 |
Planches II et III
(Entre les pages 270 et 271). |
|
plan de cette page |
|
|
page suivante |
|
|
dernière page |
|
|
Planches II et III
(Entre les pages 270 et 271). |
* Ce chapitre est un chapitre de paléontologie c'est-à-dire d'histoire des êtres vivants, ce n'est donc pas un chapitre de science expérimentale. «Paléontologie» vient du grec "paléo"=ancien, "ontos"=l'être et "logo"=parler. Le terme d'«histoire» est pris au sens de "science des événements passés" et non dans le sens restreint des "événements ayant eu lieu depuis l'avènement de l'écriture", les périodes plus anciennes étant désignées sous le vocable de préhistoire. Avec le terme de "paléontologie", on définit à la fois l'objet (les êtres vivants du passé) et la méthode (historique au sens large). Il me paraît nécessaire d'insister sur le fait que nous ne sommes plus dans une science expérimentale mais dans une science historique. Voir pour une ébauche de discussion à ce sujet les pages sur la méthode .
* Ce chapitre est un chapitre de
paléoécologie: il n'étudie ni les
êtres vivants seuls, ni la terre vue comme un milieu sans
vie.
Ce n'est pas un chapitre de géologie, même si nous
utilisons les résultats de la stratigraphie, de la
géochimie...Ce n'est pas non plus un chapitre de biologie,
même si nous utilisons les résultats de l'embryologie,
de l'anatomie, de la
cytogénétique... Il s'agit
de "biogéologie"...En fait nous allons faire de
l'écologie
historique plutôt que de la géologie
historique. C'est pour cela que j'ai cité en bibliographie des
ouvrages d'écologie générale qui tous deux
possèdent une partie historique qui cadre tout à fait
avec notre programme. En fait ce chapitre traite réellement de
l'histoire de la terre et de la vie et donc de
l'évolution. L'évolution (au sens
"évolutionniste".. nous préciserons ces mots mais
j'entends par là une théorie de l'évolution
du vivant) n'est pas, à mon sens, l'évolution des
êtres vivants sur terre, mais bien l'évolution du
système terre-êtres vivants qui, à mon
avis encore, ne forme qu'un seul système. Séparer la
terre des êtres vivants est un artifice, qui peut être
utile mais qui devient un obstacle pour les naturalistes
("biogéologues") que nous sommes . Je reprends par
exemple les mots de R. Barbault, dans son précis
d'écologie générale : «...la
biosphère recouvre partiellement les trois grands
compartiments qui composent la terre, la lithosphère,
l'hydrosphère et l'atmosphère. ... certains
écologues préfèrent utiliser dans ce cas le
terme d'écosphère... Que l'on
parle d'écosphère ou de
biosphère il est clair que les processus
écologiques lient étroitement les organismes vivants
à leur environnement physico-chimique. Dissocier les
êtres vivants de leur milieu est, pour l'écologue,
dénué de sens. En écologie, le terme de
biosphère est toujours utilisé dans un sens fonctionnel
(et non descriptif) ; sa signification est bien celle donnée
au concept d'écosphère.» La terre (globe
terrestre avec tous ses composants, vivants ou non, dont l'homme fait
bien évidemment partie), telle que nous la connaissons, n'est
pas peuplée, elle est vivante, c'est un lieu de vie (je joue
un peu sur les mots mais je veux simplement insister sur cette
profonde interaction terre-êtres vivants).
Remarque:
Je pense qu'il est intéressant de parler ici
rapidement d'une théorie dont certains refusent de parler mais
qui n'est peut-être pas inintéressante :
considérer la terre comme un être
vivant.
|
(voir par exemple l'interview de Peter Westbroek : «la terre est-elle un superorganisme ?», La Recherche, 295, février 1997, 100-101 ; ou encore l'"encyclopédie Hachette multimédia : Science interactive" à l'article "Vie"...) |
|
|
L'hypothèse |
|
|
Auteurs |
L'idée semble avoir été reprise récemment par James Lovelock sous le terme de Gaïa (Les âges de Gaïa, Lovelock J., 1990, Robert Laffont) mais, comme ce nom est très ancien (philosophies pour lesquelles je n'ai fait aucune recherche) et a été utilisé par des non scientifiques avec des visées très différentes ("philosophie" new age ?), le groupe de scientifiques qui a repris l'idée préfère que l'on parle de l'hypothèse Lovelock ou de géophysiologie. Une société de géophysiologie s'est constituée autour de Peter Westbroek en avril 1997 à Oxford... |
|
Critique personnelle |
Pris comme une hypothèse
scientifique, ce concept a des implications que certains
considèrent comme très intéressantes
sur notre compréhension des phénomènes
biochimiques et biophysiques mais pour l'instant il n'y a
pas de développement très poussé. Il
est évident que de nombreux scientifiques
considèrent que c'est une hypothèse
franchement irréelle et que l'on perd du temps
à l'étudier. |
Cet exemple vous permettra d'apprécier
combien les modèles sont variés et si certains
s'imposent actuellement dans la communauté scientifique ils ne
sont pas plus prouvés ou définitifs que d'autres. Ce
n'est pas le consensus qui fait la vérité. En histoire,
la vérité peut même être
définitivement (?) cachée, inaccessible. Je n'ai pas de
vérité à proposer par contre je pense pouvoir
aider un élève à réfléchir...
Il y a tout de même un aspect sur l'évolution qui est
essentiel mais qui ne tranche pas forcément avec les questions
que se pose tout historien. A quoi sert d'étudier l'histoire ?
A quoi sert d'étudier l'histoire de la vie ? Pour le
biogéologue la réponse est claire ce me semble : elle
va lui permettre d'élaborer un modèle incluant
le temps qui sera prédictif. En fait l'histoire est un
moyen d'atteindre l'avenir. Le modèle prédictif
élaboré devient testable, vérifiable et donc
scientifique. Il y a une sorte de contradiction : on élabore
un modèle historique qui est testé scientifiquement.
(J'ai déjà abordé cette question de
modèle d'une part dans les pages sur la
méthode et d'autre part dans une
petite page complémentaire sur les
modèles scientifiques
expérimentaux, mais le débat est loin d'être
clos).
* L'actualisme est un principe premier pour le paléontologue .
|
|
|
(actuelles, vérifiables, donc toujours soumises à l'expérience...) étaient aussi valables par le passé. |
Que considère t-on ? la durée des
phénomènes, la fréquence, la vitesse ? ou la
validité des lois, ou la réalité du passé
qui n'est plus mais qui a été ? Selon les formulations
il y a de profondes différences. Je n'ai pas fait de
recherches historiques approfondies mais il est tout à fait
possible que la formulation de ce principe ait beaucoup
évolué et que la définition que j'en donne ne
soit pas acceptée par tous (si je ne me trompe c'est d'abord
le catastrophisme de Cuvier qui a dominé
(révolutions subites) et a été
remplacé par l'uniformitarisme de Lyell et de Lamarck
(théorie du développement continu) pour finalement
déboucher sur cette présentation moderne d'un principe
philosophique plus ancien qui dépasse de loin la querelle
catastrophisme-uniformitarisme ; voir par exemple : Histoire de la
création des êtres organisés d'après les
lois naturelles, Ernest Haeckel, troisième leçon (p
52-53) et quatrième leçon, C. Reinwald et Cie,
libraires-éditeurs à Paris, 1874 ). La formulation
ci-dessus me convient et me paraît incontournable.
Le principe des causes actuelles nous permet de dire que les causes
de l'évolution sont actuellement décelables car
actuellement en jeu. Je précise : les causes et non pas
forcément les mécanismes, dans le sens où
il peut très bien y avoir eu des étapes dans
l'évolution qui sont passés et ne se reproduiront plus.
Ce sur quoi il est nécessaire de s'accorder c'est sur la
causalité et donc sur la validité des lois
expérimentales par le passé. Les lois
expérimentales découvertes actuellement s'appliquaient
par le passé.
En énonçant ce principe le scientifique sait qu'il fait
reposer sa connaissance sur un principe raisonnable mais qui n'est
pas scientifique dans le sens où il n'est pas
démontrable expérimentalement. Toute discussion de la
validité de ce principe reste dans le cadre d'une discussion
scientifique mais se fait non pas avec les outils du scientifique
mais avec ceux du philosophe.
L'uniformitarisme est une version très
différente qui stipule que les mécanismes actuels sont
les mêmes que par le passé. Il ne s'agit plus des causes
mais des phénomènes eux-mêmes, de leur
vitesse...on y oppose le catastrophisme qui lui
préfère des mécanismes différents.
Aucun de ces deux principes n'est utile pour le biogéologue
(je veux dire qu'aucun n'est indispensable, et qu'on peut donc les
rejeter comme principes), il s'agit de points de vue dont la
querelle est à mon avis dépassée actuellement
(notamment présenter les discussions sur la crise K-T à
l'aide de ces deux concepts relève à mon avis d'une
erreur... : que l'on préfère une vision où
l'autre ne change rien au fait que les deux partis appliquent le
principe des causes actuelles qui lui est incontournable).
* Il n'y a pas de temps
réel en dehors de la vie.
Du temps en biologie au temps en géologie : une
petite vision bergsonienne du
temps dans une page sur le
temps sur le site associé.
Le temps du géologue a longtemps été
marqué par le non vivant (pour ne pas dire l'inanimé).
Il serait donc judicieux non pas de séparer les
méthodes de datation (détermination de l'âge d'un
objet géologique) , comme je l'ai fait pour la
spécialité et comme on le fait habituellement, en
datation expérimentale et en datation logique,
mais au contraire de distinguer d'une part "la méthode qui
tient compte de la vie": la méthode
paléontologique, et d'autre part une méthode
physique, qui ne tient pas compte de la vie (essentiellement la
datation expérimentale isotopique).
