méthode (abstraction) se
séparent des sciences expérimentales. Mais elles n'en
sont pas moins une vraie connaissance du réel et les
êtres vivants font partie de leurs objets de connaissance.
Le courant signalé ci-dessus me semble particulièrement
favorable à une harmonieuse collaboration entre
mathématiques, sciences expérimentales et philosophie.
René Thom, mathématicien de renom, a
poussé très loin la recherche d'une ontologie nouvelle,
à la fois mathématique et scientifique, qualifié
de réalisme écologique. Le plus connu des
développements de cette pensée est la théorie
des catastrophes.
Si vous n'êtes pas mathématicien et vous souhaitez
quelques éclaircissements sur cette théorie ou des
exemples d'applications (modèles simples mais aussi un exemple
d'utilisation pour l'analyse de données expérimentales
sur les conditions de croissance de la levure Candida albicans
(article
en anglais)), visitez le site de Monsieur
Lucien Dujardin
(http://perso.wanadoo.fr/l.d.v.dujardin/ct/fr_index.html). Les
modèles catastrophiques sont des modèles
mathématiques qui rendent comptent de brusques variations de
la valeur d'un paramètre à l'aide d'une description
géométrique non plane (surfaces plissées...) des
ensembles de valeurs des différents paramètres d'un
système. Cette théorie à été
utilisée dans le deuxième exemple proposé
ci-dessous.
Je vous propose en effet 2 exemples qui peuvent vous faire
comprendre les enjeux de ces avancées:
1 - les membranes biologiques peuvent
être décrites par un modèle
géométrique peu connu, très efficace; ce premier
exemple est du à Y. Bouligand, darwinien convaincu.
2 - les mesures réalisées sur la face interne du
crâne humain à
différents chez l'adulte et lors du développement
embryonnaire mettent en évidence une dynamique propre de
l'homme (contraction crânio-faciale et le basculement de la
partie postérieure du crâne) dont des traces peuvent
être retrouvées sur les os de base des crânes
d'hominidés fossiles, ce qui conduit l'auteur à
proposer une nouvelle dynamique de l'hominisation. Ce deuxième
modèle est du à A. Dambricourt, non
darwinienne.
Remarque:
L'ensemble des travaux de Rosine Chandebois sur l'embryologie
peuvent s'insérer aussi dans ce cadre, à mon avis. Je
vous renvoie, en plus de ses ouvrages cités dans la bibliographie,
à une longue page sur le
développement à l'attention des collègues
enseignant en classe de seconde, où j'ai essayé de
présenter le développement animal en discutant les
références habituelles accessibles aux enseignants.
1. Premier exemple: les membranes biologiques :
Ces données expérimentales conduisent à un modèle global : un modèle de membrane qui tienne compte de tous ces paramètres et qui repose sur la connaissance la plus détaillée de tous les composants de la membrane. Le modèle le plus répandu est le modèle de la membrane fluide que je ne redessine pas ici et qui est dans tous les ouvrages.
Il résulte, pour ce que j'en sais, de la composition de plusieurs modèles issus chacun d'une des techniques ... si l'on cherche à le formaliser mathématiquement, cela devient très vite impossible du fait du nombre de paramètres à prendre en compte
Il existe donc des modèles physiques simplifiés. La première chose que les physiciens font c'est de ne pas tenir compte des protéines : on modélise donc le comportement d'une ou de deux couches de molécules amphiphiles. Le modèle interne le plus courant essaye de prendre en compte les forces entre les molécules amphiphiles pour modéliser le comportement de cette membrane fluide. On en arrive très vite à des équations extrêmement complexes et très lourdes à manipuler.
Existe-t-il des modèles externes ? Oui, on vient de m'en présenter un et je pense qu'il y a là une nouveauté fondamentale : il a été élaboré par Yves BOULIGAND, Professeur à l'École Pratique des Hautes Études, et publié très récemment (Remarks on the geometry of micelles, bilayers and cell membranes, Y. BOULIGAND, 1999, Liquid Crystals, vol. 26, n°4, 501-515). La démarche de Y. Bouligand correspond tout à fait à celle du naturaliste qui ne peut pas matériellement aller rechercher les lois qui gouvernent tous les paramètres de son système et qui se contente d'une loi externe de comportement. Au lieu de chercher à modéliser le comportement des composants de la membrane, il modélise tout simplement la géométrie membranaire et le résultat est surprenant. La géométrie des membranes n'est pas habituellement modélisée en tant que telle, mais simplement décrite à l'aide de deux paramètres reliés aux deux surfaces de la membrane (face externe, face interne) qui correspondent à la vision que nous avons de la membrane comme une superposition de deux couches de molécules amphiphiles (les surfaces considérées étant les surfaces hydrophiles, tournées vers l'extérieur de la membrane). La modélisation d'Y. Bouligand, qui est basée sur la compréhension de la géométrie de la membrane et non sur un modèle préétabli à deux couches, le conduit à envisager tout simplement, non pas deux, mais trois couches : aux deux surfaces citées s'ajoute une surface qui correspond à la couche médiane hydrophobe. Le résultat est magnifique : je ne résiste pas à l'envie de vous recopier quelques-uns des ses schémas qui montrent comment, avec une simple surface supplémentaire on peut décrire une immense variété de formes que l'on retrouve dans le vivant.