Je vous renvoie au chapitre de géologie
de spécialité où sont
développés quelques éléments sur la
mesure du temps en géologie. En quelques mots, on peut retenir
que la datation expérimentale permet d'atteindre un
âge qualifié à tort d'âge absolu
alors qu'il doit être donné avec une incertitude
(c'est donc plus exactement un âge
expérimental, donné avec une incertitude
expérimentale absolue). Alors que la datation logique
repose sur un certain nombre de principes exposés
dans le cours de spécialité et permet d'atteindre un
âge paléontologique ou encore historique et logique.
Si l'on souhaite continuer dans le sens d'une écologie
historique il faut donc essayer d'intégrer les temps multiples
aux modèles pour obtenir une description de l'évolution
qui serait le mouvement du système terre-vie
(écosphère ou biosphère au sens large).
Remarque à propos de l'enregistrement
sédimentaire (ce que l'on nomme les "archives"
paléontologiques) :
Il me semble que la vision "sédimentaire" de l'enregistrement
paléontologique c'est-à-dire que chaque couche
fossilifère (ou roche) se superpose dans le temps aux
précédentes (principes de superposition et
d'identité paléontologique, voir cours de
spécialité),
n'est pas tout à fait exacte (si j'en crois mes sources ce
principe de superposition nous vient de Cuvier et était
lié aux révolutions subites prônées
par son auteur (Histoire de la création des êtres
organisés d'après les lois naturelles, Ernest Haeckel,
troisième leçon, p 53, C. Reinwald et Cie,
libraires-éditeurs à Paris, 1874 ) puis il a
été modifié par Lamarck et Lyell notamment
montrant que l'on peut trouver dans deux couches sédimentaires
superposées les mêmes fossiles : ils voulaient insister
sur l'aspect continu et progressif du développement du vivant
dans le temps ; mais mon propos est autre : c'est l'âge du
fossile, c'est la notion même de fossile appartenant à
une couche sédimentaire que je souhaite remettre en cause).
Dans cette vision de couches sédimentaires fossilifères
d'âge donné, il suffit de retrouver une couche ou une
roche ancienne pour avoir accès aux êtres vivants qui
vivaient à l'époque dans le sédiment ayant
donné naissance à cette roche. On se rend bien compte
qu'il peut y avoir des modifications à la suite de
l'enfouissement, des phénomènes tectoniques, des
remaniements géochimiques, une action du métamorphisme,
mais globalement on ne remet pas en cause cette superposition
horizontale de couches (roches) de plus en plus récentes vers
le haut. On a une vision fixiste de la sédimentation
qui éventuellement est modifiée
ultérieurement.
Pourrait-on avoir une vision dynamique ?
Si les interactions entre la terre milieu de vie et les
écosystèmes qui la peuplent ont été aussi
importantes par le passé que actuellement, dans ce cas la vie
modifie toutes les couches, y compris les roches en formation. Il n'y
a pas de milieu sédimentaire azoïque (le sédiment
est un milieu de vie): on retrouve des bactéries jusque dans
des roches profondes jusqu'à 2,8 km (voir par exemple
l'article : les micro-organismes de l'intérieur du globe,
James Fredrickson et Tullis Onstott, Pour la Science, 230,
décembre 1996, 90-95 , qui citait déjà 9.000
souches bactériennes lithotrophes profondes...). Les limites
physico-chimiques des conditions de vie semblent sans cesse
s'élargir et il me semble que l'on a quasiment aucun milieu
sur terre (dans cette mince - à l'échelle de la
planète - pellicule que l'on appelle biosphère)
où l'on a pu mettre en évidence des êtres
vivants. Dans ce cas, les remaniements ne sont pas l'exception mais
la règle, dans ce cas les éléments chimiques
d'une roche n'ont pas un âge unique mais il y a une grande
hétérogénéité au sein de chaque
roche. Le fossile est donc l'exception et n'est vraiment jamais
intact. Dans ce cas comment arriver à chiffrer dans une roche
sédimentaire la part des chacune des périodes qui
s'interpénètrent ? Comment même dater un fossile
? On pourrait donc substituer à la vision sédimentaire
une vision plus "écologiste" dont voici une illustration
:
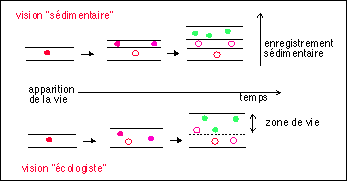 Deux visions de l'enregistrement sédimentaire au cours des temps géologiques : (les points colorés pleins indiquent des populations d'êtres vivants, les points colorés vides des populations mortes) |
dans la vision sédimentaire "fixiste", le temps qui sépare deux couches correspondant à des milieux de vie différents est considéré comme si important que l'on peut voir deux milieux superposés (un actuel, un passé) indépendants ou du moins qui peuvent être reconstitués de façon indépendante; |
|
dans la vision écologiste "dynamique" il n'y a jamais séparation réelle, les roches sédimentaires profondes ne sont pas à l'abri des modifications liées au vivant : la zone azoïque commence à plus de 3 kilomètres de profondeur en milieu continental et certainement bien plus (10 km ?) en milieu océanique. |
|
|
N.B. |
|
Si un homme croît pouvoir
créer un être vivant à partir de molécules
et d'énergie, il se prend pour Dieu
1.Une conviction qui, à mon sens, devrait être celle de
tous les scientifiques expérimentaux : l'étude des
conditions de l'apparition de la vie n'est pas du domaine de la
recherche expérimentale.
Un exposé particulièrement clair est celui de Pierre
LASZLO dans La Recherche (n°296, mars 1997, 26-28:
Origine de la vie: 100.000 milliards de scénarios): il
y affirme que la chimie prébiotique, ou évolution
moléculaire pour les darwiniens, est une science historique.
Tout scénario sur l'apparition de la vie reste une histoire et
non l'Histoire (voir dans ce même article la citation de
l'historien Paul Veyne). La certitude, l'adhésion de l'esprit
ne vient pas de la méthode expérimentale mais d'une
autre méthode (celle de l'histoire), même si elle est
aussi raisonnable.
Remarque:
Il est toujours aussi surprenant, pour un enseignant, de voir comment
on (?) a pu imprimer dans l'esprit des élèves
l'idée selon laquelle le temps, allongé à
l'infini (ou presque), permet de diluer la causalité.
Peut-être y a-t-il un rapprochement à faire avec le
paradoxe de la flèche de Xénon qui n'atteint jamais son
but étant donné que l'espace est infiniement divisible.
Cette division de l'espace, comme celle du temps abstrait, est en
puissance et non en acte, comme nous l'expliquent les
métaphysiciens. La causalité doit atteindre son objet
tout comme chaque action a une cause directe; repousser dans le temps
la cause ne tient que pour le temps abstrait et non pour le temps
concret qui fait que chaque être vivant, à chaque
instant de vie, doit être soutenu dans son être,
puisqu'il n'est pas sa propre cause.
Toute connaissance biologique est
aujourd'hui étudiée dans une perspective
évolutive
2. Un point de vue intéressant (qui peut paraître en
contradiction avec le premier point mais qui en fait le rejoint
partiellement car, si l'apparition de la vie échappe à
l'expérience, notre compréhension du vivant passe par
cette expérience et l'évolution fait partie de cette
théorie globale du vivant qui nous passionne tous):
«ce ne sont pas les biologistes moléculaires qui font
actuellement avancer les recherches sur l'origine de la vie, ce sont
des physiciens et des chimistes (et aussi des biologistes... mais
qui utilisent les outils mathématiques) qui
s'intéressent aux formes... ». Je voudrais citer Yves
Bouligand : «Qui peut aujourd'hui présenter un
séminaire sur les facteurs contrôlant la forme et la
nature du Golgi, avec une analyse des aspects évolutifs de la
question ? Le mieux que l'on puisse faire est de confronter
l'ensemble des données disponibles et de faire le tour des
problèmes, mais il faudrait tout ignorer des facteurs
physico-chimiques qui contrôlent la forme des bicouches
membranaires pour penser que tout cela n'est qu'une question de
gènes que l'on va découvrir en temps utile.»
J'en profite pour recopier un passage qui
suit : «Nombre de biologistes en France sont restés
durant de longues années les fervents défenseurs de
l'hérédité de l'acquis, et sont remplacés
aujourd'hui par de piètres avocats du darwinisme, biologistes
moléculaires en général, en des domaines qui ne
constituent certainement pas la meilleure sélection de cette
discipline, dont certains spécialistes affichent un dogmatisme
contraire à l'esprit du darwinisme. La raison de cette
état de choses tient selon nous à un manque grave
d'interdisciplinarité dans les programmes de formation
scientifique.» Yves Bouligand, La
petite fronde anti-Darwin des années récentes, in
"Pour Darwin", 1997, PUF, 751-783
Les fossiles incontestables
commencent vers 600 Ma.
3. On peut essayer de préciser POUR CES TEMPS TRÈS
ANCIENS, le sens de la recherche de fossiles (restes ou traces
d'êtres vivants) ou de roches les plus anciennes possibles
:
* La
forme des microfossiles reste le premier
critère de détermination
Or, si l'on trouve des formes, il y a toujours l'incertitude
historique, même si le chercheur peut avoir une intime
conviction : les filaments, microcapsules, microsphères,
bâtonnets en sont des exemples... (voir par exemple les photos
de lame mince de la "Gunflint Iron Formation" datée de 2Ga
dans La Recherche, 295, février 1997, p31 ainsi que la
brève journalistique "Controverse : les premières
traces de vie", Fabienne Lemarchand, La Recherche, 354,
juin 2002, p 14-15, qui rapporte la réinterprétation
(par l'équipe de Martin Brasier de l'université
d'Oxford) des traces autrefois qualifiées de
bactériennes datées de 3,465 Ga (Chinaman Creek, en
Australie) comme des productions minérales associées
à une activité hydrothermale: les techniques d'analyse
utilisées ici ont été particulièrement
complètes notamment par l'utilisation de la
spectrométrie Raman laser... mais les résultats restent
bien sûr controversés... nous ne
sommes pas dans une science expérimentale).
Par contre dès que l'on arrive à des squelettes
élaborés l'esprit se rallie avec aisance, l'incertitude
historique demeure mais la raison est bien convaincue que l'on a des
restes d'êtres vivants : des fossiles incontestables.
(Mais l'on a toujours pas de preuve expérimentale ... que l'on
aura jamais)
Remarques:
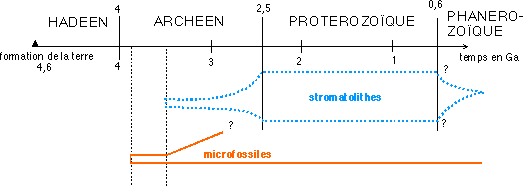
Un exemple d'utilisation de cette prudence dans le domaine historique est celui de l'étude de ce qu'on appelle les stromatolithes...ce terme, et ce qu'il recouvre, est loin de faire l'unanimité de la communauté scientifique... : voici quelques éléments de discussion :
|
(une référence : Les cyanobactéries, d'étonnants procaryotes autotrophes, Marc-André Selosse, Biologie-Géologie (Bulletin de l'APBG), n°3-1996, 481-529) |
|
Ne prétendant pas du tout être un spécialiste de cette question, je me contenterai de résumer ce que j'ai compris de la référence citée, puis je soulèverai quelques questions... Ce sont des organismes VIVANTS Les questions: |
* La
composition chimique des microfossiles n'est
déterminée qu'avec des incertitudes
expérimentales très grandes et de nombreuses
incertitudes historiques absolues;
En effet, si l'on s'intéresse à la composition
chimique, malgré les apparences, on est encore plus dans
le domaine de l'incertitude historique, même si la conviction
d'un chimiste, habitué à manier les isotopes (du
grec iso = identique et topos = lieu ; les isotopes sont des
éléments chimiques qui ont le même numéro
atomique (et donc le même nom et la même position dans le
tableau de Mendeleiev) mais qui sont de masse atomique
différente), peut être forte et contagieuse.
En ce qui concerne la minéralogie (géochimie non
isotopique) on peut par exemple utiliser des diagrammes Eh/pH qui
indiquent des conditions de précipitations dans les
conditions expérimentales actuelles : on en déduit
un environnement géochimique possible, aux incertitudes
près et toujours dans l'hypothèse actualiste (on
remarque que lorsque l'on se tourne vers des temps très
anciens, l'hypothèse actualiste est souvent confondue avec des
conditions physico-chimiques actuelles ce qui est tout à fait
différent du principe actualiste (voir plus haut
catastrophisme-uniformitarisme qui font référence
à des mécanismes): dire que les lois
expérimentales sont extrapolables au passé ne
présume pas des conditions physico-chimiques qui
étaient peut-être très différentes de
l'actuel et qui n'ont peut-être pas fini de nous
surprendre...).
En ce qui concerne la géochimie isotopique, on peut
utiliser des isotopes stables ou des isotopes
radioactifs.
Les premiers permettent à l'aide du raisonnement physique
simple du partitionnement lors des processus du vivant de
montrer une signature isotopique du vivant. En effet de nombreux
processus du vivant (précipitation de minéraux formant
des squelettes, des coquilles, photosynthèse...) partitionnent
(on dit aussi discriminent, c'est-à-dire séparent) les
isotopes.
Enfin les isotopes radioactifs permettent des datations
expérimentales (qui doivent être
présentées avec des incertitudes (et avec toujours une
incertitude absolue : la réouverture du système) : je
vous renvoie au cours de spécialité).
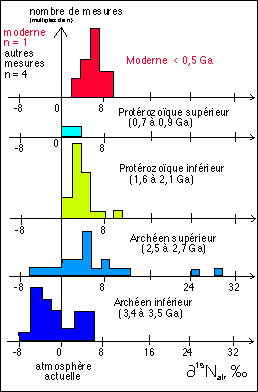
Voici un exemple d'utilisation des isotopes stables de l'azote
(j'ai du essayer de faire une analyse à partir de l'article
de Valérie Beaumont dans le dossier de Géochronique, je
manque bien sûr de sources de première main mais je
pense que l'on peut essayer de poser de bonnes questions...):
|
|
||||||||
|
|
Le diazote atmosphérique actuel est considéré comme le standard valeur 0 du delta 15NAIR en ‰ Le delta isotopique est la différence, pour un élément donné entre le rapport isotopique d'un échantillon (Re) et celui d'un standard ( Rs), divisé par le rapport isotopique du standard et exprimé en pour mille : delta = (Re-Rs)/Rs x 1000 où R, le rapport isotopique est toujours présenté avec l'isotope de masse la plus élevée au numérateur ; soir R pour l'azote = 15N/14N. |
|||||||
|
Dans les processus naturels ACTUELS les partitions
isotopiques de l'azote connues sont les suivantes : |
||||||||
|
Dans les organismes vivants actuels le delta 15N est voisin de 6‰. Comme la plupart du organismes autotrophes vis-à-vis de l'azote consomment des nitrates dissous dans l'eau (NO3-), on explique cette valeur par l'hypothèse selon laquelle l'azote lourd s'accumule dans les nitrates du fait de la dénitrification par les bactéries du sol. On retrouve donc cette valeur dans la matière organique des organismes hétérotrophes vis-à-vis de l'azote et qui consomment l'azote des protéines. Certains organismes autotrophes sont capables de FBA notamment à l'aide de bactéries symbiotiques pour des plantes de la famille des légumineuses (Fabacées) ou des Cyanophycées (vois encadré ci-dessous). Si ce diazote atmosphérique constitue leur source principale d'azote, il devraient donc présenter un abaissement de leur delta 15N. Mais Valérie Beaumont ne cite aucune mesure sur ces organismes actuels... existent-elles ? |
||||||||
|
On peut donc s'attendre à ce que, lorsque l'on
mesure le rapport entre les compositions isotopiques en
azote 14 et 15 de la matière organique d'un
échantillon, : si l'on trouve un rapport voisin de 0, on a |
||||||||
|
et Analyse |
|
|||||||
|
Conclusion |
L'utilisation des isotopes radioactifs est une méthode chère, avec une très grande incertitude (indépendamment de l'incertitude absolue liée à l'actualisme) qui permet d'aboutir, si l'on reste prudent, à des arguments intéressants mais certainement pas définitifs. La plupart du temps la prudence manque dans les conclusions. |
|||||||
Deux remarques personnelles :
* Sur quelles méthodes et arguments
reposent toutes les théories actuelles de la formation de
l'univers et de la terre ?
Réponse: sur des mesures d'abondance isotopique
(directe: roches actuelles et météorites; et
indirectes: par spectroscopie astronomique) puis des calculs
physiques intégrés dans des modèles
décrivant la matière et les forces entre ses
composants...
Donc: l'incompétence d'un enseignant naturaliste est
patente, et elle ne peut pas être comblée par une
formation continue, il s'agit vraiment d'un tout autre champ
disciplinaire. Cela n'empêche pas certains collègues de
s'intéresser à ces questions mais à mon avis
cette partie n'a rien à faire dans un programme de SVT de
terminale.
* Existe-t-il un milieu actuel directement
accessible à l'expérience sans trace de vie
?
Réponse: non.
(j'écarte volontairement le cosmos qui est peut-être
vierge de toute vie si on l'observe par un télescope mais qui
ne l'est probablement plus, malgré toutes les
précautions prises, une fois qu'il a été
observé directement à l'occasion d'un voyage
habité ou non ...).
Donc: la vie semble tout "contaminer", d'où
l'idée que le milieu n'existe pas, n'a jamais existé,
sans êtres vivants.
Conclusion:
Si l'on revient à la conception de la terre comme UN
écosystème, qui a commencé dans le temps, s'est
développé et continue actuellement de se
développer ; quelle certitude avons-nous de pouvoir atteindre
les étapes de la naissance de cet écosystème ?
Les premières étapes de la vie d'un être vivant
peuvent-elles êtres retrouvées à partir d'un
adulte ? Il n'est pas certain que l'histoire de la vie, qui est du
passé, soit accessible.
Les plus anciennes roches terrestres sont datées par les
méthodes isotopiques (qui utilisent la loi de
désintégration radioactive au cours du temps
d'éléments présents en très faible
quantité dans toutes les roches) d'environ 4.000 millions
d'années (on dit maintenant 4 Ga (giga années) ou 4
milliards d'années). On les trouve au Groënland
(formation d'Isua) et au Canada (Acasta) et elles ont un aspect
étonnamment actuel.
L'âge de la formation de la terre, comme du reste du
système solaire est estimé à 4.600 millions
d'années; cet âge est estimé à partir de
la datation de nombreuses météorites qui se seraient
formées en même temps que tous les autres
éléments du système solaire. La période
séparant la formation de la terre et l'apparition des
premières roches connues est appelée l'Hadéen
(de Hadès, le dieu des enfers). La période suivante de
-4 à -2,5 Ga étant dénommée
Archéen (du grec "archos", ancien). Le
Protérozoïque, de -2,5 Ga à -0,550 Ga,
précède ensuite le Paléozoique ou ère
primaire.