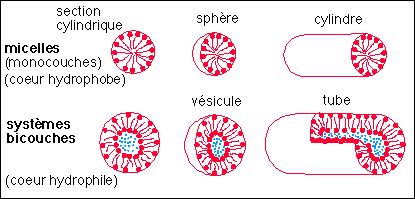
Une "microzoologie" des formes observées et supposées
de micelles "membranaires"
(d'après Remarks on the geometry
of micelles, bilayers and cell membranes, Y. BOULIGAND,
1999, Liquid Crystals, vol. 26, n°4, 501-515,
fig.7)
1 - avec un paramètre pour définir la forme :
sphérique (b), cylindrique (section circulaire : c-d) ou plane
(e)
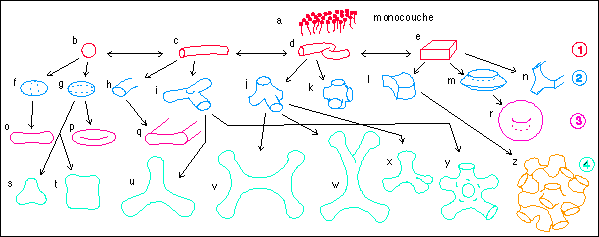
2- deux paramètres sont nécessaires pour définir
les formes : ellipsoïde de révolution (f, g), cylindre
aplati (section elliptique : h), jonction triple (de deux cylindres
avec une dépression centrale, i), jonction quadruple (deux
cylindres accolés latéralement : forme
tétraédrique, j), jonction sextuple (de trois cylindres
s'interpénétrant : forme octaédrique : k), selle
(l), chapeau (m), jonction triple de trois feuillets (n)
3 - trois paramètres sont nécessaires pour
décrire les formes suivantes : double massue (o),
ellipsoïde biconcave (discocyte comme une hématie
par exemple, p), ruban avec côtés cylindriques
(q), stomatocyte (r)
4 - quatre paramètres et plus sont utilisés pour les
formes s à z, on notera la ressemblance par exemple des formes
(y) avec le reticulum et (z) avec la structure d'une
éponge...
Il ne faut pas oublier me direz-vous que l'on est toujours dans un modèle, certes, mais ô combien plus simple que les modèles habituels.
Dans cet exemple on a une démonstration de la complémentarité des deux types d'approche : le modèle interne étant parfois privilégié car il donne une fausse confiance, on croît être plus près de la réalité parce que l'on est plus près de la matière (j'entends par là, plus près des composants non sécables (a-tomes) de la matière). N'y aurait-il pas là encore une illusion matérialiste ? En tout cas le modèle externe permet de prévoir de façon extrêmement simple le comportement d'innombrables membranes biologiques. Il est donc efficace car, prévoir, n'est-ce pas là un but pour tout modèle ?
Il est difficile de bien cerner l'enjeu que représente l'impact de la théorie de Mme Dambricourt. D'autant plus difficile que sa présentation n'est pas si simple. Les lignes qui suivent sont un effort de compréhension et de vulgarisation des idées de Mme Dambricourt, mais qui peuvent comporter des erreurs.
|
La position du trou occipital (foramen magnum) est
un des éléments majeurs que l'on associe
à la station verticale et donc à la
bipèdie des hominidés (qui s'accompagne de
nombreux autres caractères: voir cours
de terminale, paragraphe b.2.3). Cette position est
souvent associée avec un basculement
général de l'encéphale vers
l'avant. |
Le changement de position du trou occipital que l'on montre
à nos élèves de façon évidente en
comparant les vues inférieures d'un crâne d'homme actuel
et d'un chimpanzé est présenté comme
associé à la bipédie. On a même parfois
des raisonnements simplistes d'une coloration lamarckienne
(ce qualificatif est en fait erroné, voir
correction des encadrés de l'ancien
cours de TS :
"l'hérédité des caractères acquis" est
une appellation et une théorie de Darwin pour une idée
alors quasi universellement admise) lorsque les
élèves nous affirment (reprenant ainsi des formulations
maladroites de manuels) que "la station bipède,
nécessaire en milieu ouvert de savanes (des
australopithèques) par opposition au milieu forestier
fermé (des grands singes fossiles, quadrupèdes),
s'accompagne d'un basculement du trou occipital vers
l'avant". Son déterminisme est balayé ensuite d'une
phrase du style "ce basculement résulte d'une(?)
mutation (?) sélectionnée par le milieu
!!!". Essayons de reposer correctement le problème.
Point de départ: la position antérieure du trou
occipital est un trait caractéristique des hominidés.
Il résulte de processus lors de l'embryogenèse que l'on
peut suivre chez l'homme et le comparer aux processus embryonnaires
des grands singes. On peut ensuite s'efforcer de retrouver sa trace
dans des embryons fossiles, si l'on en dispose. Ce qui est le cas, en
un très petit nombre d'exemplaires cependant.