Des molécules organiques ont été trouvées
dans toutes les roches les plus anciennes comme dans de nombreuses
météorites et au sein même du système
solaire dans des objets comme les comètes ce qui conduit
certains à proposer des origines variées aux
molécules du vivant. Des chimistes étudiant ce qu'ils
appellent la "chimie prébiotique", tentent de
synthétiser des composés organiques spécifiques
du vivant, comme les acides nucléiques, sans l'aide des
systèmes biologiques.
Il existe quelques cas de structures observées en lames minces
et en microscopie optique et électronique qui peuvent rappeler
des colonies bactériennes. Certaines structures isolées
de roches sédimentaires très fines de Warrauwoona en
Australie, datant d'environ 3,5 milliards d'années, sont ainsi
interprétées comme des microfossiles. Des structures
concentriques appelées stromatolithes (ou stromatolites) dont
certaines remontent peut-être à 3,5 milliards
d'années comme celles de North Pole en Australie, sont aussi
interprétées par certains comme des concrétions
bactériennes (dues à des Cyanobactéries,
anciennement classées dans les Algues bleues, que l'on peu
également appeler Cyanophytes). Ces interprétations
sont fort discutées.
Plus hypothétiques encore les interprétations
concernant les énormes quantités de fer oxydé
(dépôts de fer rubané Fe2O3 associé
à des sulfates SO42-) datés de -2,3 à -1,7 Ga,
et qui pourraient provenir de l'oxydation du fer ferreux (Fe2+)
présent dans les océans à la suite du lessivage
des hypothétiques continents. Cette oxydation serait due
à la libération d'oxygène par des processus
biologiques comme par exemple la photosynthèse qui est
réalisée notamment par les Cyanobactéries
actuelles. Le fer aurait alors piégé l'oxygène
qui n'aurait été libéré dans
l'atmosphère que vers 1,7 Ga, une fois que tout le fer des
océans eut été oxydé en fer ferrique
(Fe3+). C'est à partir de cette époque que la teneur en
dioxygène de l'atmosphère terrestre aurait
augmenté pour atteindre les 21% actuels.
L'atmosphère primitive, évaluée à partir
des réservoirs de volatils actuels estimés (H2O, CO2,
CaCO3 et C) ou à partir des inclusions fluides de basaltes
profonds ou de météorites, aurait été
composée d'environ 80% d'eau et de 10% de CO2, le reste
étant essentiellement de l'azote. La couche d'ozone (O3)
protectrice vis-à-vis des u.v. solaires serait apparue vers
-0,4 Ga puisque c'est vers cette époque que semblent
apparaître les premiers organismes aériens.
Les découvertes d'écosystèmes très
anciens ne sont pas nombreuses. On peut citer la faune d'Ediacara
(découverte dans le Sud Australien vers 1940 ?) datée
de 580 à 560 millions d'années renfermant des formes
segmentées de taille variable (de quelques centimètres
à près d'un mètre) difficiles à
interpréter et les schistes de Burgess (découverts
à l'Est du Canada dès 1909), datés de 530
millions d'années et renfermant d'incontestables Trilobites.
D'autres découvertes de moindre ampleur ont été
faites depuis dans des roches cambriennes. Du fait de la
rareté des roches très anciennes et de
l'intensité du métamorphisme qu'elles ont subies, il
n'y a rien d'étonnant à ce que la découverte de
fossiles aussi anciens soit exceptionnelle. (Il ne semble pas que
l'on puisse affirmer qu'il existait au début du Cambrien une
faune riche et diversifiée et encore moins que les mers
grouillaient de formes animales primitives...je n'ai fait de
recherches approfondies sur ces découvertes fossiles mais
elles ne doivent pas atteindre la dizaine de sites au total !!! quand
au nombre de formes "originales", elles ne dépassent pas
quelques dizaines; les descriptions de J.J. Sepkoski dans "Le livre
de la vie" (Stephen Jay Gould, Seuil, 1993) sont peut-être un
peu trop imaginaires...).
Dès la fin du Cambrien les fossiles sont de plus en plus
nombreux à l'instar des affleurements datés de cette
période.
La formation de la terre et du système solaire est datée de 4,5 Ga (milliards d'années). La vie serait apparue dès 3,5 Ga avec des formes bactériennes réalisant des réactions chimiques en tout point semblables à celles réalisées par les organismes actuels. Notamment, on pense que la photosynthèse, utilisation de l'énergie lumineuse solaire pour réaliser des synthèses organiques, serait apparue d'emblée dans l'histoire de la vie. Cette photosynthèse s'accompagnant de dégagement de dioxygène, celui-ci est progressivement dégagé dans l'atmosphère. Vers -400 millions d'années la quantité de dioxygène devient suffisante pour former une couche d'ozone (O3) protectrice des rayons u.v. solaires et permettre ainsi l'apparition de formes vivantes aériennes. Jusqu'alors la vie était en effet cantonnée au milieu aquatique.
La vie est apparue il y a très longtemps, et elle s'est développée sur la terre.
Notez la difficulté à changer ou à ajouter des termes : l'apparition de la vie ne peut être datée que par des méthodes expérimentales (avec une grande incertitude expérimentale et une incertitude expérimentale absolue : historique - liée à l'actualisme). Les termes "très longtemps" sont les plus neutres possible. Si l'on ajoute "sur terre", on va à l'encontre de nombreuses hypothèses d'apparition de la vie dans l'espace qui sont loin d'être farfelues. Le fait que la vie se soit développée, c'est-à-dire qu'il y ait eu des changements, semble être aussi difficilement contestable. Ce développement est à mon sens, le "véritable" évolutionnisme (véritable au sens originel, je voudrais dire scientifique, mais en fait il s'agit encore une fois d'histoire, même si le consensus est quasi général : c'est ce que l'on appelait auparavant le transformisme). Mais les modalités d'apparition des différents groupes restent encore très hypothétiques et il ne me paraît pas utile de privilégier une hypothèse plutôt qu'une autre.
|
étymologiquement fossile signifie
"tiré de la terre" (du latin fodio, is, fodi,
fossum = creuser, fouir) et on trouve principalement
deux types de définitions: |
|
La fossilisation désigne l'ensemble des
phénomènes qui conduisent à la
formation d'un fossile ou plus précisément
à la conservation des êtres vivants ou de leurs
traces dans les sédiments puis dans les roches
sédimentaires. Un article: "La fossilisation : une exception conjoncturelle", Christiane Denys, Pour La Science, 292, février 2002 |
Etant donné qu'un historique de
l'apparition du concept de fossile (la création du mot dans le
sens actuel est rapporté à Lamarck
in La naissance du transformisme, Lamarck,
entre Linné et Darwin, Goulven LAURENT, Collection
inflexions, Vuibert/Adapt, 2001, p 15 ) est hors de ma
portée, je souhaite juste faire une citation: «En
même temps qu'il créait la zoologie des
Invertébrés, Lamarck en fondait en effet la
paléontologie» (op. cit., p
15) et rapporter une anecdote, racontée
elle-même par Georges Cuvier (1769-1832) dont le travail a sans
doute été déterminant dans la naissance de la
paléontologie (des Vertébrés) mais aussi de
l'anatomie comparée. «Dès son premier
mémoire sur les Éléphants fossiles, Cuvier
émet l'idée d'une création d'animaux
antérieurs à la création actuelle,
création entièrement détruite et perdue. Cette
hypothèse servira de point de départ à de
brillantes recherches qui se poursuivront pendant trente
années, malgré les plus grandes difficultés.
Dans le cas des Mammifères, auxquels s'intéressait
particulièrement Cuvier, il est infiniment rare de trouver un
squelette fossile à peu près complet: «Des os
isolés et jetés pêle-mêle, presque toujours
brisés et réduits à des fragments, voilà
tout ce que nos couches nous fournissent, et la seule ressource des
naturalistes.» Il fallait donc être capable de
déterminer ou de reconstituer, à partir d'un fragment
d'os, l'animal auquel il avait appartenu, art presque inconnu au
moment où Cuvier commença ses recherches. L'anatomie
comparée lui fournissait le principe nécessaire
à cette détermination: le principe de
corrélation des organes, selon lequel chaque partie d'un
animal peut être donnée par chaque autre, et toutes par
une seule. De la forme des dents, par exemple, on pourra conclure la
forme des pieds, celle des mâchoires, celle des intestins.
Cette déduction rigoureuse, sinon infaillible, a souvent
permis à Cuvier de reconnaître un animal à partir
d'un fragment d'os ou d'une dent. On connaît l'anecdote qu'il a
lui-même rapportée à propos de la
découverte d'un Didelphe dans le gypse de Montmartre;
l'examen des dents lui ayant montré la
parfaite analogie de ce fossile avec les Sarigues, il ne doute point,
avant d'avoir vu le bassin, que celui-ci portait des os marsupiaux.
En présence de quelques amis, il fit creuser la pierre et mit
au jour le bassin; les os marsupiaux s'y pouvaient
voir.»
Comme l'affirme Jean Piveteau : «jusqu'à Cuvier,
l'anatomie comparée n'était qu'un recueil de faits
particuliers concernant la structure des animaux. Cuvier en fit la
science des lois de l'organisation animale»
(Jean Piveteau in E.U. article "Georges
Cuvier").
Elle s'efforce , à partir d'un certain nombre de principes, de déduire des caractéristiques invisibles (anatomiques, physiologiques, comportementales...) de l'analyse (étude morphologique, géochimique (y compris isotopique)...) et de la comparaison des fossiles entre eux et de la comparaison des fossiles avec les formes vivantes actuelles (pour ces comparaison les statistiques sont un outil de choix).
Deux principes de la paléontologie (à
compléter avec les principes de la (bio)stratigraphie:
voir "datation logique" dans le cours
de spécialité):
* principe de corrélation entre organes (principe
morphologique et anatomique): chaque partie (organe) d'un organisme
(organum, voir l'introduction
du cours de seconde pour une petite
recherche étymologique) est ordonnée au tout, si
bien que par la connaissance d'une seule des parties et par
comparaison avec un organisme présentant une partie semblable,
on peut en déduire la forme de parties disparues.
* principe d'uniformitarisme ou principe de
corrélation environnementale: "en absence d'indices
contraires, on suppose que les conditions de vie des organismes
disparus ayant donné les fossiles, étaient semblables
à celles des organismes actuels les plus voisins"; c'est ainsi
que l'on détermine le faciès paléontologique (le
faciès d'une roche sédimentaire est
l'ensemble des ses caractères lithologiques (de la roche) et
paléontologiques (de ses fossiles)) qui nous permet de faire
des reconstitutions paléogéographiques. Une coquille
d'une "huître" datée de 70 millions d'années est
l'indice d'un milieu de balancement des marées, étant
donné que l'on suppose que les huîtres ayant
donné ces coquilles fossiles vivaient dans le même
milieu de vie que les huîtres actuelles. Il existe des cas
très précis pour lesquels on a pu prouver que ce
principe n'était pas applicable, par exemple pour les
Crinoïdes (lys de mer) qui ont migré du plateau
continental au talus au cours des temps géologiques.
* le principe d'uniformitarisme appliqué à la
physiologie ou principe de corrélation physiologique,
permet de la même façon de déduire des
composantes physiologiques d'un organisme disparu à partir de
celles d'un organisme actuel le plus voisin.
Les déductions réalisées par le paléontologue au moyen de cette logique sont habituellement appellées des faits paléontologiques. Cette apellation est légitime si l'on ne confond pas cette notion avec un fait expérimental.
Une fois les faits paléontologiques dégagés
de l'étude des fossiles, le paléontologue s'attache
à reconstituer l'histoire des êtres vivants d'une
façon plus globale et relie donc les faits entre eux afin de
dégager une théorie , à moins que cette
théorie ne préexiste à l'interprétation
des faits.
La principale théorie scientifique est le
transformisme (= les
espèces dérivent les unes des autres), ce que
maintenant tout le monde nomme
l'évolution (= les
espèces ont évolué au cours du temps); les
espèces actuelles sont le résultat de
l'évolution des espèces anciennes disparues qui nous
sont connues par les fossiles. (Nous
étudierons plus en détail cette question dans
un chapitre
suivant)
Comme pour les modèles en science expérimentale, il
me semble que l'y ait deux positions possibles :
* soit on essaye d'établir des parentés directes
à partir de critères dégagés à
partir des fossiles (modèle externe) ; on est dans ce
cas toujours limité par le type de renseignements,
forcément non vérifiable expérimentalement, que
fournit le fossile. Dans le cas de ce type de modèle il faut
partir du fossile et essayer d'en déduire le maximum de
renseignements.Ce sont les fossiles qui permettent
d'établir le modèle.(pour prendre une comparaison
c'est un peu comme si l'on cherchait à partir des
données historiques individuelles de chaque homme, de chaque
catégorie professionnelle, pendant une dizaine
d'années, à comprendre comment ils ont pu
réaliser une révolution)
* soit on essaye de trouver une théorie unificatrice de
l'évolution (modèle interne) et dans ce cas,
étant donné que les forces présidant à
l'évolution peuvent être considérées comme
toujours actuelles, on peut très bien utiliser les
données concernant les êtres vivants actuels tout comme
les données des fossiles. Dans ce type de modèle, on
part d'une théorie que l'on pose comme un postulat
scientifique (dont la validité peut être discutée
séparément), et on essaye de trouver tous les arguments
possibles sur les êtres vivants et fossiles qui s'accordent
avec le modèle: on intègre les fossiles au
modèle. (pour reprendre la comparaison de la
révolution, le modèle interne suppose un modèle
de révolution, social, économique, conceptuel, et on
cherche les données historiques qui s'y accordent). Je
souligne bien que dans ce dernier cas l'honnêteté
intellectuelle de l'historien n'est pas mise en cause : si celui-ci
trouve une donnée historique qui ne s'accorde pas à son
modèle il est tout à fait capable de modifier le
modèle.
Dans les deux cas, le principe de l'actualisme reste bien
sûr valable et délimite le caractère historique
de ces modèles.
Le plus étonnant est qu'un modèle peut ensuite servir
à prédire. Dans le cas d'un modèle
externe, la prédiction peut être par exemple de supposer
l'existence de formes fossiles intermédiaires. En cas de
découvertes en accord avec le modèle, on conforte
celui-ci mais on ne le valide pas expérimentalement pour
autant. Dans le cas d'un modèle interne, la prédiction
est peut-être plus difficile à établir. Etant
donné sa prétention à une explication globale de
l'évolution et donc de l'avenir du vivant, ce modèle
peut être testé dans le temps.
On peut essayer de donner un exemple : l'évolution des
Equoïdés.
documents extraits de TD, Nathan, 1983; Précis de géologie 2, paléontologie, stratigraphie, J. Aubouin, R. Brousse et J.P. Lehman, Dunod Université, 1978 ; modifiés:
|
puis récent (depuis 10.000 ans) |
|
|
(milieu du Pliocène (-2,5 Ma) à l'actuel) |
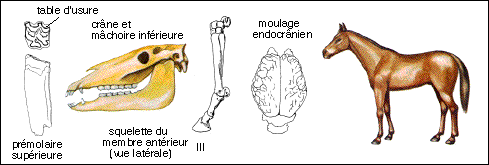 Les échelles ne sont respectées qu'entre les silhouettes reconstituées des équoïdés sur les 4 schémas... |
|
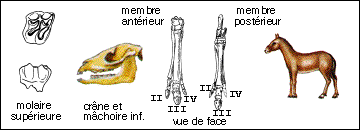
La famille actuelle des Equidés fait partie du sous-ordre des Hippomorphes et à l'ordre des Périssodactyles (Mammifères placentaires chez lesquels les membres présentent un troisième doigt prépondérant par lequel passe l'axe du membre selon lequel le poids du corps repose ; à ce groupe appartiennent aussi les Cératomorphes : Tapirs et Rhinocéros). Elle regroupe les genres Equus (espèce unique Equus caballus), Asinus (les ânes), Hemionus (Hémiones vrais et Onagres) et Hippotigris (Zèbres).La seule sous-espèce encore à l'état sauvage dans les steppes d'Asie est le cheval de Prjewalskii : Equus caballus prjewalskii. Il semble avoir été domestiqué pour la première fois en Asie à la fin du néolithique. Les races sont très nombreuses (du Poney des Shetlands au Percheron, en passant par le Cheval arabe). On trouve des Equus à l'état fossile depuis le Pliocène en Amérique du Nord puis en Europe au Quaternaire (Pleistocène : de -1,8 Ma à 10.000 ans). Mais il est étonnant de noter que lorsque les colons européens arrivèrent en Amérique au 18ème siècle, ils réintroduirent le cheval qui semblait y avoir complètement disparu (depuis la fin du Pléistocène). |
|
|
|
||
|
(Miocène sup et Pliocène) |
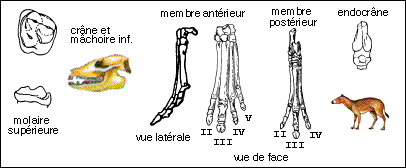
On classe dans ce groupe (Simpson) des Equidés très voisins du cheval mais dont les doigts latéraux sont plus développés que chez le cheval, dont les molaires sont convexes vers l'extérieur alors qu'elles sont droites chez le cheval. |
|
|
(Miocène) |
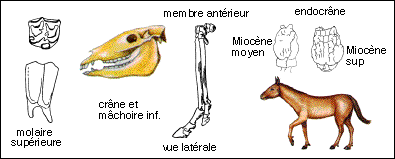 Les échelles ne sont respectées qu'entre les silhouettes reconstituées des équoïdés sur les 4 schémas... |
|
|
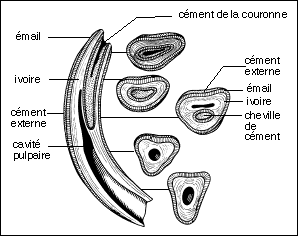
Merychippus atteignait probablement 1m à l'épaule. Le museau est plus allongé que chez les Equidés plus anciens, l'orbite se ferme postérieurement. Le cubitus et le radius sont fusionnés et le péroné se réduit à une large baguette osseuse. Les doigts latéraux sont courts, le poids du corps n'ayant probablement porté que sur le doigt médian. Les dents semblent sont du type hypsodontes (elles sont plus élevées que les dents brachydontes et elles s'usent par leur couronne) et semblent correspondre à un régime alimentaire herbivore (on peut essayer de relier ce type de dentition à l'extension des prairies à Graminées de la flore Miocène); à l'intérieur des crêtes d'émail du cément est visible pour la première fois. |
||
|
|
||
|
(Oligocène sup et Miocène) |
Les dents semblent se rapprocher plus d'un type nutrition herbivore ("mangeur d'herbe" par opposition à phyllophage : "mangeur de feuilles") du fait de l'hypsodontie (les dents hypsodontes sont plus élevées que les dents brachydontes et elles s'usent par leur couronne) mais le cément n'existe cependant pas à l'intérieur des crêtes d'émail chez Parahippus. Les hémisphères cérébraux recouvraient en grande partie les lobes olfactifs. |
|
|
(Oligocène moyen à supérieur) |