1ère étape: l'enroulement du tube neural présente des caractères propres aux Primates et à l'homme par rapport aux autres Mammifères: la rotation spirale est maximale chez l'homme
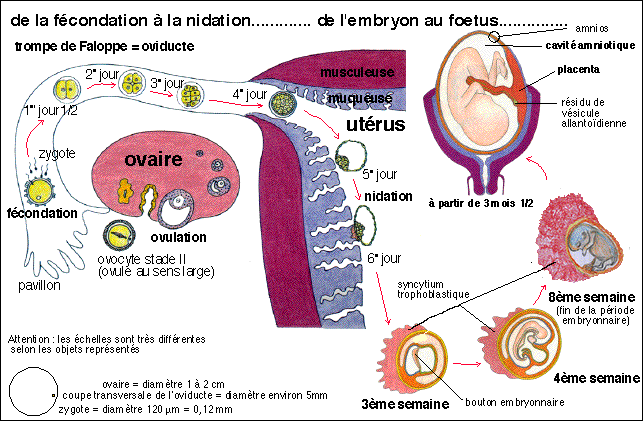
La nidation de l'embryon humain se déroule en même
temps que la gastrulation qui voit la fin de la formation de l'amnios
et du chorion, annexes embryonnaires dont la précocité
de mise en place lors du développement est en relation avec la
particularité des échanges materno-fœtaux par le
placenta (voir par exemple cours pour
les PE et notamment la figure
présentant quelques étapes de la fécondation
à la nidation). Au début de la gastrulation,
l'embryon est réduit à un disque diblastique
résultant de l'affrontement des enveloppes de deux
cavités: l'amnios et le lécithocœle. Vers le
16ème jour de développement embryonnaire on voit
apparaître la ligne primitive qui est d'abord une ligne
virtuelle entre deux zones d'épaississement: les nœuds
postérieur et antérieur (nœud de Hensen). La ligne
primitive se creuse en gouttière et devient
l'équivalent du blastopore. C'est par le nœud de Hansen
(équivalent de la lèvre blastoporale) que va
s'invaginer le tissu qui donnera la chorde et par les bords
latéraux parallèles de la ligne primitive
déjà creusée en gouttière que s'invagine
à son tour le matériel mésodermique. On observe
au cours de la gastrulation un déplacement relatif du
nœud de Hensen vers l'arrière.
La chorde de l'homme est d'abord, comme chez les autres
Mammifères un tube creux (canal chordal) qui s'invagine
à partir de l'ectoderme dorsal de l'embryon (vers le
17ème jour de développement embryonnaire). Ce tube
s'ouvre et se soude provisoirement à l'entoblaste (vers le
19ème jour) pour former une plaque chordale puis
s'isole en une chorde (cordon cellulaire plein). C'est
la chorde qui semble être à l'origine à
l'induction de l'ectoblaste sus-jacent en neuroblaste (plaque
neurale) qui va s'invaginer.
La neurulation, processus essentiel de l'organogenèse des
vertébrés, aboutit à un embryon doté d'un
tube creux ectodermique (tube neural clos vers le 29ème jour
de développement chez l'homme) appelé neurula.
La fermeture de la gouttière neurale chez l'homme est
initiée en plusieurs points et les anomalies de fermeture de
cette gouttière sont assez fréquentes chez l'embryon
(l'anencéphalie résulte d'une absence de
fermeture antérieure et est létale; le spina
bifida est une non fermeture plus ou moins étendue de la
région postérieure dont la gravité dépend
des son extension; l'anencéphalie représente
près de 0,1% des grossesses....). la différenciation ou
régionalisation du tube neural (un peu plus de 30 jours de
développement embryonnaire chez l'homme) termine la
neurulation. On passe progressivement d'un tube présentant
à l'avant trois vésicules (pro-, més-, et
rhombencéphale) à un tube à 5 vésicules
(tél-, di-, més-, mét-, et myencéphale)
auxquelles s'ajoutent de nombreuses dilatations. Le tube
antérieur gonflé est prolongé par la moelle
épinière qui s'allonge vers l'arrière. Chez
l'embryon de poulet, le rôle de la pression exercée sur
les vésicules en formation, par les tissus dorsaux qui
entourent l'étranglement situé à la jonction
encéphale-moelle épinière, a été
mis en évidence expérimentalement: l'absence de
pression provoquant une diminution du volume encéphalique et
une moindre multiplication des cellules nerveuses
(Biologie du développement, S. F. Gilbert,
1996, De Boeck Université, p 254 et fig. 7.10). Les
cellules du tube neural évoluent en cellules nerveuses:
neurones, cellules de la glie (névroglie) et de
l'épendyme.
La partie de l'ectoderme située entre le tube neural et
l'épiderme dorsal forme les crêtes neurales dont les
cellules migrent et donnent naissance au système nerveux
périphérique, aux mélanocytes, à la
médullo-surrénale et à certaines parties
cartilagineuses du squelette crânien dont nous reparlerons
ci-dessous.
La chorde n'atteint pas l'extrémité antérieure
de l'embryon car la multiplication du neuroblaste dans la partie
céphalique antérieure détermine une courbure du
tube neural que l'on observe chez tous les embryons de
vertébrés au moins jusqu'à la fin de la
neurulation (stade bourgeon caudal). Cette courbure n'est plus
visible dans les stades ultérieurs chez les
vertébrés dépourvus de cou.
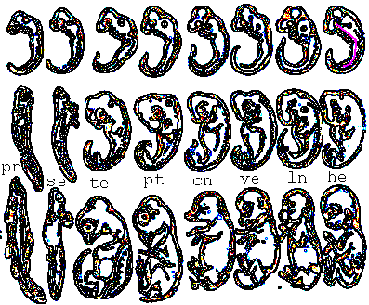
C'est à ce moment là (milieu du deuxième mois de la vie embryonnaire humaine qui se termine conventionnellement à deux mois pour laisser place à un fœtus) que commence un mouvement propre aux primates: la rotation spirale vers l'arrière (rotation selon un axe horizontal perpendiculaire à l'axe antéro-postérieur et dorso-ventral de l'embryon) qui accompagne l'allongement et la fermeture du tube neural vers l'avant.