Les échelles ne sont respectées qu'entre les silhouettes reconstituées des équoïdés sur les 4 schémas... |
Formes "transitoires" qui ne se distinguent de Mesohippus que par des différences mineures notamment au niveau des os du tarse. |
|
(Oligocène) |
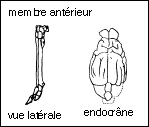
Découvert en Amérique dans le Dakota (Badlands) avec 14 excellents squelettes avec un aspect que l'on imagine très semblable à celui d'un petit cheval (60 cm de hauteur à l'épaule). La tête rappelle celle du cheval mais avec une mandibule plus grêle. Les prémolaires 2, 3 et 4 sont molarisées et montrent comme les molaires des crêtes d'émail nettes. L'alimentation probable est la même que celle de l'hyracotherium. Apparaît chez Mesohippus pour la première fois une cupule d'usure au niveau des incisives (ou cornet), utilisée pour vérifier l'âge des chevaux dans les marchés de bétail. Le doigt externe (VI) est présent au niveau du membre antérieur mais fortement grêle (vestige ?). Dans l'encéphale, des sillons caractéristiques de la surface cérébrale des Equidés actuels sont nombreux et les hémisphères cérébraux recouvraient partiellement le cervelet. |
 |
|
|
|
|
(Amérique du Nord) = Hyracotherium (Europe) |
Les échelles ne sont respectées qu'entre les silhouettes reconstituées des équoïdés sur les 4 schémas... |
|
Eohippus a été trouvé pour la première fois dans les Montagnes Rocheuses (Bassin de Wasatch) et daté d'environ 40 Ma (Eocène sup : Sparnacien, Bartonien). Il possède les mêmes caractères que l'Hyracotherium trouvé en Europe (dans la province de Kent par Owen en 1839) et antérieur (Sparnacien, environ -53 Ma) mais en bien moins grande abondance. On doit donc utiliser le nom d'Hyracotherium qui est le premier à avoir été donné. Il avait une hauteur à l'épaule estimée entre 25 et 50 cm (un gros lapin !). Il possède une denture complète de 44 dents mais une barre entre les incisives et les molaires apparaît : il s'y logeait une petite canine (cette dent n'existe que chez les mâles des chevaux actuels). Les deux prémolaires antérieures sont coupantes tandis que les deux prémolaires postérieures et les molaires sont broyeuses. Les molaires ont une couronne présentant 4 tubercules principaux et deux tubercules accessoires. Les dents broyeuses sont basses (type brachydonte) et la nourriture devait constituer de feuilles succulentes et de fruits car l'on pense que l'herbe siliceuse (graminées) aurait usé beaucoup trop vite des dents de ce type. A la patte antérieure seul le premier doigt manque alors que le cinquième fait aussi défaut à la patte postérieure. Métacarpe et métatarses sont assez longs. Le moulage endocrânien montre des lobes olfactifs très développés, un encéphale quasiment lisse, les hémisphères cérébraux ne recouvrant pas les tubercules quadrijumeaux. |
|
Voici un extrait de L'évolution du vivant de Pierre Paul
Grassé (p93-96) dans lequel il parle de la présentation
de l'évolution des équoïdés faite par G.
Simpson, notamment dans son livre "Horses" (1951) : «Le
récit que G. Simpson donne de l'histoire des
Equoïdés édifie sur l'influence qu'exerce la
doctrine sur l'observateur. Le darwinien inconditionnel nie
l'existence de l'évolution orientée, bien qu'elle soit
évidente, et la confond à tort avec
l'orthogenèse, malgré cela, il y fait un appel constant
et involontaire.
Il se range à l'opinion classique d'après laquelle tous
les Equoïdés dérivent des genres Hyracotherium
(Eocène d'Europe) et Eohippus (Eocène d'Amérique
du Nord), qu'il met en synonymie, et qui ont donné deux
lignées : les Equidés vrais en Amérique du Nord
(ils ont passé en Europe au
Pliocène-Pléistocène) et les
Paléothéridés en Europe. Ne considérons
que les Equidés qui ont buissonné en petits rameaux
évolutifs, tels ceux des Anchitherium, des Hypohippus, des
Stylohipparion... et en Equidés proprement dits, lesquels se
terminent par le genre Equus. Selon Simpson, la diversité des
lignées démontre l'inexistence de l'orientation
évolutive ; or, ce qu'il décrit est
précisément l'évolution buissonneuse ou
verticillée, dont la forme caballine est le leitmotiv dans
toutes les sous-lignées; elle y est plus ou moins bien
réussie mais toujours présente et, à
l'intérieur de chacune d'elles, avec le temps, la
réalisation de l'idiomorphon caballin progresse. Le tableau
que, dans son livre Horses, Simpson (1951) propose de
l'évolution des Equoïdés nous agrée; il met
fort bien en évidence les tendances évolutives, et son
auteur fait de l'«orthogenèse» comme M. Jourdain
faisait de la prose, sans le savoir.
Il ne relève, dans cette évolution, qu'un seul
caractère apparaissant brusquement : le remplissage par du
cément (tissu osseux) des vallées que dessinent les
plis d'émail sur la face triturante des dents jugales du
Merychippus, Pliocène moyen d'Amérique du Nord. En
vérité ce caractère avait fait son apparition
plus tôt, en Europe à l'Oligocène, chez
Plagiolophus javali, un Palaeothéridé européen,
et s'insère dans le faisceau des particularités
caballines; chez Plagiolophus, il se trouve associé à
l'hypsodontie et à une très forte tendance à la
monodactylie : le doigt médian (III) l'emporte de beaucoup sur
les deux autres qui le flanquent.
Un tel ensemble de caractères, se manifestant des deux
côtés de l'Atlantique (plus tôt en Europe qu'en
Amérique), exclut toute intervention du hasard. Les
populations d'Equoïdés d'Europe et d'Amérique ont
réalisé une expérience naturelle dont il
convient de tirer les enseignements qu'elle comporte. Elle ne tourne
sûrement pas à l'avantage de la doctrine
darwinienne.
Au total, dans toutes les lignées d'Hippomorphes, au fur et
à mesure que les genres se succèdent : la taille
s'accroît, le massif facial l'emporte de plus en plus en volume
sur le neurocrâne, les dents gagnent en hauteur, le
cément apparaît, les replis d'émail se
compliquent, les membres s'allongent, le cubitus et le
péroné se réduisent à des vestiges
soudés respectivement au radius et au tibia, la
prédominance du 3ème doigt est un fait constant,
amorcée dans des familles qui, bien que tridactyles, sont
fonctionnellement monodactyles, elle s'exprime pleinement dans la
pure monodactylie des Equidés. L'idiomorphon caballin
présente bien d'autres particularités, parmi lesquelles
s'inscrivent la structure cérébrale, la suspension du
crâne, la forme générale du corps, etc., qui sont
présentes à des degrés inégaux dans les
diverses sous-lignées, et par cela même
"personnalisées". Dans son exposé théorique,
Simpson ne s'attarde pas à la structure du sabot et pourtant
elle est l'aboutissement d'une évolution toute d'invention et
de précision. Ce sabot, en harmonie avec le membre qu'il
coiffe à la manière d'un dé, protecteur de la
3ème phalange, amortit sans ressorts, ni caoutchouc, des chocs
dont la force peut dépasser une tonne. Il n'a pu se constituer
par hasard car, à l'examen, il se révèle
être un musée de coaptations et de nouveautés
organiques. La paroi cornée (muraille) par ses lames
verticales kératophylleuses s'engrène avec les lames
podophylleuses de la couche kératogène. Les longueurs
respectives des os, leur mode d'articulation, les courbes et formes
des surfaces articulaires, la texture des os (orientation, agencement
des travées osseuses), ligaments, tendons glissant dans des
gaines, coussinets amortisseurs, os naviculaire, synoviales et leur
liquide lubrifiant séreux impliquent une continuité
dans la construction que les hasards, forcément chaotiques et
incomplets, sont impuissants à maintenir. Cette
énumération ne concerne pas les ultrastructures
où brillent plus encore les adaptations, qui donnent leurs
solutions aux problèmes de mécanique des solides que
pose la locomotion rapide sur des membres monodactyles.»
Voici deux schémas issus du Précis de zoologie de
Pierre Paul Grassé chez Masson (1977) pour vous aider à
suivre ce texte :
 |
 |
|
d'une incisive de cheval adulte (in Précis de zoologie, Pierre Paul Grassé, Masson, 1977, modifié) |
avec quelques détails concernant la structure du sabot (in Précis de zoologie, Pierre Paul Grassé, Masson, 1977, peu modifié) |
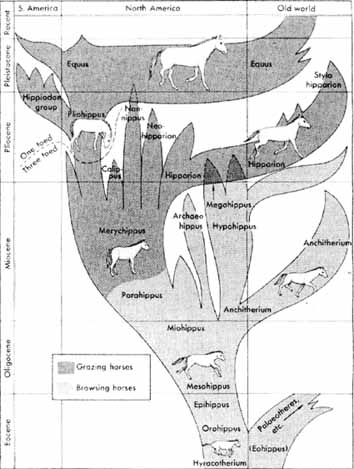
Voici la phylogénie présentée par Christine Janis dans Le Livre de la Vie, Seuil, 1993 et le texte qui l'accompagne :
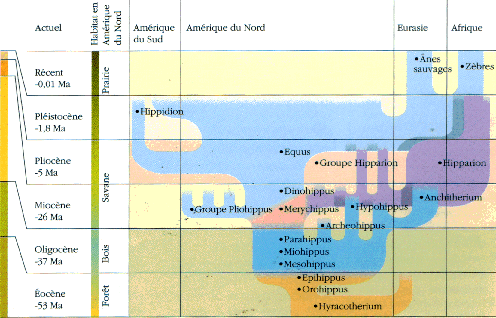
«L'Eocène ("l'aube des temps nouveaux") doit son
nom au fait qu'il a vu apparaître, entre -57,2 et -34 millions
d'années, beaucoup d'ordres actuels et même certaines
familles de mammifères vivant aujourd'hui.... Parmi les
premiers membres de l'ordre (…) des Périssodactyles, il y
a eu le tout premier des ancêtres du cheval, Hyracotherium.