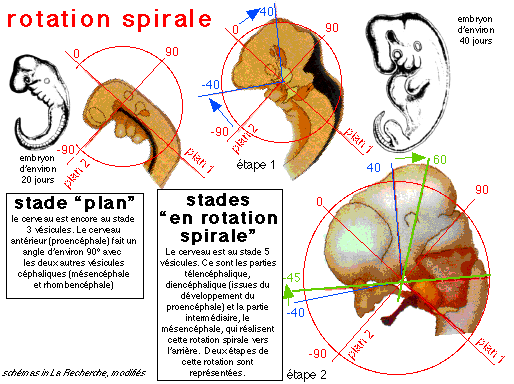
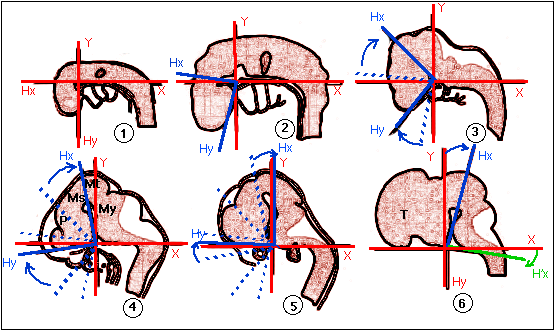
La rotation spirale d'après les schémas
(modifiés) de l'auteur (voir figure sur le site de l'uip:
http://uip.edu/images/ADMFigA.gif)
(légendes: 1= stade 12, 3-5 mm, 26-30 jours; 2 =
stade 13, 4-6 mm, 28-32 j; 3 = stade 14, 5-7 mm, 31-35 j;
4 = stade 16, 8-11 mm, 37-42 j - P=
proencéphale, Ms = mésencéphale,
Mt = métencéphale, My =
myencéphale; 5 = stade 17, 11-14 mm, 42-44 j; 6
= stade 20, 18-22 mm, 51-53 j - T =
télencéphale)
«Au-dessus de la chorde, une partie du cerveau embryonnaire reste plane. Autour de la chorde, du tissu cartilagineux va se former, c'est l'origine des os de la base du crâne qui bordent le trou occipital. La rotation spirale a une amplitude telle que cette partie du crâne bascule à son tour, entraînée par le mouvement général de rotation. Vers la huitième semaine, les tissus cartilagineux qui enchâssent la chorde se redressent et les capsules otiques, qui correspondent aux futures oreilles, basculent. La face inférieure contient un arc cartilagineux qui relie les deux capsules otiques, et qui donne la forme générale de la mandibule. Chez l'homme, à ce moment-là, la partie antérieure de cet arc, qui correspond au menton, se met à former un crochet, comme si elle se redressait, puis laisse apparaître, vu de face, un vide triangulaire. Après les huit premières semaines, la trajectoire se poursuit, tandis que les hémisphères cérébraux commencent à se développer, tout en suivant l'enroulement spiral (Nouveau regard sur l'origine de l'homme, Anne DAMBRICOURT-MALASSÉ, La Recherche, 286, avril 1996, p 49) .
Cet enroulement du tube neural de l'homme se répercute sur
les relations spatiales entre la face et la base du crâne. Mme
Dambricourt pense ainsi que c'est lui qui détermine le
mouvement de bascule du cou des hominidés ainsi que le retrait
du prognathisme (la contraction qui fait que nous sommes les seuls
primates à avoir les dents sous le front) et fait
apparaître le petit vide symphysaire que nous avons tous au
milieu du menton et qui n'existe chez aucun singe actuel ou
passé.
C'est donc un déterminisme embryonnaire qui est à
l'origine de la bascule du cou, ce qui n'est pas du tout impossible
à expliquer dans une théorie darwinienne mais est bien
une donnée embryonnaire (nouvelle).
2ème étape: la signature du déterminisme embryonnaire, et donc l'amplitude de la rotation spirale, se retrouve dans les relations topologiques entre les os du crâne de l'adulte et de l'enfant: un repère octaédrique (nommé pantographe par Mme Dambricourt) permet d'en suivre les variations au niveau du crâne et de la mandibule.
Le passage de l'enroulement du tube neural à la topologie des différents composants du crâne et de la mandibule est évidemment complexe. D'autant plus que l'ossification crânienne est mixte (à la fois dermique et enchondrale, voir plus loin pour une définition de ces mots). Pour expliciter cette relation voir l'annexe en bas de page (in E. U. et "Biologie animale, les Cordés, anatomie comparée des Vertébrés", Beaumont et Cassier, Dunod Université, 1987)
On en arrive maintenant aux relations topologiques entre les différents os du crâne et de la mandibule.
Un pantographe est un petit appareil composé de tiges rigides dont l'assemblage ingénieux (les tiges sont assemblées au moyen d'axes verticaux permettant une rotation de celles-ci dans un plan unique) permet de reproduire fidèlement une figure en agrandissant ou en réduisant son échelle. L'utilisation de ce terme est probablement voulu par l'auteur pour mettre l'accent sur les relations plus ou moins fixes qui existent entre les différents points ayant servi à définir les pôles de la figure qui est une superposition de triangles. La figure la plus stable dessine un octaèdre qui est qualifié de pantographe.
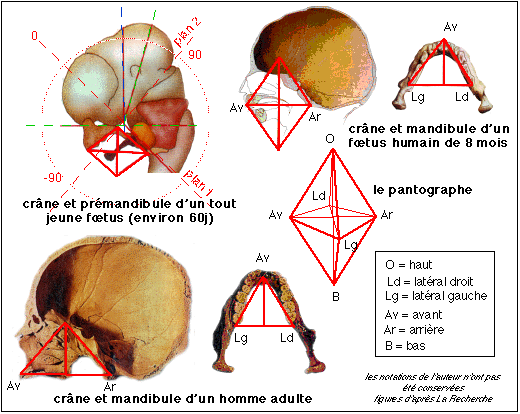
«On retrouve la trajectoire [de l'enroulement spiral] au niveau de la mâchoire inférieure osseuse, dans les trois plans de l'espace. On peut alors modéliser le développement de la base du crâne, selon deux plans. Au pantographe tracé de profil, on ajoute un pantographe transversal qui tient compte de la future arcade dentaire, et de l'orientation de la base du crâne cérébral quand on regarde par en dessous. A la naissance du bébé humain ce double pantographe est très fermé dans le plan vertical et très ouvert dans le plan transversal. Il montre comment les os viennent se positionner plus ou moins sous le cerveau » (Nouveau regard sur l'origine de l'homme, Anne DAMBRICOURT-MALASSÉ, La Recherche, 286, avril 1996, p 49).
L'idée de Mme Dambricourt est donc ici d'utiliser le double pantographe pour suivre les caractéristiques des processus de croissance à l'œuvre chez le fœtus puis chez l'enfant. La découverte consistant dans l'idée qu'un même processus est à l'œuvre chez tous les mammifères et à fortiori chez les humains. L'outil mathématique proposé pour décrire ce processus est un attracteur harmonique. Cet outil est peu connu des non spécialistes et je renvoie à la bibliographie spécialisée pour plus de détails. Dans le vocabulaire courant le terme d'attracteur renvoie bien à une structure stable structurante. Le terme harmonique s'oppose à chaotique étant donné sa transmission héréditaire stable.(voir page sur les modèles de René Thom)
«Tout au long de cette ontogenèse, c'est le même attracteur que l'on voit à l'œuvre. Il procède par contractions successives. J'ai nommé cette trajectoire contraction crânio-faciale. Mémoire ontogénique qui répète inlassablement depuis des milliers d'années, réitérée chez chacun d'entre nous, chez les six milliards d'hommes actuels, dont elle transcende la singularité individuelle, c'est l'attracteur fondamental Sapiens. Attracteur harmonique, il ne perd pas la mémoire de sa propre identité, malgré les milliards de bifurcations qui suivent la fécondation. Les déséquilibres infantiles qui surviennent actuellement montrent qu'il est présent en oscillation au point de rompre des corrélations fonctionnelles. On ne constate pas de tels désordres dans les cimetières des époques historiques» (Nouveau regard sur l'origine de l'homme, Anne DAMBRICOURT-MALASSÉ, La Recherche, 286, avril 1996, p 49).
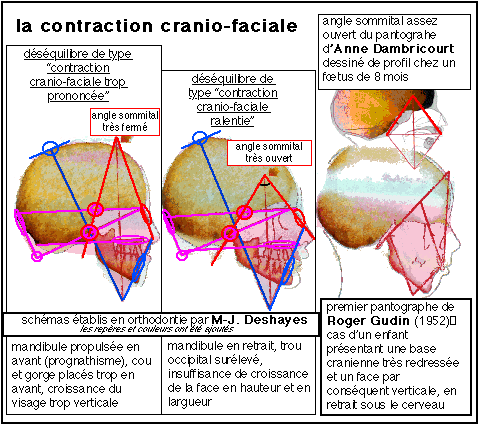
En orthodontie, où plutôt en orthopédie dento-maxillo-faciale, il a été découvert une fenêtre dynamique, c'est-à-dire une période pendant laquelle une trajectoire de contraction en déséquilibre est réorientable. Cette fenêtre permet donc d'intervenir en orthopédie dès l'apparition du déséquilibre.
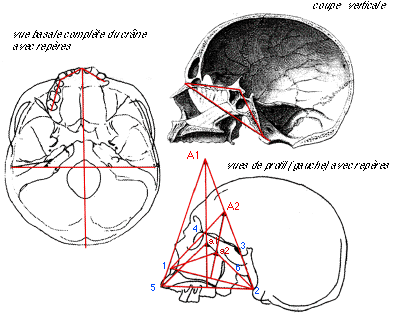
Je suis bien conscient qu'il manque plus que des détails
sur la construction de ce double pantographe.
L'ultime étape de la démonstration de Mme Dambricourt
consiste, à partir de l'observation des proportions des
éléments du double pantographe chez le fœtus,
l'enfant et l'adulte, à énoncer une loi:
|
Cette loi traduisant une dynamique embryonnaire de contraction crânio-faciale pouvant être modélisée à l'aide d'un attracteur harmonique. |
Je ne peux dire que peu de choses sur cet attracteur harmonique,
n'étant pas compétent dans ce domaine et n'ayant pas
accès aux publications spécialisées auxquelles
l'auteur renvoie (L'hominisation et la théorie des
systèmes dynamiques non linéaires (Chaos), 1992, in
Revue de biologie mathématique, 117-119;
Dambricourt-Malassé, A., Les attracteurs inédits de
l'hominisation. Ontogenèse fondamentale, attracteurs
chaotiques et attracteurs harmoniques, 1993, Acta
Biotheorica, 43; ). L'auteur précise que cette
formalisation de sa théorie est en cours mais qu'elle est loin
d'avoir trouvé une solution satisfaisante. Elle s'appuie sur
certains résultats de René Thom tout en
précisant que ses résultats ne sont pas
intégrables à une théorie du chaos ou à
la théorie des structures dissipatives de Prigogine.
« La contraction crânio-faciale est un constat
angulaire. On peut écrire le comportement d'un angle y en vue
transversale, fonction d'un autre angle x en vue de profil et
calculer l'équation y=f(x). On peut le faire à chaque
embranchement fossile qui correspond à une nouvelle
équation. Formaliser l'évolution crânio-faciale
sur soixante millions d'années, c'est tout simplement
écrire l'équation de l'évolution des
équations qui dérivent l'une de l'autre. C'est une
analyse qui consiste à regarder l'évolution de la pente
des équations, on écrit f(x) ou quelque chose comme
y=ax+b (très simplifié). La pente c'est la valeur a.