… L'Eocène fut une époque particulièrement
faste pour les
périssodactyles. Ils se diversifièrent en toute une
série de familles, certaines d'entre elles survivant
aujourd'hui (chevaux, rhinocéros, tapirs), d'autres
étant éteintes (brontothères,
chalicothères, paléothères). …
A la fin de la période initiale de l'Eocène, les
températures à l'échelle planétaire
connaissent le pic le plus élevé de tout le Tertiaire.
Du milieu à la fin de l'Eocène, la tendance au
refroidissement et à la sécheresse fût plus
marquée au niveau des plus hautes latitudes, les gelées
d'hiver s'y manifestant vers la fin de cette époque, il y a 34
millions d'années. … A la fin de l'Eocène, les
zones de végétation de type tropical se
trouvèrent comprimées au niveau des latitudes
équatoriales, tandis qu'aux latitudes plus
élevées se développa un nouveau type de
végétation, la forêt tempérée
comprenant à la fois des conifères et des arbres
à feuilles caduques, semblable à la forêt
actuelle couvrant le Canada et le nord de l'Europe.…
Chez les herbivores, il se produisit un profond bouleversement, dans
le cadre duquel les ongulés à nombre pairs de doigts
prirent la tête. Leurs dents montrèrent de plus en plus
de signes d'adaptation à un régime alimentaire
contenant davantage de fibres, ce qui était la
conséquence des saisons plus marquées,
favorisant le
développement à long terme d'une
végétation devenue plus vieille et donc plus coriace,
au lieu de la production constante d'abondantes jeunes pousses.
… Les ongulés à nombre impair de doigts, les
périssodactyles, qui avaient été les herbivores
dominants au début et au milieu de l'Eocène, gardaient
encore une grande importance au sein de la faune, malgré
l'essor de nouveaux types d'ongulés à nombre pairs de
doigts. Il y avait en Europe, de nombreuses sortes
d'équidés primitifs et de paléothères,
apparentés au cheval, bien que le nombre des
équidés ait globalement décliné en Asie
et en Amérique du Nord.(Ils rebondirent à la fin de
l'Eocène en Amérique du Nord, avec l'apparition de
membres plus herbivores et de plus grande taille, de la sous-famille
des Anchitheriinae, des brouteurs spécialisés, tels que
Mesohippus).
(L'Oligocène) Apparemment, ce fut une époque de
stase évolutive,
une accalmie entre les moments d'apparitions et d'extinctions
d'espèces au début du tertiaire, et les
nouvelles radiations et
migrations de types plus modernes de mammifères
vers la fin du tertiaire. … Les nombreux équidés
d'Amérique du Nord étaient des brouteurs forestiers de
la taille du poney, comme Miohippus et Anchitherium ; mais il n'y
avait pas de grands brouteurs d'herbes dans les plaines.
Commençant à -23 millions d'années, le
début du Miocène vit s'amorcer un changement en
direction d'un climat plus chaud et considérablement plus sec.
C'est à peu près à se moment que s'ouvrit le
détroit de Drake entre l'Antarctique et l'Amérique du
Sud, livrant le passage à un courant circumpolaire d'eau
froide.… Un facteur supplémentaire fut le
rétrécissement des mers intérieures peu
profondes sur les continents, par exemple, la Thethys fut
fermée par un pont de terre entre l'Afrique et l'Eurasie, ce
qui isola la Méditerranée. … Entrant pour la
première fois en contact avec le continent eurasiatique, la
faune africaine, jadis unique en son genre, en fut
irrévocablement
transformée. Beaucoup d'espèces
africaines originales purent pénétrer en Eurasie, et en
particulier les proboscidiens et les hominoïdes (les grands
singes) en train de se répandre. La plus belle réussite
évolutive fut présentée par les
équidés. Les premiers d'entre eux, ancêtres du
cheval actuel, surgirent au début du Miocène. Il y eut
une immense radiation de diverses espèces de Merychippus, des
équidés dotés de plus longues pattes et de
molaires à couronne plus
élevée, capables de faire
face à un habitat plus ouvert, et à une
végétation plus rude, promue par les changements
climatiques du début du Miocène. Du début au
milieu du Miocène, la totalité de
l'hémisphère nord vit se développer
une tendance selon laquelle les animaux
adaptés à des habitats plus ouverts augmentèrent
en nombre, au détriment de ceux plus adaptés aux
forêts. ...
En Amérique du nord, les équidés plus
spécialisés et les animaux de la famille du cerf
surclassèrent les oréodontes. Pourtant il n'y avait pas
encore de traces de vraies prairies ouvertes. Ces herbivores
paraissaient adaptés à un régime alimentaire
comprenant à la fois des feuilles et des herbes,
peut-être avec des périodes de sécheresse
saisonnière, mais ils n'étaient pas encore de vrais
brouteurs d'herbe, comme les chevaux modernes et les
antilopes....
Une caractéristique fondamentale à la fin du
Miocène fut l'apparition en Amérique du Nord et du Sud
de prairies du type des savanes, l'un des événements
capitaux de ces derniers 500 millions d'années...... Les
animaux capables de manger des herbes
ont ainsi l'assurance d'une ressource alimentaire qui s'autoremplace
en permanence et qui peut se répandre sur presque des
centaines de kilomètres carrés sans
interruption. Le revers de la médaille est que
les feuilles contiennent beaucoup de silice, un minéral
abrasif, et leur consommation use les dents beaucoup plus vite que
celle des feuilles des arbres. La
réponse
évolutive des animaux brouteurs d'herbe a
été de développer
des dents hypsodontes, c'est-à-dire des dents
dont les couronnes sont plus hautes qu'elles ne sont longues ou
larges, avec d'épaisses couches de cément et
d'émail très repliées. Donc, une
conséquence de la
présence de ces particules de silice (phytolithes) a
été de stimuler
l'évolution des animaux capables de brouter les herbes de
façon si adaptée qu'ils en sont pratiquement
dépendants - à la façon d'un
marché captif. (Les changements dans la flore de l'intestin
ont été également importants ; ces bataillons en
sabots qui peuplent la savane dépendent fondamentalement de
leur estomac pour leur survie). ... Les faunes
nord-américaines développèrent une grande
quantité d'équidés manifestement adaptés
à un régime alimentaire composé presque
exclusivement d'herbe, comme le sont le cheval ou le zèbre
actuel. L'équidé à trois doigts Hipparion
traversa le détroit de Béring pour devenir l'un des
membres les plus importants des faunes de l'Ancien Monde.
... Bien que le grand échange entre les faunes du nord et du
sud de l'Amérique n'ait pas commencé avant le
Pliocène, ces continents étaient, à la fin du
Miocène, suffisamment proches pour que les premiers pionniers
franchissent par hasard
les eaux les séparant....
Au début du Pliocène, il y a environ 5 millions
d'années, les tendances au climat planétaire plus froid
et sec avaient imposé des conditions très semblables
à celles d'aujourd'hui, et ce fut à ce moment là
que les nouveaux grands types de végétation courants
aujourd'hui commencèrent à apparaître. Au
début du Pléistocène, il y a environ 1,8
millions d'années, le monde entra dans une période plus
froide d'alternance de glaciations et de phases interglaciaires.
Même maintenant, dans l'actuelle phase interglaciaire, le
climat est plus froid et sec qu'à l'époque du
Pliocène....
Dans la gamme des équidés aux différentes
mœurs alimentaires, seuls les brouteurs d'herbe
survécurent en Amérique du Nord...
L'Amérique du Nord a reçu, en provenance du Vieux
Monde, le cerf et le bison, aussi bien que des mammouths et des
félins aux dents de sabre, qui ne persistèrent pas au
delà du Pleistocène. Les chameaux, les canidés
et les vrais chevaux (brouteurs d'herbe et dotés de pattes
à un seul doigt) migrèrent dans l'autre sens,
c'est-à-dire à partir de l'Amérique du Nord. ...
Et finalement, à la fin du Pléistocène,
l'être humain (Homo sapiens) quitta l'Afrique pour se
répandre sur les autres continents du monde entier.