La représentation devient abstraite, ce ne sont pas des
droites, mais des figures que l'on appelle
attracteurs ou
espace des phases. Et
l'équation finale permet de dégager une loi
mathématique, sans nécessairement connaître la
réalité physique qui lui correspond
» (La légende maudite du XXème
siècle, l'erreur darwinienne, A. Dambricourt, La Nuée
Bleue/DNA, Strasbourg, 2000, fig. 1, 39) .
Remarque:
pour des lecteurs souhaitant avoir une idée de la
théorie des catastrophes dont René Thom a
été le précurseur et au sein de laquelle ont
été développées les notions
d'attracteurs, je renvoie au site de Monsieur
Lucien Dujardin
(http://perso.wanadoo.fr/l.d.v.dujardin/ct/fr_index.html); voir
aussi sur ce site une page sur les
modèles de René Thom avec la définition d'un
attracteur et une page sur les
modèles continus présentant l'intérêt
du travail dans l'espace des phases.
Pour ce qui est de la validité de la loi pour les autres
groupes; la rotation spirale du tube neural existant chez tous les
Mammifères, elle est bien évidemment présente
chez tous les Primates. Seule l'amplitude de flexion-contraction est
variable selon les espèces. Elle reste toujours
inférieure à celle de l'homme.
La position du trou occipital, directement lié à
l'amplitude de cette rotation spirale, est habituellement d'autant
plus en avant que la contraction est forte (minimale chez les
nouveau-nés de lémuriens, intermédiaire chez les
petits singes, plus importante chez les grands singes, maximale chez
l'homme). Cependant la contraction crânio-faciale peut, par
exemple dans le cas du Tarsier, être légère,
malgré une flexion importante du tube, ce qui donne une
position antérieure du trou occipital malgré une face
peut contractée (mandibule en V très pointu par
exemple). Dans cet exemple la cause en est au développement
très exceptionnel des capsules optiques.
La loi semble donc, être valable pour les Primates, sauf
exception: l'amplitude de la rotation spirale du tube neural
détermine la flexion et donc la position du trou occipital et
la contraction-cranio-faciale des lémuriens à l'homme
en passant par les petits puis les grands singes.
3ème étape: application de la loi à l'analyse des fossiles crâniens et mandibulaires dans la lignée des hominidés.
L'hypothèse est que cette loi «perdure et se réitère depuis l'apparition des primates, il y a 60 millions d'années », on peut alors « situer les déséquilibres faciaux des enfants actuels dans une perspective évolutive à grande échelle ».
L'examen de la base des crânes et des mandibules fossiles dans la lignée humaine a conduit l'auteur à présenter ses résultats sous la forme d'un figure majeure (La Recherche, 286, p 46-47, figure sur le site de l'uip: http://uip.edu/images/ADM fig B copier.gif ou dans La légende maudite du XXème siècle, l'erreur darwinienne, A. Dambricourt, La Nuée Bleue/DNA, Strasbourg, 2000, fig. 2, 22 qui est reprise, légèrement modifiée ci-dessous) où apparaissent des paliers de contraction (qui correspondent à des regroupement d'espèces paléontologiques présentant les mêmes types de flexion-contraction crânio-faciale). On notera que l'homme actuel forme ainsi un genre à part, nommé Sapiens par l'auteur.
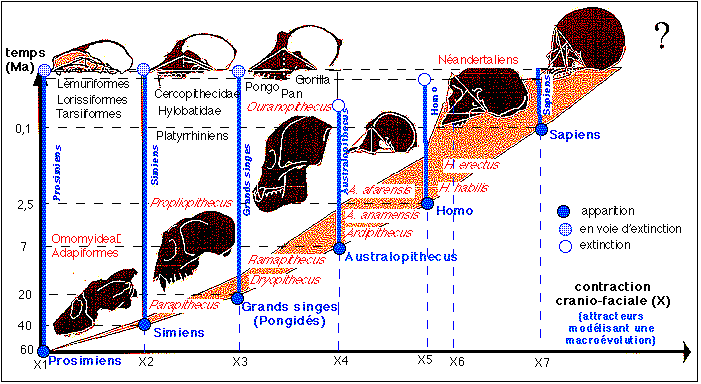
Exemple des australopithèques robustes
(Paranthropes)
« La quête de l'ancêtre commun n'a plus la
même signification. Les ancêtres sont connus.
L'ancêtre des Australopithèques c'est le fond
embryonnaire grand singe que l'on voit depuis 20 millions
d'années. L'ancêtre des hommes archaïques c'est le
fonds embryonnaire commun à toutes les espèces
d'Australopithèques. Ce que nous ignorons en revanche, c'est
comment l'attracteur reste stable, comment l'embryon se
réorganise, et comment la dynamique se complexifie. (...) Avec
ou sans effondrement de la Rift Valley et la disparition de la
forêt, l'Australopithèque est possible parce qu'il
existe un processus qui permet d'une manière ou d'une autre de
complexifier l'organisation d'un ontogenèse. Ce qui est en
cause, c'est la conservation de ce processus, autrement dit sa
transmission. A partir d'un certain moment, le psychisme devient sans
doute lui-même un paramètre. Lui aussi en rapide
évolution, il peut influencer l'aire de répartition de
la nouvelle ontogenèse fondamentale. La population en vient
à sélectionner son milieu par des actes
déjà volontaires. Plusieurs fossiles observés
entre les Australopithèques et les premiers hommes modernes
suggèrent des évolutions ontogéniques
chaotiques, qui ne conservent pas la mémoire de cette
dynamique évolutive : ces fossiles émergent de logiques
inattendues.
On le voit avec les paranthropes, vers 2,4 millions d'années.
Ils ont un trou occipital placé plus près de la face,
mais celle-ci n'est pas sous le front. On ne constate pas de
régression du système masticateur, comme chez les Homo.
Pour Gilles Berillon, il n'est pas impossible que l'on soit là
en présence de deux macroévolutions. On peut tenter une
interprétation. Il faut se représenter une vaste
populations d'embryons d'Australopithèques, oscillant
soudainement autour de l'équilibre crânio-facial et
céphalo-caudal. Différents points de bifurcation ont
donné les embryons qui deviendront des paranthropes d'un
côté, des hommes de l'autre. Sans doute très
légèrement différentes dans leurs trajectoires
embryonnaires crânio-faciales, ces deux amplifications donnent
deux évolutions ontogéniques qui divergent totalement.
l'une retrouve la logique évolutive à l'origine des
Australopithèques, l'autre innove de façon
imprévisible, mais va s'éteindre. Celle en laquelle on
reconnaît le processus, est aussi celle qui donnera Homo
sapiens.
Il faut raisonner en termes de systèmes dynamiques
éloignés de l'équilibre. On admet une
populations d'embryogenèses qui oscillent: quelques solutions
sont viables, la sélection agit tôt sur les
évolutions embryonnaires trop chaotiques, puis les
paranthropes disparaissent. On remarque alors que les
ontogenèses sélectionnées sont celles qui ont le
potentiel évolutif de l'hominisation. Chez le paranthrope, la
base du crâne est contractée, mais selon des
réajustements différents des hommes,
l'ontogenèse retombe dans un ancien attracteur qui devient
dominant, avec le fort développement de l'appareil
masticateur. La face supérieure est presque
désolidarisée du cerveau, un petit goulot osseux les
rattache. Les hommes ont gardé au contraire l'attracteur
neural beaucoup plus longtemps, indiquant une
cérébralisation fœtale plus importante, tandis que
l'attracteur facial masticateur est retardé.»
(Nouveau regard sur l'origine de l'homme, Anne
DAMBRICOURT-MALASSÉ, La Recherche, 286, avril 1996,
53)
Dans ces lignes Anne Dambricourt cherche à expliquer les
mécanismes mêmes de l'évolution, en se
référant aux hypothèses de Gilles Berillon;
c'est une partie moins convaincante car elle pose plus de questions
qu'elle n'apporte de réponses. On est encore loin d'un
modèle global des mécanismes évolutifs non
darwiniens. Quelle sont les causes de la variabilité des
embryogénèses qualifiée d'oscillatoire ? Des
erreurs de développement, des hétérochronies
accidentelles ? (ce que peut nous faire penser l'usage du terme
"chaotique"). Pourquoi l'évolution mémoriserait-elle
des trajectoires nouvelles ? Parce que toute trajectoire d'un
individu est mémorisée pour ses propres descendants ?
(et dans ce cas il n'y aurait pas de mémoire de
l'espèce mais une seule mémoire individuelle, ou au
moins une mémoire de l'espèce
individualisée).
Perspectives
Comme souvent c'est à sa fécondité que l'on
pourra juger de l'importance de cette théorie. De jeunes
chercheurs du Muséum de Paris se sont déjà
attelés à la tâche de rechercher d'autres signes
d'une dynamique embryonnaire étendue à tout l'axe
crânio-caudal et conservée au cours de
l'évolution.
Madame Dambricourt poursuit sa réflexion
paléontologique:
« Il est manifeste que nous sommes là en
présence de déterminismes internes ontogéniques
très puissants, indépendants du milieu.(...) Qu'est-ce
qui a évolué exactement ? On voit que de nombreuses
espèces actuelles ont la même racine embryonnaire que
les espèces fossiles. Il existe donc des fonds embryonnaires
communs qui traversent les espèces et les époques. Cela
m'a conduite à définir une unité biologique, que
j'appelle ontogenèse fondamentale. C'est la trajectoire
embryonnaire commune que partagent certaines espèces actuelles
et fossiles. Voilà l'unité qui aurait
évolué des premiers Primates aux
Australopithèques, puis aux hommes modernes. Nous sommes loin
des conceptions actuelles qui font de l'Australopithèque, ou
de l'homme, des espèces de grands singes bipèdes,
autrement dit des espèces de la mémoire embryonnaire
grand singe.(...) Nous ne sommes plus en présence d'une
évolution adaptative. C'est l'ontogenèse fondamentale
d'un grand nombre d'espèces qui élève son niveau
d'organisation. (...) La sélection naturelle conserve
cette dynamique, elle n'est pas la dynamique elle-même.(...)
Ainsi conçue l'hominisation ne se
fait pas graduellement par petites retouches d'un unique plan
embryonnaire initial comme dans le schéma traditionnel. C'est
bien de macroévolution qu'il s'agit, d'évolutions
discontinues de plans d'organisation ». (Nouveau
regard sur l'origine de l'homme, Anne DAMBRICOURT-MALASSÉ,
La Recherche, 286, avril 1996, 52-54)
|
Le crâne de l'homme résulte
principalement du développement d'os
dermiques qui se construisent sans passer par la
formation d'un crâne embryonnaire
cartilagineux, processus dit enchondral. A sa base
et sur ses côtés, cependant, les os
enchondraux dominent. Dans la série des
vertébrés on observe une nette
tendance à la réduction de
l'endocrâne et à la fusion des os qui
le composent, ce qui rend parfois difficile les
homologies, qui restent donc parfois à
l'état d'hypothèses. On se base
pourtant sur ces homologies pour affirmer que des
processus embryonnaires donnant des os homologues
(même origine embryonnaire, même
position anatomique et même fonction, voir
discussion)
indiquent une filiation évolutive forte. Un
caractère embryonnaire mis ainsi en
évidence a une importance majeur dans une
phylogenèse, qui dépasse de loin des
ressemblances moléculaires (voir cours
de terminale). Dans le cas d'os enchondraux la
filiation est plus simple à mettre en
évidence que dans le cas d'os dermiques, du
fait de la présence d'un modèle
cartilagineux avant ossification, ce qui nous
facilite la représentation d'une forme par
l'imagination. Mais cette distinction n'est
probablement qu'un leurre. En effet, les deux
processus embryonnaires résultent de
mécanismes histologiques semblables: la
genèse de la forme y prend place de la
même façon par les interactions entre
populations (voir par exemple la
théorie
de Rosine Chandebois, paragraphe
2.3.2). Le crâne des Vertébrés se compose de 2 parties: le neurocrâne , qui protège et soutient l'encéphale et les organes sensoriels céphaliques (olfactifs, visuels et stato-accoustiques), et le splanchnocrâne , qui entoure la cavité buccale et pharyngienne. Le tissu lâche (mésenchyme) qui fournit les tissus cartilagineux et osseux des vertébrés est soit d'origine intermédiaire (mésoderme) soit d'origine externe (ectoderme), soit d'origine mixte. Lors du développement embryonnaire, le crâne se construit d'abord par la formation d'un chondrocrâne, cartilagineux, qui est le crâne définitif et unique des Cyclostomes (comme la lamproie) et des Chondrychtiens (comme les raies ou les requins). 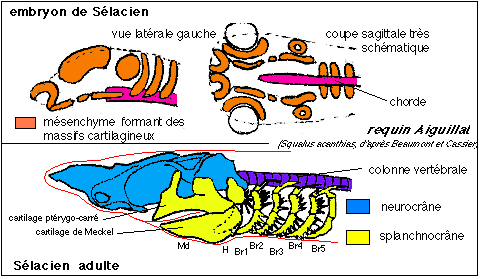 Mise en place des massifs cartilagineux allant former le neurocrâne chez un Chondrychtien (requin aiguillat); on notera que les massifs ont une disposition paire de part et d'autre de la chorde. Chez l'adulte les cartilages issus du mésenchyme formant le chondrocrâne (crâne cartilagineux) qui regroupe le neurocrâne (qui protège et soutient l'encéphale) et le splanchnocrâne (qui entoure la bouche et le pharynx). La disposition métamérique (par segments) du splanchnocrâne est rappelée par la dénomination des arcs viscéraux, primitivement au nombre de 7 paires: mandibulaire (Md), hyoïde (H), 5 arcs branchiaux (Br1 à Br5). Seuls les 2 premiers arcs sont articulés sur le neurocrâne. Chez les autres Vertébrés un crâne
osseux
(ostéocrâne)
apparaît soit aux dépens du chondrocrâne
primitif, qui est partiellement au moins remplacé par
un ostéocrâne enchondral , soit est
ajouté par ossification dite dermique
(ostéocrâne dermique). 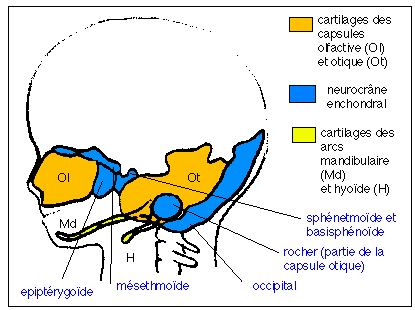 Coupe schématique d'un crâne de fœtus humain (vers 12 semaines de vie embryonnaire ce qui correspond à un fœtus replié d'environ 80 mm de longueur) montrant la réduction du neurocrâne enchondral (endocrâne); en bleu. Le sphénoïde humain est
considéré en anatomie comparée comme
résultant de la fusion embryonnaire de 6 os
(dont 4 par paires) : 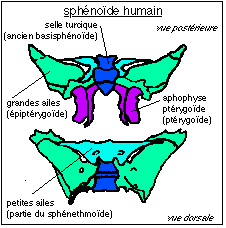 Sphénoïde humain (in Beaumont et Cassier) La mandibule (mâchoire
inférieure) des Mammifères diffère de
celles des autres vertébrés à
mâchoire (Gnathostomes) car elle est constituée
par un seul os dermique (le dentaire) qui s'articule
directement avec le squamosal du toit dermique: c'est
l'articulation mammalienne qui s'oppose à
l'articulation dite reptilienne (entre le carré,
faisant partie de l'ostéocrâne enchondral du
complexe palatin ; et l'articulaire, os enchondral de la
mandibule). 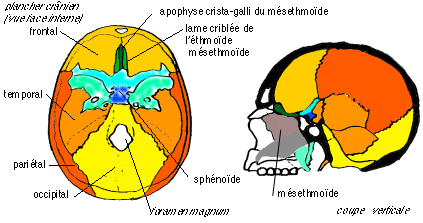 Crâne humain adulte. Les deux schémas du haut (en couleurs) présentent le sphénoïde en place (d'après Beaumont et Cassier) .
|
{kind=link}
{kind=link}
{kind=link}