A la fin du Pléistocène, les types de mammifères
suivants étaient tous éteints : tous les grands animaux
porteurs de trompe, autres que les éléphants, dans le
monde entier, et les éléphantides (mammouths) en
Amérique du Nord et dans le nord de l' Eurasie ; les
félins aux dents de sabre, les hyènes ressemblant au
guépard, les canidés ressemblant aux hyènes ;
les sivathères girafidés, les chalicothères et
les damans géants. L'Amérique du Sud perdit ses
marsupiaux carnivores, ses ongulés indigènes
(notongulés et lipoternes) et ses grands édentés
(paresseux terrestres et glyphodontes) ; l'Amérique du Nord
perdit les mêmes grands édentés, présents
en tant qu'immigrants, de même que ses chevaux, chameaux et
tapirs. En Australie, les kangourous mangeurs de feuilles, les
dirpodontides et le lion marsupial disparurent (bien que des
récits et des contes aborigènes suggèrent que
certains de ces types ont peut-être perduré
jusqu'à des époques plus récentes). Les faunes
du monde actuel sont lourdement réduites, comparées
à celles d'il y a seulement 12.000 ans, et surtout en
matière de grands mammifères. Aujourd'hui, il n'y a
plus guère que l'Afrique, à avoir conservé
quelque chose du nombre et de la diversité des ces grands
mammifères qui s'étaient épanouis jusqu'à
récemment partout dans le monde. » Christine Janis,
Victoire par forfait : La succession mammalienne, dans Le Livre de la
Vie, Seuil, 1993, p 169-217
Présenter une histoire vraie, certaine, qui fasse
l'unanimité, n'est pas à la portée de l'homme et
va à l'encontre des principes des méthodes scientifique
expérimentale et historique. Chacun a cependant le droit
d'être convaincu de la justesse de sa vision et d'essayer de
convaincre le lecteur. Je ne suis pas un observateur impartial, mais
j'essaye d'en être conscient. Ces deux textes ne sont pas
choisis au hasard, l'un est plus axé sur la
paléontologie, plus proche des fossiles, alors que l'autre est
plus dirigé vers le grand public et raconte une histoire (un
conte ?). L'un est peut-être plus pédagogique que
l'autre, mais justement, c'est parce qu'il sous-tend une
théorie (modèle interne), facilement assimilable, et
répétée de nombreuses fois au cours du discours.
J'ai essayé d'en repérer quelques
éléments significatifs que j'ai fait ressortir
en rouge dans le texte. Par contre le
texte de Pierre-Paul Grassé est plus polémique car il
s'attaque à une théorie établie - au niveau
universitaire alors et dans le grand public de nos jours - et il ne
présente pas de théorie (modèle interne) : il
reste au niveau du modèle externe.
Résumé en quelques mots chaque théorie est un
exercice périlleux et forcément injuste (partial) mais
il faut quand même essayer de dégager les grandes lignes
de chacune:
* Pierre-Paul Grassé part d'un type caballin
(idiomorphon) actuel et voit une évolution
orientée par l'acquisition progressive de
caractères de plus en plus proches de ceux du cheval dans des
groupes diversifiés qui apparaissent puis disparaissent. Il
met l'accent sur l'harmonie qu'il existe entre les nombreuses
fonctions réalisées chez un herbivore, placentaire,
coureur... qu'est le cheval.
Il rejette le hasard comme facteur d'évolution. Pour lui,
«le cheval n'est pas … le produit raté de hasards
successifs»(p 43). « Les tendances évolutives
sont si nombreuses qu'elles ne font nul obstacle à la
diversité... L'impression de désordre que laisse
parfois l'examen superficiel des faunes d'hier et d'aujourd'hui
s'efface quand on approfondit l'étude des formes, qu'on
précise la chronologie et qu'on découvre les
lignées évolutives » (p 27).
* Christine Janis explique avant tout les êtres vivants
passés comme des individus soumis aux conditions du milieu
(climat, végétation et donc disponibilité
nutritive, répartition des terres émergées...)
et suppose que les changements du milieu imposent les modifications
chez les êtres vivants qui s'adaptent ou dont certaines
caractéristiques, apparaissant au hasard, sont
sélectionnées. Cette vision repose sur la
sélection naturelle qui s'impose à la suite de
la fécondité excessive et la
variabilité spécifique, ce qui est la
théorie darwinienne de l'évolution.
Une critique importante de cette théorie concerne l'action de
cette supposée sélection naturelle orientée de
façon similaire à des moments et des lieux
différents (Pierre-Paul Grassé - p103) : comment la
sélection naturelle a-t-elle pu aboutir à plusieurs
formes identiques en même temps (ou avec un décalage de
quelques millions d'années) en des lieux très
éloignés et n'ayant pas exactement les mêmes
conditions de milieu ? Pour en rester à l'exemple des
équidés: il est tout à fait faux de dire que le
cheval est le résultat d'un grand nombre d'adaptations, qui
seraient apparues une seule fois, et qui s'additionnent dans le temps
: l'hypsodontie, les sabots, la perissodactylie.... et d'innombrables
caractères de ce type sont apparus à des moments
différents, en des lieux différents et dans des
lignées différentes. Mais pas dans des embranchements
différents... il faut donc remonter au-delà de
l'espèce, de l'ordre et de la famille pour essayer de
comprendre les mécanismes de l'évolution. Faire jouer
la sélection naturelle au niveau de l'embranchement me
paraît donc envisageable mais par au niveau de l'espèce
qui est déjà un membre trop spécialisé de
l'embranchement.
Les faits paléontologiques considérés sont
les genres présentés plus haut.
On représentera sur la même trame que la
phylogénie de Christine Janis les différents
genres par des traits verticaux. Le début de chaque
trait correspondant à l'âge du fossile le plus ancien;
la fin du trait indiquant l'âge du fossile le plus
récent.
Les traits noirs sont donc les faits paléontologiques;
tous les traits bleus sont des
traits de construction....
J'ai relié par des traits pointillés horizontaux et
verticaux les genres identiques trouvés dans des
localités différentes (Hyracotherium et
Equus). Cela signifie que je pense qu'un genre donné
n'est apparu qu'une fois (monophylétisme) en un lieu
donné, même s'il a pu migrer ensuite et conquérir
d'autres territoires. Il ne faut pas confondre ce trait de
construction avec des traits de parenté (continus) que nous
allons ensuite nous efforcer de placer.
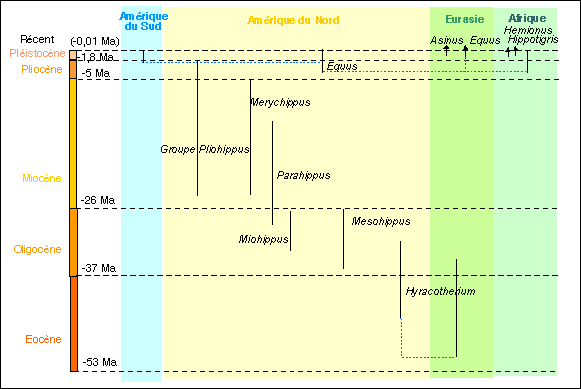
Le travail consiste maintenant à établir des
filiations entre ces genres. Même si les données sont
très incomplètes, la méthode peut cependant
être appliquée dans un but pédagogique.
* Pour les genres d'équidés actuels (Asinus, Hemione
et Hippotigris) on peut raisonnablement penser qu'ils
dérivent du cheval (Equus) au cours du Quaternaire. On
les relie donc à l'aide de traits. Dans une
représentation moderne on choisit habituellement des traits
horizontaux qui indiquent que la différenciation en genres
nouveaux est un phénomème rapide. Lorsque plusieurs
genres apparaissent en même temps à partir d'un
même type, on parle de radiation. Dans la théorie
évolutive la plus en faveur actuellement on pense que ces
étapes de radiation sont séparées par des
périodes de stabilité (stase). C'est le
modèle "saltationniste" ou modèle des
équilibres ponctués développé
notamment par Stephen Jay Gould. A la fin d'une période de
stabilité les genres dispariassent: c'est la phase
d'extinction.
Pour visualiser ces périodes de radiation il est parfois
nécessaire d'avancer sur le schéma la date d'apparition
d'un genre. Lorsque les données des fossiles viennent ensuite
confirmer cette antériorité d'un genre, c'est un bon
argument en faveur du modèle (qui permet de prévoir)
mais il est aisé de comprendre qu'il ne constitue en rien une
preuve.
* L'origine du cheval demande ensuite par exemple à être
discutée. On peut le faire dériver de genres encore
présents au Pliocène (représentation choisie
ci-dessous puisque je n'ai utilisé que des traits horizontaux
et le minimum de traits verticaux) ou de genres intermédiaires
ancestraux, non encore découverts.
* Un exemple de branchement d'un genre à partir d'un autre
assez bien documenté est celui de Miohippus à
partir de Mesohippus, à l'Oligocène.
Brusquement, le genre Miohippus apparaît et coexiste
avec Mesohippus.
Voilà le schéma précédent
complété dans cette optique.
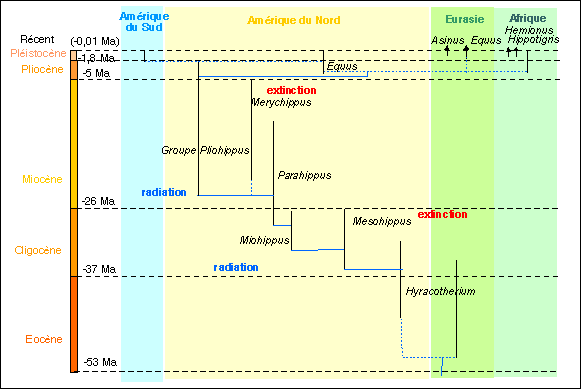
Cependant, il ne faut pas oublier que ce schéma n'a aucune valeur paléontologique étant donné qu'il est incomplet (et imprécis).
Conclusion:
l'étude de cet exemple a au moins les mérites
suivants:
- faire apparaître la théorie sous-jacente
à l'interprétation qui est cachée alors qu'elle
me semble être le plus important (et pourtant le moins
apparent)... ; Aucune théorie n'est définitive dans le
cadre de l'histoire de la vie... peut-être même cette
histoire est-elle définitivement hors d'atteinte ?
- mettre en évidence le point central des discussions entre
paléontologues: les classifications évolutives ou
phylogénies qui reposent sur la notion
d'espèce paléontologique, ce que nous allons
donc étudier maintenant.
Remarques: