Histoire et évolution de la
terre et des êtres vivants
suite
retour plan cours
/ partie I : génétique / partie
II : immunité / partie III : physiologie
/ partie IV : évolution
programme commenté de cette partie
|
page précédente
|
|
|
plan de cette page
|
- 3.L'évolution
- a . Les différentes théories
évolutives doivent toujours être
enseignées et non une seule
- b. Une brève histoire des concepts
d'évolution
- c. Panorama simplifié sous forme de petit
vocabulaire évolutif
- d. Vers d'autres théories
- d1. Dans la recherche des causes de
l'évolution, l'opposition n'est pas entre
finalisme et mécanisme mais
plutôt entre vitalisme et
mécanisme.
- d2. Les critiques de Pierre-Paul
Grassé
- d3. Des critiques au sujet de la
sélection naturelle
- d4. Une théorie de l'évolution
qui est une théorie globale du vivant par
Rosine Chandebois
- d5. Une autre théorie non darwinienne:
celle d'Anne Dambricourt sur l'apparition de
l'homme
- 4.Phylogénies
- a . Les phylogénies ou
classifications évolutives classent... des
caractères.
- a1. Caractères anatomiques
- a2. Caractères embryologiques
* Ernest Haeckel , la loi
biogénétique fondamentale et la
sélection naturelle
* les hétérochronies
- a3. Caractères moléculaires,
génétiques et chromosomiques
a31. les ressemblances entre deux molécules
sont chiffrées par la distance
génétique.
a32. la comparaison des génomes
associés à la synthèse de
molécules homologues chez des organismes
apparentés permet de retracer une histoire
(dans le cadre de l'hypothèse
évolutive)
a33. la comparaison des caryotypes chez des
organismes apparentés permet de proposer des
hypothèses concernant la spéciation
(formation de nouvelles espèces) par
isolement reproducteur
- b. Les phylogénies cladistes
(d'après la méthode de Willi Hennig)
classent des caractères polarisés
- c. On peut faire des classifications
évolutives qui ne soient pas des
cladogrammes
- d. L'espèce est à la base de toute
classification ou dénombrement du monde vivant
- d1. le problème philosophique de
l'espèce
- d2. l'espèce vivante (biologique)
- d3. l'espèce morte
(paléontologique)
- d4. la spéciation: vers une
définition évolutive de
l'espèce
- 5. Les grandes lignes de l'histoire de
la vie
- a. L'échelle des temps
géologiques
- b. La crise
Crétacé-Paléocène (ou la
limite K-T)
- c. Les grandes dates de l'évolution du
vivant
|
|
page suivante
|
|
(pour la bibliographie voir page
précédente)
3. l'évolution
Le temps des querelles de noms est sans aucun doute
dépassé : le terme «évolution» est
pris dans ces pages au sens moderne d'« histoire de la
vie». L'évolution c'est
l'histoire de l'écosystème terrestre.
C'est une certitude pour la plupart des personnes, dont je fais
partie, même si elle ne relève pas de la science
expérimentale.
a . Les différentes théories évolutives
doivent toujours être enseignées et non une seule
Pourquoi ne plus parler de théories évolutives ? Si
l'évolution relève d'un consensus les théories
restent très débattues.
La réponse est loin d'être simple car des questions de
politique, d'argent, d'influences de différents
laboratoires... bref des questions "humaines", non seulement se
greffent mais tendent à occulter les différentes
positions théoriques des protagonistes (on se reportera au
magazine La Recherche et parfois aussi à Pour la
Science pour suivre le débat de façon
épisodique...). Le programme officiel - et les manuels
scolaires qui doivent s'y conformer- ne demande de ne
présenter qu'une seule théorie,
sans la citer (le nom de Darwin n'apparaît
pas une seule fois dans le programme et dans les manuels à ma
connaissance) et donc en la présentant comme une
vérité "scientifique"(c'est moi qui interprète
ce non-dit mais je suis sûr de ne pas me tromper
complètement). Ceci n'est pas acceptable. L'idéologie
politique «dénoue les liens du passé au
présent et à l'avenir, ce qui est le propre du
mensonge» (j'emprunte cette phrase à
Marguerite Léna dans un ouvrage sur l'éducation
dont j'ai présenté quelques idées sur une page
de ce site): l'idéologie politique masquée est
mensonge et un programme d'enseignement aussi tronqué
participe à mon avis d'une idéologie politique
pragmatique.
J'ai déjà dit dans les
pages sur la science combien j'étais conscient de ne pas
être impartial mais qu'à mon sens cela n'empêchait
pas l'enseignant de présenter une question avec cœur en
d'essayer de montrer plusieurs aspects des différentes
théories. Ma vision est donc certainement personnelle mais je
ne vois pas d'autre solution que de présenter le cours de
cette manière. Il est sûr que lors d'un examen,
forcément unifié, cette diversité demandera une
certaine ouverture d'esprit de la part des correcteurs, mais il faut
miser dessus, sinon il n'y a plus de liberté
pédagogique.
b. Une brève histoire des concepts d'évolution
L'évolution, comme elle est comprise de nos jours, est en
fait historiquement le
transformisme
(même si ce terme n'a été
introduit par Broca qu'en 1867 et son usage ne devint courant, comme
celui d'évolution, que vers les années
1870).
Le transformisme est la théorie selon laquelle les
êtres vivants dérivent les uns des autres au cours des
temps géologiques par une série de transformations.
Le fondateur du transformisme et donc de l'évolution,
au sens moderne de théorie scientifique, est
indiscutablement Lamarck.
L'évolutionnisme est une
doctrine philosophique dont les sens ont été
fluctuants au cours de l'histoire. Actuellement c'est «une
philosophie du développement graduel et continu de la
nature et de l'humanité, animé par un principe interne
et orienté vers le progrès. En ce sens, Saint
Augustin est évolutionniste et non transformiste; Lamarck,
Spencer, Bergson, Teilhard de Chardin sont à la fois
évolutionnistes (comme philosophes) et transformistes (comme
savants); Darwin est transformiste et non évolutionniste. Les
biologistes contemporains, même lorsqu'ils se disent
évolutionnistes, sont en réalité transformistes:
ainsi J. Monod, qui attaque, comme non scientifiques, les
philosophies évolutionnistes (Le Hasard et la
Nécessité ). Dans ces conditions, et malgré
l'usage établi, on peut regretter que
«transformisme», qui est univoque, disparaisse devant
«évolutionnisme», qui demeure ambigu.»
(Pour plus de détails voir l'article
de Jacques Roger)
Le terme
d'évolution
n'apparaît que dans la sixième édition de
L'origine des espèces (Darwin) et il ne sera pour la
première fois (?) pris dans le sens de transformisme,
théorie scientifique, par Spencer (qui n'était pas
transformiste) puis par de nombreux autres auteurs dont le plus connu
fût peut-être Thomas Huxley. Ces auteurs étant
vite qualifiés à tort de darwiniens ou
évolutionnistes, par opposition aux non-darwiniens, non
évolutionnistes, ce qui est historiquement et
philosophiquement faux. (voir par exemple deux
articles de l'E.U. sur l'historique du concept: un article
de Jacques Roger et un article
de Pierre-Paul Grassé ou La
naissance du transformisme, Lamarck, entre Linné et
Darwin, Goulven LAURENT, Collection inflexions, Vuibert/Adapt,
2001, 22-23).
Le transformisme s'oppose au
fixisme.
Le fixisme est la théorie selon laquelle les
organismes vivants ne se transforment pas et ne dérivent pas
les uns des autres mais sont apparus tels quels au cours des temps
géologiques et que chaque espèce forme un
système indépendant: tout individu d'une
espèce étant issue d'un individu de cette même
espèce sans qu'il n'y ait jamais de passage d'une
espèce à une autre (spéciation). Cette
doctrine n'étant plus acceptée par aucun biologiste,
à moins que cela ne soit pour des raisons philosophiques ou
religieuses.
Remarque: Certaines personnes mal
informées (ou peut-être parfois mal
intentionnées) confondent fixisme et
créationnisme qui devrait désigner une
croyance (religieuse) selon laquelle tous les êtres vivants
ont été créés individuellement par
Dieu. Le problème est que ce dogme de la création
par Dieu ne devrait pas être appelé
créationnisme, il dépasse la doctrine philosophique,
c'est une foi. C'est la reconnaissance de la dépendance
absolue de chaque être vivant à chaque instant envers
son Créateur qui le maintient dans l'être. C'est
plutôt ce que l'on pourrait appeller une
création continuée. En utilisant ce
sens de "croyant en une création individuelle des
êtres vivants", on peut être à la fois
créationniste et transformiste (comme le propose
actuellement l'église catholique) mais aussi à la
fois créationniste et fixiste (comme le sont quelques
groupes protestants, notamment aux États Unis,
d'après ce que je crois savoir).
Pour simplifier on pourrait dire que, historiquement et
philosophiquement, c'est Lamarck qui a
développé le premier le
transformisme
contre Cuvier qui était
fixiste.
|
Jean-Baptiste de Monet de
LAMARCK (1744-1829)
Jean-Baptiste Pierre Antoine de Monet, chevalier de
Lamarck, naquit à Bazentin-le-Petit (Somme); il
quitta le collège de Jésuites d'Amiens - et un
avenir écclesiastique - dès la mort de son
père, en 1760, pour prendre rang dans l'armée
où il brilla; mais il dut en 1968, à la suite
d'un accident, renoncer au métier des armes.
Pendant quatre ans, ne disposant que d'une maigre pension
complétée par un appointement de petit
employé de banque, Lamarck poursuivit à Paris
des études médicales et botaniques (sous la
direction de Bernard de Jussieu). Il publia, en 1778,
grâce à la protection de Buffon, une Flore
française qui lui valut une gloire
immédiate, concrétisée l'année
suivante par son élection, à trente-cinq ans,
à l'Académie des sciences.
Il est chargé, grâce à une intervention
de Buffon, des humbles fonctions de gardien de l'herbier du
Jardin des plantes. Lors de la fondation du Muséum
national d'histoire naturelle par la Convention (10 juin
1793), il obtint, à cinquante ans, la Chaire
d'enseignement des Insectes et des Vers. C'est lui qui
devait créer le terme d'Invertébrés et
fonda non seulement la zoologie des
Invertébrés mais aussi leur
paléontologie. Dès ses premiers cours au
Muséum en 1794, sa distinction entre Animaux à
vertébres et Animaux sans vertèbres,
connût un immense succès. Cuvier, par exemple,
employait ces termes dans ses Leçons d'Anatomie
Comparée, dès 1800.
Le labeur colossal qu'il accomplit (« ce que
Linné avait fait pour les plantes, Lamarck l'a fait
pour les invertébrés ») le conduisit
à publier en 1801 la première édition
du Système des animaux sans
vertèbres qu'il devait réviser sans
cesse. Sa cécité commença vers 1818 et
c'est elle qui eut raison de son souci d'améliorer
sans cesse son travail. Ses premières formulations
transformistes sont introduites dans son Discours
d'ouverture de son cours de l'an VIII (1800) dont
ses autres publications sur le sujet ne sont que des
développements (Recherches sur l'organisation des
Corps vivans, 1802; Philosophie zoologique
(1809); Introduction (1815) à
l'Histoire naturelle des Animaux sans vertèbres
(1815-1822). De très nombreux élèves, y
compris étrangers, suivaient ses cours et il
était consulté par de très nombreux
zoologistes, qui ,de plus en plus nombreux, se
lançaient dans l'étude des
Invertébrés, maintenant qu'il leur
était facile de le faire, grâce à la
classification ingénieuse et claire que leur
fournissait Lamarck.
C'est une légende récente qui raconte qu'il
est mort pauvre (alors qu'il touchait ses émoluments
d'académicien et de professeur du Muséum
jusqu'à sa mort à 85 ans
(Laurent. p 134), un grand
âge pour l'époque) et oublié (alors que
sa renommée touchait tout le monde occidental).
Certes, il était peu aimé de certains, dont
Cuvier, mais cela ne justifie pas la légende.
Lamarck proposa le premier le
transformisme
comme explication de toute classiffication
signifiante du monde vivant et ne cessa par la suite
d'apporter des preuves palontologiques et zoologiques. Il le
développa contre le fixisme ambiant.
Comme tout scientifique d'immense valeur, il chercha
à développer une véritable
réflexion philsophique sur sa science. C'est ainsi
qu'il utilisera pour la première fois
(Hydogéologie, 1802, p 7-8) et définira
le mot biologie comme une nouvelle science
unifiée (matérialiste et
évolutionniste) de tous les êtres vivants:
« elle comprend tout ce qui a rapport aux corps
vivans, et particulièrement à leur
organisation, à ses développemens, à sa
composition croissante avec l'exercice prolongé des
mouvemens de la vie, à sa tendance à
créer des organes spéciaux, à les
isoler, à centraliser l'action dans un foyer,
etc.» (Recherche sur l'organisation des Corps
vivans, 1802, p. 202 in La naissance du
transformisme, G. Laurent, Vuibert/Adapt, col.
inflexions, 2001, p 110)
Quatre lois (in Charles Bocquet)
régissent selon Lamarck l'organisation des
individus (1816):
- «Première loi : La vie,
par ses propres forces, tend continuellement
à accroître le volume de tout corps qui la
possède, et à étendre les dimensions
de ses parties, jusqu'à un terme qu'elle
amène elle-même.
- Deuxième loi : La production d'un
nouvel organe dans un corps animal
résulte d'un nouveau besoin
survenu qui continue de se faire sentir et d'un nouveau
mouvement que ce besoin fait naître et
entretient.
- Troisième loi : Le
développement des organes et
leur force d'action sont constamment en raison
de l'emploi de ces organes.» On
retrouve là cet effet de l'usage et du non-usage
que Lamarck avait défini dans sa Philosophie
zoologique : «Dans tout animal qui n'a
point dépassé le terme de ses
développements, l'emploi plus fréquent et
soutenu d'un organe quelconque fortifie peu à peu
cet organe, le développe, l'agrandit et lui donne
une puissance proportionnée à la
durée de cet emploi; tandis que le défaut
constant d'usage de cet organe l'affaiblit
insensiblement, le détériore, diminue
progressivement ses facultés, et finit par le
faire disparaître.
- Quatrième loi : Tout ce qui a
été acquis,
tracé ou changé, dans l'organisation des
individus, pendant le cours de leur vie, est
conservé par la
génération, et
transmis aux nouveaux individus qui
proviennent de ceux qui ont éprouvé ces
changements.»
Je préfère à cette vision
réductrice du Lamarckisme les principes
philosophiques qu'il expose dans son Discours d'ouverture
du cours de 1814 (Inédits de
Lamarck, Vachon et alii, 1972, p. 181, in La
naissance du transformisme, G. Laurent, Vuibert/Adapt,
col. inflexions, 2001, p 120-121):
- 1er principe: tout fait ou phénomène
que l'observation peut faire connoître est
essentiellement physique, et ne doit son existence ou sa
production qu'à des corps, ou qu'à des
relations entre des corps.
- 2e principe: tout mouvement ou changement, toute
force agissante, et tout effet quelconque,
observés dans un corps tiennent
nécessairement à des causes
mécaniques, régies par des lois.
- 3e principe: tout fait ou phénomène
observé dans un corps vivant, est à la fois
un fait ou phénomène physique, et un
produit de l'organisation.
- 4e principe: il n'y a dans la nature aucune
matière qui ait en propre la faculté de
vivre. Tout corps en qui la vie se manifeste, offre dans
le produitd 'organisation qu'il possède, tds
ceid'un uite de mouvemens excités dans ses
parties, le phénomène physique et organique
que la vie constitue, phénomène qui
s'éxécute et se maintient dans ce corps,
tant que les conditions essentielles à sa
production subsistent.
- 5e principe: il n'y a dans la nature aucune
matière qui ait en propre la faculté
d'avoir ou de se former des idées,
d'éxécuter des opérations entre les
idées, en un mot, de penser. Là où
de pareils phénomènes ses montrent (et l'on
en observe de cette redn e nimaux les plus parfaits),
l'on trouve tjours un système d'orgnepariclier
propre à les produire ; système dont
l'étendue et l'intégrité sont
constamment en rapport avec le degré
d'éminence et l'état des
phénomènes dont il s'agit.
- 6e principe: enfin, il n'y a dans la nature aucune
matière qui ait en propre la faculté de
sentir. Aussi, là où cette faculté
peut être constatée, là seulement se
trouve, dans le corps vivans qui en est doué, un
système d'organes particulier capable de donner
lieu au phénomène physique,
mécanique et organique qui, seul, constitue la
sensation.
(Charles Bocquet in E.U. article
"Lamarck" corrigé par La naissance du
transformisme, Lamarck, entre Linné et
Darwin, Goulven LAURENT, Collection inflexions,
Vuibert/Adapt, 2001 )
|
|
La conception de la vie de Lamarck (qui fonde un
véritable philosophie de la vie) versus celle
de Cuvier
|
|
Lamarck, dont la philosophie était un
matérialisme (voir G.
Laurent), conçoit la vie comme un
mécanisme (G. Canguilhem, dans
son essai sur la
notion de milieu (et voir
paragraphe ci-dessous d1), décrit le
pseudo-mécanicisme lamarckien comme un "vitalisme nu"
et dans l' E.U.( article
"vie") , il rejette plus ou moins
le qualificatif de "matérialiste" attribué
à Lamarck): l'accumulation et
l'intériorisation continues et progressives de
mouvements de fluides dans les solides, sous la forme
initiale d'un tissu cellulaire, «gangue dans
laquelle toute organisation a été
formée». «... dans la
réalité la vie n'est qu'un mode ou qu'un ordre
de choses relatives dans lequel tout changement qui
détruit cet ordre, constitue ce que l'on nomme la
mort ».(Sur l'origine des
êtres vivans, 1801-1803, Inédits de
Lamarck, Vachon et alii, 1972, p. 181, in La
naissance du transformisme, G. Laurent, Vuibert/Adapt,
col. inflexions, 2001, p 113). «... pour
faire exister cette force vitale, et lui donner toutes les
propriétés qu'on lui connaît, la nature
n'a pas besoin de lois particulières; celles qui
régissent généralement tous les corps
lui suffisent parfaitement pour cet objet
» (Philosophie zoologique,
1809, t. 2, p. 96 in La naissance du transformisme,
G. Laurent, Vuibert/Adapt, col. inflexions, 2001, p 113)
: il n'y a plus de frontière entre la
connaissance du vivant et du non vivant; les êtres
vivants supérieurs viennent des inférieurs par
génération et les tout premiers se sont
organisés de façon spontanée à
partir du non vivant (complexification naturelle ou
mouvement du vivant), l'homme lui-même étant un
produit de cette complexification (voir
La naissance du transformisme, G. Laurent,
Vuibert/Adapt, col. inflexions, 2001, p 116-118 où
Lamarck est reconnu comme le premier scientifique à
proposer l'origine simiesque de l'homme). Cette
manière de concevoir le vivant est extrêmement
répandue de nos jours «toutes les
facultés, sans exception, sont complètement
physiques, c'est-à-dire, que chacune d'elles
résulte essentiellement d'actes de l'organisation ;
en sorte qu'il sera facile de montrer comment de l'instinct
le plus borné, dont la source peut être
aisément aperçue, la nature a pu parvenir
à créer les facultés de l'intelligence,
depuis celles qui sont les plus obscures, jusqu'à
celles qui sont les plus développées
» (Philosophie zoologique, t1, p
369-370 in La naissance du transformisme, G. Laurent,
Vuibert/Adapt, col. inflexions, 2001, p 118).
Tout autre est la conception de Cuvier. (...)
«La vie est un tourbillon continuel dont la
direction, toute compliquée qu'elle est, demeure
constante, ainsi que l'espèce des molécules
qui y sont entraînées, mais non les
molécules individuelles elles-mêmes; au
contraire la matière actuelle du corps vivant n'y
sera bientôt plus, et cependant elle est
dépositaire de la force qui contraindra la
matière future à marcher dans le même
sens qu'elle. Ainsi la forme de ces corps leur est plus
essentielle que leur matière, puisque celle-ci change
sans cesse tandis que l'autre se conserve»
(Histoire des progrès des sciences naturelles
depuis 1789 jusqu'à ce jour , 1810).
|
|
Georges CUVIER (1769-1832)
Georges Cuvier est né en 1769 à
Montbéliard, ville alors rattachée au
duché de Wurtemberg, d'une famille originaire d'un
village du Jura qui était venue s'y établir au
moment de la Réforme.
La lecture d'un ouvrage de Buffon, trouvé par hasard
dans la bibliothèque d'un de ses parents,
éveille en lui le goût de l'histoire
naturelle.
Après de brillantes études à
l'Académie de Stuttgart, il quitta cette ville en
1788 et devint précepteur dans une famille de
Normandie. Il passa ainsi les années de la
Révolution dans la campagne du pays de Caux, en
contact direct avec la nature, à peu près sans
livres. On peut penser avec Flourens qu'au cours de cette
période commencèrent à germer dans son
esprit les deux projets qui allaient marquer toute son
œuvre: comparer les espèces fossiles aux
espèces vivantes et refondre la classification du
règne animal, ce dont il eut l'idée en
récoltant des Térébratules et en
disséquant des Mollusques et autres animaux
marins.
À cette époque, il entre en relation avec
Tessier, médecin-chef de l'hôpital militaire de
Fécamp, qui, frappé de l'étendue du
savoir de Cuvier, en fait part à ses amis du jardin
des Plantes. En 1795, Cuvier est nommé
suppléant de Mertrud, alors chargé de
l'enseignement de l'anatomie comparée; en 1799, il
succède à Daubenton à la chaire
d'histoire naturelle du Collège de France; enfin, en
1802, à la mort de Mertrud, il devient professeur au
jardin des Plantes. Les honneurs et les charges n'allaient
cesser d'arriver: en 1803, il était promu
secrétaire perpétuel de l'Académie des
sciences pour les sciences physiques et naturelles; en 1808,
il entrait au conseil de l'Université; en 1813, il
devenait maître des requêtes. La Restauration
lui octroya des distinctions nouvelles; il fut conseiller
d'État, chancelier de l'Instruction publique et, en
1831, pair de France. Fait sans doute unique, il appartenait
à trois académies de l'Institut de France:
l'Académie française, celle des sciences et
celle des inscriptions, et il était membre de toutes
les académies savantes du monde.
En dépit de ces tâches multiples, Cuvier put
mener à bien une œuvre scientifique qui est un
des monuments de l'histoire naturelle. (Jean
Piveteau in E.U. article "Cuvier")
Voir l'anecdote
sur le Didelphe dans la page
précédente qui
essaie de montrer l'importance des raisonnements de cet
homme qui fût un des premiers
paléontologues.
Voici en quelques mots la position
fixiste de Cuvier: «Pourquoi les races
actuelles ne seraient-elles pas des modifications de ces
races anciennes que l'on trouve parmi les fossiles,
modifications qui auraient été produites par
les circonstances locales et le changement de climat, et
portées à cette extrême
différence par la longue succession des
années?» À quoi on peut
répondre, déclarait-il, que, si les
espèces avaient changé par degrés, on
devrait trouver des traces de ces changements. Entre la
faune à Palaeotherium et la faune
à Mastodontes, entre la faune à Mastodontes et
la faune actuelle, on devrait trouver des
intermédiaires, «et jusqu'à
présent cela n'est point arrivé». Et
même si les espèces anciennes n'avaient pas
été fixes, les révolutions nombreuses
dont notre globe a été le théâtre
ne leur auraient pas laissé le temps de se livrer
à leurs variations.
|
A la suite de Lamarck, c'est Darwin qui développera
le transformisme, notamment grâce à un travail
extrêmement documenté sur les variations des
espèces (à l'occasion de son périple de 57 mois
(12/1931-10/1936) comme naturaliste embarqué sur un navire
d'observation scientifique: le Beagle). Son livre majeur sur
l'évolution: L'Origine des espèces parut
le 24 novembre 1859; Darwin y expose dans le détail sa
théorie de la sélection
naturelle: a theory of modification through natural
selection , a theory of descent with subsequent
modification .
|
Charles Robert DARWIN (1809-1882)
Darwin, né en 1809 à Shrewsbury, dans le
Shropshire, était le petit-fils d'un médecin,
physiologiste et poète. Il fréquenta
l'école de Shrewsbury et s'intéressa à
des collections variées (coquilles, médailles,
sceaux, franchises postales, plantes, minéraux), au
jardinage, à l'entomologie, à l'ornithologie,
à la poésie et à la peinture de
paysages, mais surtout à la chimie
expérimentale et à la pratique médicale
que lui inculqua son père. De 1825 à 1828, il
fréquenta l'université d'Édimbourg pour
y poursuivre des études de médecine auxquelles
sa sensibilité ne résista pas. Il rencontra
à cette période William MacGillivray, qui lui
apprit à naturaliser les oiseaux, et Robert Edmond
Grant, qui lui parla de Lamarck. Renonçant à
en faire un médecin, son père lui proposa, en
1828, de devenir clergyman, perspective qui, temporairement,
plut à Charles. Celui-ci entra donc à Christ's
College, à l'université de Cambridge, dont,
étudiant médiocre, il sortit cependant
bachelier ès arts trois ans plus tard; il y subit
l'influence de professeurs qui demeurèrent ses amis,
le botaniste John Stevens Henslow et le géologue Adam
Sedgwick. Il ne serait peut-être resté qu'un
simple whig fortuné si une occasion
fantastique, qui allait décider de toute sa vie, de
toute son œuvre, ne s'était
présentée à lui. Le capitaine Fitzroy
offrait une place de naturaliste à bord du
Beagle , qui devait compléter certains
relevés cartographiques et effectuer des mesures
chronométriques autour du monde. Pressenti par
Henslow, soutenu par son oncle Wedgwood contre les
réticences de son père, Charles Darwin devint
ce naturaliste privilégié.
Le Beagle quitta Devonport le
27 décembre 1831. Il regagna Falmouth le
2 octobre 1836. «Darwin visita Santiago, dans
les îles du cap Vert, Saint-Paul, Fernando de Noronha,
passa des mois au Brésil, à Bahia et autour de
Rio. Deux ans, le Beagle releva les côtes est et
sud de l'Amérique à partir de La Plata, et le
naturaliste parcourut l'Uruguay, l'Argentine, la Patagonie.
Il vit les îles Falkland et deux fois la Terre de Feu,
resta plus d'un an au Chili et au Pérou, escalada les
Andes, fit escale aux îles Galapagos, puis à
Tahiti, en Nouvelle-Zélande, en Australie, en
Tasmanie, à l'île Keeling, aux Maldives,
à l'île Maurice, au Cap, à
SainteHélène, à l'Ascension, à
nouveau au Brésil, au cap Vert et, enfin, aux
Açores» (M. Prenant, Darwin ).
Durant ces cinquante-sept mois, Darwin accumula une
multitude d'observations et réfléchit
longuement aux faits qu'il découvrait. Il exploita
dès son retour cette masse inestimable de
connaissances et de réflexions, en l'augmentant
continuellement par les renseignements qu'il
dégageait de ses lectures, par les informations
apportées par ses nombreux correspondants
(zoologistes, botanistes, éleveurs, horticulteurs).
Sa vocation de naturaliste étant cette fois
définitive, il devint secrétaire de la
Geological Society de 1838 à 1841 et se lia
d'amitié avec sir Charles Lyell, dont il admirait
grandement le livre The Principles of Geology ,
publié en 1831.
Il épousa, en janvier 1839, sa cousine Emma Wedgwood.
Quatre fils devaient naître de cette union. Darwin et
sa femme vécurent à Londres jusqu'en 1842,
puis s'installèrent à Down, dans le
comté de Kent, où ils menèrent une
existence très retirée. La santé de
Darwin, après son retour en Angleterre, resta en
effet toujours très précaire, au point de lui
faire redouter ou fuir les réunions et espacer les
visites. À l'abri de tout souci matériel,
grâce à la fortune dont il avait
hérité et au succès de ses livres,
habile aux bons placements et fort soucieux de ses revenus,
économe et pourtant généreux,
partageant sa vie régulière entre quelques
heures de travail quotidien, des promenades et des lectures,
élevant pour ses études des pigeons et des
lapins, cultivant des plantes pour ses expériences,
Darwin ne devait jamais quitter Down, où il
rédigea tous ses ouvrages fondamentaux. Il y
reçut cependant souvent ses amis, C. Lyell,
T. Huxley, J. Lubbock et de nombreux savants et
philosophes, toujours avec affabilité.
La gloire était venue avec l'âge. Darwin mourut
le 19 avril 1882; son corps repose à l'abbaye de
Westminster. (d'après Charles Bocquet
in E.U. article "Darwin")
Une définition de la
sélection
naturelle avec les propres mots de Darwin:
«Étant donné que plus d'individus sont
produits qu'il n'en peut survivre, il doit exister dans
chaque cas une lutte pour l'existence, soit entre un
individu et un autre individu de la même
espèce, soit entre individus d'espèces
différentes. Peut-on, dès lors,
considérer comme improbable, puisque des variations
utiles à l'homme sont manifestement survenues, que
d'autres variations utiles en quelque manière
à chaque être vivant, dans la grande et
complexe bataille pour la vie, se soient parfois produites
au cours de milliers de générations? S'il en
va ainsi, pouvons-nous douter (en nous rappelant que bien
plus d'individus naissent qu'il n'en peut survivre) que les
individus possédant un avantage quelconque, si minime
soit-il, sur les autres auraient une meilleure chance de
survivre et de procréer leur propre type?
Inversement, nous pouvons être assurés que
toute variation, délétère à
quelque degré, serait impitoyablement
éliminée. Cette préservation des
variations favorables et ce rejet des variations
défavorables, je l'appelle la sélection
naturelle.» C. Darwin, The Origin of Species by
Means of Natural Selection, or the Preservation of Favoured
Races in the struggle for Life (1859)
|
c. Panorama simplifié sous forme de petit vocabulaire
évolutif
|
Petit vocabulaire évolutif
(une première approche)
Ces définitions
sont expliquées avec mes propres mes mots et non ceux
des auteurs de ces théories. De plus ces mots sont
pris dans leur sens moderne et non historique. D'où
à mon avis le côté caricatural de telles
définitions. Mais je les donne pour essayer d'aider
les élèves, comme un point de départ
à dépasser... lorsqu'ils en auront le
loisir.
Dans ce tableau théorie fait
davantage référence à une philosophie
sous-jacente que le terme de
modèle, plus neutre.
|
|
deux anciennes théories évolutives
dérivées de la pensée de deux
scientifiques
|
lamarckisme
|
Théorie évolutive (empruntée
à Lamarck, 1744-1829) selon laquelle il existe
une graduation, les organismes tendant naturellement vers
une complexité de plus en plus poussée ; les
besoins font naître de nouveaux organes; les organes
sont perpétuellement modelés en fonction de
leur usage; les modifications organiques sont
héréditaires
|
|
darwinisme
|
Théorie évolutive (empruntée
à Darwin, 1809-1882) selon laquelle les
populations naturelles se reproduisent en nombre excessif et
subissent des modifications dues au hasard; un certain
nombre d'individus doivent donc être
éliminés par la sélection naturelle ;
il y a hérédité des caractères
acquis par l'usage.
|
|
"L'hérédité des caractères
acquis" est une formulation que l'on trouve chez Darwin
et non chez Lamarck et qui désigne une vision
courante à l'époque selon laquelle
«les êtres vivants transmettent à leurs
descendants les caractères qu'ils ont acquis durant
leur existence par l'usage ». Darwin proposa comme
hypothèse ce que l'on nomme la Pangenèse:
« les caractères acquis créaient dans
le corps des «gemmules» qui transmettaient ces
modifications dans les cellules
reproductrices ». (La
naissance du transformisme, Lamarck, entre Linné et
Darwin, Goulven LAURENT, Collection inflexions,
Vuibert/Adapt, 2001, p 131)
|
|
La sélection naturelle au sens de Darwin
semble avoir été équivalent de
"reproduction différentielle" (le terme de
"sélection" étant emprunté aux
éleveurs-sélectionneurs d'après
Ernst Mayr). Après tant
de débats, je ne suis pas sûr que chaque auteur
n'utilise pas ce mot dans des sens différents. Il me
paraît cependant être une
causalité donnant une direction à
l'évolution : le milieu (pression sélective du
milieu) sélectionne les individus les plus aptes (non
pas comme projet intelligent mais comme contrainte aveugle).
Certains scientifiques appliquent ce principe jusqu'aux
molécules dont l'organisation spatiale et
structurelle peut-être vue comme une
adapatation-sélection. La causalité est alors
diluée dans les mécanismes physico-chimiques
décrivant le réel. On est alors vraiment dans
le cadre d'une philosophie mécaniciste, ce qui n'a
plus grand chose à voir avec le darwinisme, qui est
une théorie du vivant qui peut s'accommoder d'un
vitalisme. Pour des détails voir ci-dessous, des
textes annexes sur la sélection
naturelle et surtout les textes de G. Canguilhem,
notamment le vivant et son milieu
qui pose à mon avis le vrai problème
philosophique: « Chez Darwin, on peut dire que
le finalisme est dans les mots (on lui
a assez reproché son terme de sélection) il
n'est pas dans les choses. Chez Lamarck, il y a moins
finalisme que vitalisme.»
|
|
deux nouveaux courants théoriques issus des
développement de la biologie du XXème
siècle, notamment des résultats de la
génétique
|
néolamarckisme
|
Simple référence au lamarckisme permettant
de proposer des interprétations qui ne cadrent pas
vraiment avec le darwinisme: notamment pour la
macroévolution.
|
|
néodarwinisme
|
Théorie évolutive du XXème
siècle issue du darwinisme et intégrant les
connaissances de génétique du XXème
siécle. L'espèce biologique est
définie comme une population interféconde
occupant une niche écologique; elle est en
quelque sorte réduit à un ensemble de
gènes qui s'échangent et se conservent au sein
de l'espèce. La génétique moderne s'est
ainsi nourrie du néodarwinisme mais en retour
celui-ci l'a intégrée en l'enfermant dans une
théorie actuellement fort contestée.
L'hérédité ne se limitant pas aux
gènes, il y a une spéciation non
génétique et pourtant basée sur ce qui
est héritable.
|
|
la thoérie la plus moderne, issue du
néodarwinisme
|
théorie
synthétique de l'évolution
|
Théorie évolutive
selon laquelle les mutations héréditaires se
font au hasard (évolution buissonnante, graduelle,
à vitesse constante) et sont triées
(modifications héréditaires présentant
un avantage reproductif conservées et
amplifiées, modifications héréditaires
diminuant l'efficacité reproductive
éliminées) par la sélection naturelle
(pression sélective du milieu)
|
|
paramètres, modèles et théories
partielles (ne prenant en compte qu'un aspect de
l'évolution)
|
spéciation
|
La spéciation désigne
les processus, modalités et
mécanismes de la formation des nouvelles
espèces. Pour beaucoup de biologistes
l'espèce est la seule classification naturelle des
êtres vivants. La spéciation est donc au
cœur de la compréhension de
l'évolution.
Pour les néodarwiniens la spéciation commence
par un isolement reproducteur qui permet
l'évolution de l'ensemble des gènes d'une
population (d'une espèce) sans échange avec
des populations voisines (de la même espèce) et
donc le maintien de la variabilité
génétique, cause de la variabilité des
espèces.
|
|
mutationnisme
|
Théorie imaginée par le botaniste Hugo De
Vries (1848-1935) selon laquelle les mutations sont le
matériau (moteur) de l'évolution, le passage
d'une espèce à l'autre se fait par mutation
brusque. Actuellement, du point de vue transformiste, on lui
préfère le terme de variabilité
génétique, qui ne fait pas implicitement
référence à une théorie
évolutive et qui dépasse les simples
mutations. On distingue ainsi deux types d'évolution
génétique:
|
|
deux aspects de l'évolution souvent
opposés, notamment au niveau
génétique
|
microévolution
|
Evolution entre individus et espèces
supposée se faire imperceptiblement.
Modèle évolutif selon lequel
l'évolution génétique est progressive
et lente (pratiquement continue) notamment par des mutations
au niveau des gènes de structure (qui sont
associés à des protéines intervenant
dans la structure des cellules et non dans leur
fonctionnement ou la régulation de leur
activité). C'est la partie la mieux documentée
de l'évolution génétique.
|
|
macroévolution
|
Evolution brusque ou lente mais majeure entre grands
groupes d'organismes.
Modèle évolutif selon lequel
l'évolution génétique est brusque et
rapide notamment par modifications de gènes de
régulation et de développement, par des
mécanismes chromosomiques comme des translocations,
duplications....c'est la partie la moins documentée
des modèles génétiques de
l'évolution (la plus hypothétique).
|
|
deux approches du rôle de la sélection
naturelle sur les produits des mutations
|
neutralisme
|
Théorie selon laquelle les mutations sont
"neutres" vis-à-vis de la sélection naturelle:
elles ne se traduisent ni par un avantage, ni par un
désavantage; la sélection naturelle ne fait
qu'éliminer les mutations les plus nocives; la
persistance ou l'élimination d'une modification
génétique est aléatoire.
L'environnement ne modifie pas la fréquence des
mutations
|
|
sélectionnisme
|
Théorie selon laquelle les mutations sont
sélectionnées par la sélection
naturelle de façon différente selon
l'efficacité reproductive qu'elles sont susceptibles
de conférer à l'individu: la sélection
est diversifiante, on parle aussi d'avantage à
l'hétérozygotie, car la nouveauté ne
peut provenir que des hétérozygotes, au moins
dans un premier temps. La variabilité est plus forte
dans un habitat instable qu'un habitat stable.
|
|
deux modèles pas forcément
incompatibles:
saltationnisme
et
gradualisme
|
Stephen Jay Gould, N. Eldredge,
S. M. Stanley, paléontologistes
américains, ont proposé dans les années
soixante-dix, un nouveau modèle d'évolution,
dit des «équilibres ponctués»
selon lequel l'apparition de nouvelles espèces est un
phénomène «rapide» (à
l'échelle des temps géologiques) et suivi de
longues périodes de stabilité des formes (ou
«stases») : on parle aussi d'évolution
discontinue et saltationniste (ou
encore buissonnante), par opposition à un
modèle continu et gradualiste.
Ce modèle permettait de réconcilier la macro
et la microévolution qui divisaient les
généticiens. Depuis la théorie
synthétique de l'évolution
préfère parler d'évolution
buissonnante.
|
d. Vers d'autres théories
d1. Dans la recherche des causes de l'évolution,
l'opposition n'est pas entre finalisme et mécanisme
mais plutôt entre vitalisme et mécanisme.
Pour une définition de ces mots en -isme et
un peu d'épistémologie voir le cours
des professeurs des écoles sur le
site associé.
On veut parfois opposer les finalistes (souvent qualifiés de
lamarckiens ou néo-lamarckiens) et les darwinistes (ou
néodarwiniens). Or je ne pense pas qu'il y ait des philosophes
pour affirmer que le darwinisme exclut le finalisme. Tout au
contraire. Le finalisme est, à mon avis, le passage
obligé de toute interprétation signifiante de
l'évolution.
Voici quelques extraits de la
conférence de G. Canguilhem sur le vivant
et son milieu qui traitent secondairement de l'opposition
lamarckisme-darwinisme mais qui se rapporte tout à fait
à notre problème :
« Lamarck écrit dans la Philosophie
zoologique (1809) que si, par action des circonstances ou action des
milieux, on entend une action directe du milieu extérieur sur
le vivant, on lui fait dire ce qu'il n'a pas voulu dire . (...) Selon
Lamarck, la situation du vivant dans le milieu est une situation que
l'on peut dire désolante, et désolée. La vie et
le milieu qui l'ignore sont deux séries
d'événements asynchrones. Le changement des
circonstances est initial, mais c'est le vivant lui-même qui a,
au fond, l'initiative de l'effort qu'il fait pour n'être pas
lâché par son milieu. L'adaptation c'est un effort
renouvelé de la vie pour continuer à « coller
» à un milieu indifférent. L'adaptation
étant l'effet d'un effort n'est donc pas une harmonie,
elle n'est pas une providence, elle est obtenue et elle n'est jamais
garantie. Le lamarckisme n'est pas un mécanisme; il serait
inexact de dire que c'est un finalisme. En réalité,
c'est un vitalisme nu. Il y a une
originalité de la vie dont le milieu ne rend pas compte, qu'il
ignore. Le milieu est ici, vraiment, extérieur au sens propre
du mot, il est étranger, il ne fait rien pour la vie. C'est
vraiment du vitalisme parce que c'est du dualisme. La vie, disait
Bichat, est l'ensemble des forces qui résistent à la
mort. Dans la conception de Lamarck la vie résiste
uniquement en se déformant pour se survivre.
(...)
Darwin se fait une tout autre idée de l'environnement
du vivant, et de l'apparition de nouvelles formes. Dans
l'introduction à l'Origine des Espèces, il écrit
: « Les naturalistes se réfèrent continuellement
aux conditions extérieures telles que le climat, la
nourriture, comme aux seules causes possibles de variations, ils
n'ont raison que dans un sens très limité. » Il
semble que Darwin ait regretté plus tard de n'avoir
attribué à l'action directe des forces physiques sur le
vivant qu'un rôle secondaire. Cela ressort de sa
correspondance. (...) Darwin cherche l'apparition des formes
nouvelles dans la conjonction de deux mécanismes : un
mécanisme de production des différences qui est la
variation, un mécanisme de réduction et de critique de
ces différences produites, qui est la concurrence vitale et la
sélection naturelle. Le rapport biologique fondamental,
aux yeux de Darwin, est un rapport de vivant à d'autres
vivants; il prime le rapport entre le vivant et le milieu,
conçu comme ensemble de forces physiques. Le premier
milieu dans lequel vit un organisme, c'est un entourage de
vivants qui sont pour lui des ennemis ou des alliés, des
proies ou des prédateurs. Entre les vivants
s'établissent des relations d'utilisation, de destruction, de
défense, Dans ce concours de forces, des variations
accidentelles d'ordre morphologique jouent comme avantages ou
désavantages. Or la variation, c'est-à-dire
l'apparition de petites différences morphologiques par
lesquelles un descendant ne ressemble pas exactement à ses
ascendants, relève d'un mécanisme complexe ; l'usage ou
le non-usage des organes (le facteur lamarckien ne concerne que les
adultes), les corrélations ou compensations de croissance
(pour les jeunes); ou bien l'action directe du milieu (sur les
germes).
En ce sens on peut donc dire que selon Darwin, contrairement à
Lamarck, l'initiative de la variation appartient quelquefois, mais
quelquefois seulement, au milieu. Selon qu'on majore ou minore
cette action; selon qu'on s'en tient à ses oeuvres classiques
ou au contraire, à l'ensemble de sa pensée telle que sa
correspondance la livre, on se fait de Darwin une idée un peu
différente, Quoiqu'il en soit, pour Darwin, vivre c'est
soumettre au jugement de l'ensemble des vivants une différence
individuelle. Ce jugement ne comporte que deux sanctions : ou mourir
ou bien faire à son tour, pour quelque temps, partie du jury.
Mais on est toujours, tant que l'on vit, juge et jugé. On
voit, par conséquent , que dans l'œuvre de Darwin, telle
qu'il nous l'a laissée, le fil qui relie la formation des
vivants au milieu physico-chimique peut paraître assez
ténu. Et le jour où une nouvelle explication de
l'évolution des espèces, le mutationisme, verra
dans la génétique l'explication des
phénomènes (que Darwin connaissait mais qu'il a
sous-estimés) d'apparition de variations spécifiques
d'emblée héréditaires, le rôle du
milieu se trouvera réduit à éliminer le pire
sans avoir part à la production de nouveaux êtres,
normalisés par leur adaptation non
préméditée à de nouvelles conditions
d'existence, la monstruosité devenant règle et
l'originalité banalité provisoire.
Dans la polémique qui a opposé lamarckiens et
darwiniens il est instructif de remarquer que les arguments et
objections sont à double sens et à double
entrée, que le finalisme est dénoncé et le
mécanisme célébré, tantôt chez
l'un, tantôt chez l'autre. C'est sans doute le signe que la
question est mal posée. Chez Darwin, on peut dire
que le finalisme est dans les mots (on lui a assez
reproché son terme de sélection) il n'est pas
dans les choses. Chez Lamarck, il y a moins finalisme que
vitalisme. L'un et l'autre sont d'authentiques
biologistes, à qui la vie paraît une donnée
qu'ils cherchent à caractériser sans trop se
préoccuper d'en rendre compte analytiquement. Ces deux
authentiques biologistes sont complémentaires. Lamarck pense
la vie selon la durée, et Darwin plutôt selon
l'interdépendance ; une forme vivante suppose une
pluralité d'autres formes avec lesquelles elle est en rapport.
La vision synoptique qui fait l'essentiel du génie de Darwin
fait défaut à Lamarck. Darwin s'apparente davantage aux
géographes, et on sait ce qu'il doit à ses voyages et
à ses explorations. Le milieu dans lequel Darwin se
représente la vie du vivant, c'est un milieu
biogéographique. »
On en arrive donc à un paradoxe pour le moins
étonnant : c'est le milieu physique de Lamarck, vitaliste, qui
est rejetté actuellement au profit du milieu vivant, et
pourtant mécaniste, darwinien, des biogéographes,
parfois poussé jusqu'au sens des anthropogéographes
d'organisme vivant (voir plus haut l'hypothèse de Gaïa ou
la géophysiologie).
Mais le débat est loin d'être clos car si l'on pousse
l'analyse il faut aller jusqu'à l'homme et à ses
limites de connaissance.
On peut encore citer G. Canguilhem, toujours dans la même
conférence sur le vivant et son
milieu « A partir de Galilée, et aussi de
Descartes, il faut choisir entre deux théories du milieu,
c'est-à-dire au fond de l'espace : un espace
centré, qualifié où le
milieu est un centre ; un espace
décentré, homogène,
où le milieu est un champ intermédiaire.» et
encore « Le milieu propre de l'homme c'est le monde de sa
perception (...). Mais l'homme, en tant que savant,
construit un univers de phénomènes et de lois qu'il
tient pour un univers absolu. La fonction essentielle de la science
est de dévaloriser les qualités des objets composant le
milieu propre; en se proposant comme théorie
générale d'un milieu réel, c'est-à-dire
inhumain. (...) En fait, en tant que milieu propre de
comportement et de vie, le milieu des valeurs sensibles et techniques
de l'homme n'a pas en soi plus de réalité que le milieu
propre du cloporte ou de la souris grise. » et il
conclut sa conférence avec une note d'espérance :
« Un sens, du point de vue biologique et psychologique, c'est
une appréciation de valeurs en rapport avec un besoin. Et un
besoin c'est pour qui l'éprouve et le vit un système de
référence irréductible et par là absolu.
»
Remarque :
Je viens de prendre connaissance de l'analyse "didactique" de la
notion de milieu que fait Jean-Pierre Astolfi dans le Que-sais-je sur
"La didactique des sciences" (P.U.F., 1996, p. 61-64): je
m'étonne d'y voir une progression, mais peut-être
suis-je complètement dans l'erreur. En tout cas pour moi, il
n'y a aucune progression dans ce type de représentation
(toutes les flèches non représentées entre tous
les milieux dégagés peuvent exister). Les positions
philosophiques me paraissent escamotées dans une
"flèche" de la connaissance (même si l'auteur semble se
décrier et affirmer qu'elle ne représente pas une
progression optimale, il n'en reste pas moins qu'il suppose une
progression...). Cette didactique repose certainement sur des
théories de la connaissance qui me sont
étrangères. En tout cas, le moins que je puisse dire
est qu'elles ne me convainquent pas. Autant le discours philosophique
de G. Canguilhem me semble passionnant, autant le discours
"didactique" de J-P. Astolfi me semble stérile (d'ailleurs,
cet auteur situe à C. Bernard le début de la
méthode expérimentale (p. 23 de l'ouvrage cité),
ce qui est pour le moins surprenant , ensuite il remet en cause la
notion même de science expérimentale et lui substitue
une épistémologie du savoir scientifique ; il est
à craindre que si la philosophie des sciences est
incontournable, la didactique des sciences ne soit l'entreprise la
plus stérile entreprise depuis longtemps (je sens que je vais
peut-être regretter cette phrase mais je ne résiste
pas... ) dans la mesure où elle repose sur une position
philosophique très particulière ; ce n'est pas de la
didactique des sciences mais une application d'une certaine
épistémologie à l'enseignement des sciences: il
est tout de même surprenant qu'en prenant un livre d'une
collection comme Que-sais-je ?, on ne puisse comprendre que des
bribes de la pensée des auteurs du fait de concepts qui,
d'après eux, sont tellement novateurs qu'ils en sont
totalement obscurs).
d2. Les critiques de Pierre-Paul Grassé
Je souhaite ici me faire l'écho de ces lignes qui,
même si elles ne permettent pas de clore le débat, ont
été souvent occultées et sont pourtant de vraies
questions intéressantes. Elles ne constituent pas une
théorie mais permettent son éclosion.
Pierre-Paul Grassé parle d'« amortissement
» de l'évolution (p 124) « ... la marge de
manoeuvre de l'évolution n'a cessé de s'amenuiser :
à l'Ordovicien, la genèse des embranchements
s'arrête, au Jurassique celle des classes, au
Paléocène-Eocène, celle des ordres.
»
*(p 130) « la filiation d'un type d'organisation à un
autre ne s'opère jamais par l'intermédiaire de types
spécialisés. La grande évolution s'est faite de
forme archaïque en forme archaïque qui, par leur plan
d'organisation, appartiennent bien à une unité
systématique définie, mais conservent une structure de
type général.... Elles sont les méres
d'où jaillissent les phyllums qui réalisent un certain
type morphologique ou idiomorphon, en s'abandonnant à la
spécialisation.... L'évolution créatrice prend
sa source dans les formes mères ; en leur absence, de nouveaux
types d'organisation n'apparaissent jamais.... »
*(133) « Bien des espèces n'ont pas
évolué depuis leur origine ou presque et sont les
survivants de groupes éteints » Voici quelques
espèces panchroniques ou reliques citées par
Pierre-Paul Grassé : Lingules (Brachiopodes de l'Ordovicien
connus encore actuellment dans nos mers), de nombreux
Gastéropodes, des Nautiles (aux formes très voisines
des actuelles dès le début du Primaire vers 320
millions d'années), des Odonates (libellules au sens
très large depuis le Carbonifère), le Cœlacanthus
(Poisson connu depuis le Carbonifère), le Sphénodon ou
Hatteria (un Reptileconnu depuis le Jurassique et encore
présent dans des petites îles de
Nouvelle-Zélande, ou encore les Oppossums ou Sarigues,
Mammifères connus depuis à quelques détails
osseux et dentaires près depuis le Crétacé
supérieur.
* certains groupes comme les Echinodermes (Ousins, Etoiles de mer,
Crinoïdes....) contiennent de très nombreux genres
actuels connus à l'état fossile. Certains sont connus
dès le Cambrien, ils ont eu des apogées à des
moments variés. Mais ils n'ont cessé d'évoluer
et n'ont perdu que 3 lignées sur les 8 connues.( p137-139).
Cette observation nous suggère que de nombreux groupes
inférieurs (Procaryotes, Protistes...) pour lesquels nous
n'avons pas de beaucoup de structures fossilisées certaines
(historiquement, paléontologiquement), étaient
probablement présents dans des temps très
reculés (on connaît, bien sûr, des Protistes
à tests, comme les Foramminifères ou les Radiolaires
connus depuis le précambrien...).
* La thèse de Pierre-Paul Grassé est que l'«
évolution actuelle se limite à la
spéciation » : c'est le cas par exemple de
quelques oiseaux isolés dans des îles comme les
Passériformes Drépaniidés des îles
Hawaï, les Géospizidés des Galapagos ("pinsons de
Darwin"), les Vangidés de Madagascar, les Pigeons de
Polynésie, ou d'Eponges, de Planaires ou de Crustacés
dans des lacs (lac Baïkal par exemple). Il cite comme exemple
majeur le colibacille (Escherichia coli) qui malgré ses
innombrables mutations conserve impeturbable ses
caractéristiques spécifiques d'une
génération à l'autre. Les mutations ne sont pas
un facteur d'évolution.
* « L'histoire se présente comme une suite
d'événements liés les uns aux autres par des
causes si nombreuses qu'on ne peut jamais être sûr de les
connaître toutes. ... En tant que faits historiques, tous les
phénomènes évolutifs se montrent
irréversibles. Et parmi eux, les genèses des grands
plans d'organisation sont des phénomènes
entièrement révolus » .
* Pierre-Paul Grassé cite le célèbre mot d'Henri
Poincaré : "Le hasard n'est que la mesure de notre
ignorance".Il précise ensuite combien il est faux de
parler de hasard quand on parle de combinaison des génotypes
parentaux dans un gamète ou un oeuf : la combinaison est une
loi de déterminisme (hasard probabiliste) et non un hasard par
lequel n'importe quoi pourrait arriver (accident dont les causes sont
si nombreuses qu'elles ne peuvent être connues et que l'on
appelle donc le hasard). (Chapitre IV p 157 et s.).
*Les études statistiques de génétique des
populations ne sont que de piètres modèles des
écosystèmes où jamais les individus ne sont
confinés comme le sont les drosophiles des élevages par
exemple.
* La théorie darwinienne de l'évolution
nécessite la notion de préadaptation dans le sens
où, le milieu changeant, un individu ne survivra dans ce
nouveau milieu que s'il est préadapté, afin que joue la
sélection naturelle. Cette « préadaptation
équivaut à une harmonie préétablie,
latente , que certains animaux, certaines plantes portent en eux...
»
*à propos de la sélection naturelle je vous engage
à lire le chapitre V que je ne saurais résumer, en
voici la conclusion presque complète : « Dans la
nature, la sélection exerce son action sur les espèces
en éliminant le "pire", les tarés, les infirmes...
C'est là son principal effet. La compétition
interspécifique joue un rôle dans la répartition
spatiale des populations et des espèces. Elle intervient dans
certaines évictions et limite le nombre des espèces
sympatriques. Il s'agit généralement
d'équilibres locaux ; l'issue de la lutte n'est point la
même partout; ici l'espèce est rejetée d'un
biotope, là elle est victorieuse et prospère. En
conséquence, les espèces sont d'autant plus
menacées dans leur existence, qu'elles habitent un territoire
plus restreint. Les faunes des îles de petite dimension sont
très sensibles aux introductions d'espèces
étrangères. Les Oiseaux aptères (Ralidés,
Columbiformes) qui prospéraient dans les archipels de
l'océan indien avant que l'homme les peuplât n'ont pas
tenu devant les prédateurs venus d'Europe. Mais l'homme a tant
massacré les espèces insulaires qu'il est difficile
d'apprécier la part qui, dans la destruction, revient aux
autres prédateurs. Les faits se rapportant de
près ou de loin à la sélection concernent bien
plus le peuplement de notre planète que l'évolution
biologique. Confondre les deux, c'est commettre une
erreur. La prétendue "évolution en action" de J. Huxley
et autres biologistes, n'est que la constatation de faits
démographiques, de fluctuations locales des génotypes,
des répartitions géographiques. Souvent il s'agit
d'espèces qui n'ont pratiquement pas changé depuis des
millions de siècles !... »
d3. Des critiques au sujet de la
sélection naturelle
Quelque soit l'origine de la variation, mutationisme, vitalisme,
préadaptation... (voir ci-dessus la citation de P-P.
Grassé et l'interprétation de Darwin que fait G.
Canguilhem et qui donne une plus ou moins grande importance au
milieu) on ne peut écarter la notion de sélection
naturelle. Elle est devenue pour certains une évidence.
Est-elle une constatation biologique, un corollaire à
l'adaptation au milieu (pris dans le sens biogéographique,
donc darwinien), ou bien une vision déformée,
malthusienne et anthropomorphique ? Pour éviter de donner ici
une interprétation uniquement personnelle je joins deux
textes de référence.
Pour moi la sélection naturelle n'est qu'une vision, à
mon avis fausse de la réalité. Elle n'est
qu'interprétation. Elle n'a pas de base métaphysique,
elle est opinion. Si l'on reste au niveau des causes
matérielles accessibles à la science la
sélection naturelle n'est qu'une formulation des contraintes
extérieures à l'organisme. Érigée en
principe causal, elle s'oppose à l'harmonie de
l'écosphère. Personnellement, j'ai toujours
préféré décrire les paramètres
extérieurs à l'organisme en terme de relations,
d'échanges plutôt que de contraintes
sélectives.
Dans le cadre d'une théorie de l'évolution qui
recherche les mécanismes évolutifs au niveau des
organismes en développement je précise un point pour
moi lumineux: l'évolution ne s'est pas faite entre
formes évoluées mais entre formes en cours de
développement (cellules sexuelles, gamètes,
zygotes, embryons...). (voir ci-dessous) La sélection
naturelle sur un individu adulte ou déjà bien
formé (variabilité de croissance, de morphologie, de
fécondité... due au milieu) est tout à fait
différente de celle que l'on peut imaginer peser sur une forme
embryonnaire. Présenter une contrainte sur un résultat
de l'évolution (le passsage d'un reptile coureur à un
reptile volant) est d'une façon évidente la description
d'une finalité.
d4. Une théorie de l'évolution qui est une
théorie globale du vivant par Rosine Chandebois
Un nouvel ouvrage de synthèse simplifiée est paru en
1999 : "Comment les cellules construisent l'animal "(voir
bibliographie). La théorie
évolutive de Mme Chandebois est cependant plutôt
traitée dans "Pour en finir avec le darwinisme: une
nouvelle logique du vivant", 1993. J'ai essayé
d'intégrer une petite partie des idées de Mme
Chandebois dans une page sur le travail du
développement à destination des enseignants dans le
cadre du nouveau programme de la classe de seconde des
lycées.
d5. Une autre théorie non darwinienne: celle
d'Anne Dambricourt sur l'apparition de l'homme
Deux références accessibles aux non
spécialistes; un article: Nouveau regard sur l'origine de
l'homme, Anne DAMBRICOURT-MALASSÉ, La Recherche,
286, avril 1996, 45-51 et un livre: "La légende maudite du
XXème siècle. L'erreur darwinienne" , La
Nuée Bleue/DNA, Strasbourg, 2000.
Je renvoie à quelques pages (que j'ai utilisées
ci-dessous) sur ses démêlés avec la critique des
scientifiques attachés au paradigme dominant darwinien:
http://uip.edu/critiques.html.
Les travaux d'Anne Dambricourt l'on conduit à deux
interprétations vraiment nouvelles puis à une
théorie.
* La première interprétation concerne le
développement de l'embryon. La bascule de la partie
postérieure du crâne est commandée par
l'enroulement du tube neural et non par la locomotion bipède
comme on l'a toujours cru (voir figure sur le site de l'uip:
http://uip.edu/images/ADMFigA.gif)
. Plus ce tube s'enroule, dans les premiers stades embryonnaires,
plus la bascule est importante. Le processus qui se répercute
sur les tissus règle aussi les rapports entre la face et la
base du crâne, c'est lui qui fait apparaître le petit
vide symphysaire que nous avons tous au milieu du menton et qui
n'existe chez aucun singe actuel ou passé. Il s'agit d'un fait
objectif que chacun peut vérifier, et qui est d'une grande
importance pour notre compréhension de l'apparition des
hominidés avec le cou basculé et le retrait du
prognathisme (la contraction qui fait que nous sommes les seuls
primates à avoir les dents sous le front).
* La deuxième interprétation concerne les
fossiles des crânes de la lignée humaine dont les
dimensions sont réinterprétées à l'aide
des outils utilisés par les spécialistes de
l'ondotologie actuelle. Les étapes de la lignée humaine
correspondraient à des "palier de contraction" (voir
la figure sur le site de l'uip: http://uip.edu/images/ADM
fig B copier.gif et l'article de la recherche :
Nouveau regard sur l'origine de l'homme, Anne
DAMBRICOURT-MALASSÉ, La Recherche, 286, avril 1996, 45-51). On
notera qu'ainsi l'homme fossile devient un genre à part
entière (genre fossile Sapiens), franchement
séparé des néandertaliens rattachés au
genre fossile Homo.
* La théorie proposée par Anne Dambricourt
reprend ses résultats sur la lignée humaine.
L'évolution dans cette lignée serait non graduelle. On
passe sans intermédiaire d'un palier de contraction
embryonnaire à un autre, chaque palier étant
défini par une embryogenèse fondamentale qui
résulte d'une refonte du plan d'organisation qui
intègre les fondements de l'ancien plan. La théorie se
fonde également sur le constat que la succession des plans
d'organisation évolue toujours de la même façon,
celle d'une "contraction cranio-faciale" avec un enroulement du tube
neural plus important. C'est le signe de l'existence d'un
phénomène qui ne doit rien au hasard des explications
darwiniennes classiques. En effet, on peut observer un processus,
qui, pendant 60 millions d'années et quelles que soient les
innombrables modifications de l'environnement que traversent les
espèces qui le portent, se répète comme induit
par un déterminisme interne. Il est également
remarquable que ce processus aille en s'accélérant,
l'arrivée d'un nouveau palier de contraction mettant toujours
moins de temps pour apparaître que celui dont il est issu.
4. Les phylogénies
Pour cette partie, l'évolution est
l'hypothèse de départ. Ainsi, désormais,
toute classification naturelle des être
vivants doit refléter leurs liens de parenté. Il
semblerait que déjà Darwin avançait que les
classifications deviendraient des
phylogénies. Le fait est
que, actuellement, le terme de phylogénie ou
phylogénèse (du grec phulon = race, et
génèse ou génie) est devenu
absolument équivalent à celui de classification
évolutive. Alors qu'à son origine
"phylogénie" se trouve dans le vocabulaire de Haeckel
(1874) et désigne l'évolution de la lignée d'un
organisme au cours des temps géologiques, qui est reprise dans
le processus d'ontogénèse, développement de
l'être vivant, comme nous allons le voir ci-dessous.
On peut cependant penser que l'on peut construire des classifications
non évolutives et intéressantes. Nous traiterons ce
point dans le prochain chapitre.
a. Les phylogénies ou classifications évolutives
classent... des caractères.
Nous distinguerons 2 types de caractères qui permettent de
construire des phylogénies, chacun apportant un
éclairage différent mais conduisant à des
phylogénies pas toujours conciliables :
- des caractères anatomiques, qui sont les plus
accessibles pour la paléontologie
- des caractères embryologiques qui permettent de
construire des phylogénies pour les seuls animaux
- des caractères moléculaires,
génétiques et chromosomiques qui permettent de
construire des phylogénies basées sur l'idée que
l'évolution biologique est la conséquence de
l'évolution génétique.
a1. Caractères anatomiques
Ce sont les caractères de l'anatomie comparée que
nous nous sommes efforcés de décrire rapidement dans
les méthodes de la
paléontologie puis que nous avons traité avec
l'exemple des Equidés
fossiles.
Au sujet de la signification évolutive des
caractères anatomiques il faut préciser la notion
d'homologie.
Deux structures homologues occupent une
même position anatomique et ont une origine embryonnaire
commune.
Cette définition semble actuellement discutée: je
renvoie à un article de Corinne Fortin dans l'APBG:
Classification et évolution, Biologie-Géologie
(Bulletin de l'APBG), n°3-2000, p 525-537. L'auteur propose un
aperçu historique du concept d'homologie et souligne que deux
structures homologues n'ont pas du tout forcément la
même fonction, ce que je croyais. (En effet, je pensais que le
premier critère de ressemblance était fonctionnel, ce
qui me permettait autrefois d'utiliser l'appellation "gènes
homologues" sans trop d'état d'âme). Elle y ajoute enfin
un critère évolutif: deux structures homologues
doivent être héritées d'un ancêtre
commun. Ce critère est justifié d'après
l'auteur par le fait que le terme d'homologie,
utilisé par Owen en 1843 pour décrire les relations
topologiques entre organes en dehors d'une interprétation
évolutive, vient en fait de l'analogie de
position proposée par Geoffroy Saint-Hilaire en 1818
dans sa Philosophie anatomique ("un organe est plutôt
altéré, atrophié, anéanti, que
transposé"). J'avais personnellement lu que le concept
d'homologie en biologie venait de J. W. von Goethe qui travaillait
sur le crâne de mouton. Il édifia ainsi, en 1820, la
«théorie vertébrale du crâne», selon
laquelle chacun des constituants du crâne est assimilable,
à une vertèbre. Cette thèse fut
développée par Lorenz Oken (1807), en Allemagne, par
Richard Owen, en Angleterre, et admise, malgré les
réserves de Cuvier, par la plupart des anatomistes de la
première moitié du XIXe siècle, Thomas
Huxley mit les choses au point en 1858, en montrant que le
développement du crâne se déroule de
manière semblable chez tous les Vertébrés et
sans trace de segmentation comparable à celle de la colonne
vertébrale (in Encyclopedia Universalis, article
"crâne"). Dans cette optique on serait bien dans le cadre d'une
hypothèse non évolutive a priori, contrairement
à ce qu'affirme Corinne Fortin. Je renvoie aussi à
l'article "analogie" de Pierre Delattre dans l'Encyclopédie
Universalis. L'homologie mathématique fait bien
référence à une ressemblance de fonction. Bref,
je crois avoir compris que Corinne Fortin ne souhaite pas prendre
parti sur un sens univoque d'homologie à utiliser en biologie.
Ce qui est fréquemment le cas de mots étudiés
avec une vision historique mais qui pose bien des problèmes
dans le cas de mots actuels. Il va donc être nécessaire
de changer de vocabulaire.
Pour en revenir à la question de départ :
l'évolution est-elle diversifiante ? la réponse que
l'on apporte est loin d'être définitive : si l'on
regarde les faunes ou les flores fossiles dans leur ensemble, il est
certain qu'on a une impression de diversité, dans les temps
géologiques, bien plus riche qu'à l'heure actuell. Mais
n'est-ce pas parce que l'on suppose involontairement que les
espèces fossiles s'ajoutent aux espèces connues? Qui
peut affirmer que les espèces au primaire étaient plus
nombreuse, plus diversifiées qu'actuellement ? Personnellement
je suis bien persuadé du contraire. Pierre-Paul Grassé
n'a certes pas été le seul à souligner combien
l'évolution n'a en fait créé que peu
d'embranchements et qui, d'après lui, ILS SONT TOUS ACTUELS.
Voici quelques mots extraits une fois encore de L'évolution du
vivant : «La genèse des plans fondamentaux
d'organisation, qui caractérisent les embranchements et les
classes, rameaux maîtres de l'arbre généalogique
du Règne animal, a été le grand oeuvre de
l'évolution. Les créations ont été peu
nombreuses : moins de vingt embranchements et quatre-vingt classes
pour le Règne animal (moins de la moitié pour le
Règne végétal). Elles sont de date très
ancienne. Le dernier des embranchements, celui des
Vertébrés, fit son apparition avec les Agnathes
(Ostracodermes, Cyclostomes) à l'Ordovicien, et avec les
Poissons au Dévonien, il y a quelques 450 millions
d'années. Depuis le Jurassique (Rhétien, 200 millions
d'années), époque des premiers Mammifères et des
précurseurs des Oiseaux (Portlandien, 135 millions
d'années), aucune nouvelle classe n'est apparue.»
Ou encore un peu plus loin (p 115) : «Chaque lignée,
au cours de son évolution, se comporte comme un
super-organisme qui obéit à son destin propre.
Plusieurs suivent un scénario qui comprend trois
périodes passant insensiblement de l'une à l'autre. A.-
Les pionniers de la lignée, ordinairement peu nombreux,
occupent une aire géographique restreinte. Leur anatomie se
spécialise peu mais décèle cependant des
particularités dont le nombre et l'ampleur augmentent avec
l'âge de la lignée. Leur évolution est lente,
voire très lente. C'est la période de jeunesse. B.-
Puis, à cette période d'installation, d'attente, de
tâtonnements, succède un temps de grande activité
; l'évolution s'accélère, se précipite :
les lignées se subdivisent, se différencient, se
spécialisent et poussent des rameaux latéraux. C'est la
période de maturité. C.- Enfin, les nouveautés
se raréfient, une relative stabilité succède
à la variation rapide. Des lignées meurent, d'autres
s'étiolent : c'est la période de sénescence ;
mais quelques-unes conservent leur vigueur et persistent
encore.»
Comment peut-on affirmer qu'aucun embranchement n'a disparu ? Pour un
non spécialiste, la reconstitution paléontologique et
le rapprochement de tel ou tel embranchement est loin d'être
quelquechose d'évident. Il y a là une sorte d'intuition
qui repose à mon avis sur la classification que l'homme fait
des espèces vivantes, ce que nous essaierons de
détailler un peu dans le chapitre suivant.
a2 . Caractères embryologiques
Si l'on essaye de se représenter l'évolution des
animaux au travail actuellement, on est tenté de penser
qu'elle doit se réaliser lors du développement
embryonnaire, dans ces étapes où le moindre
décalage dans le temps ou la moindre anomalie dans les
interactions entre populations cellulaires, semble pouvoir perturber
le développement et donc l'individu final obtenu. Si cette
idée peut facilement s'appliquer à la formation de
monstres ou d'embryons anormaux, il est cependant plus difficile
d'imaginer que ces différences puissent générer
de nouvelles structures.
Deux idées viennent cependant conforter le poids des arguments
embryologiques dans les phylogénies:
- Ernest Haeckel , la loi biogénétique
fondamentale et la sélection naturelle
Par une analogie lumineuse, Ernest Haeckel semble avoir
été le premier à comparer cette
évolution du vivant avec le déroulement dans le
temps d'un être vivant (mais il s'est limité aux
premières étapes de la vie !): c'est la "loi
biogénétique fondamentale" que l'on
résume souvent par "l'ontogénèse
récapitule la phylogenèse". On peut essayer
de détailler un peu les conditions dans lesquelles il a
énoncé cette loi.
Haeckel définit la loi de la différenciation,
ou loi de la division du travail comme une
conséquence de la sélection naturelle.
|
La loi de différenciation est
aussi appelée polymorphisme ou
divergence des caractères par Darwin :
tendance générale de tous les
êtres organisés à se
développer graduellement mais inégalement,
en s'écartant sans cesse du type primitif
commun. D'après Darwin, cette loi est
causée par le fait que plus deux individus se
ressemblent, plus la lutte entre eux est intense !!!
Haeckel développe ensuite son idée et
reprend sa théorie selon laquelle
«l'idée d'espèce n'est pas plus
absolue que les idées de variété, de
famille, d'ordre, de classe, etc. »Il finit en
affirmant : «les variétés sont
des espèces qui commencent. De la
variabilité ou la faculté d'adaptation des
espèces résultent nécessairement,
sous l'influence de la lutte pour l'existence, la
différenciation toujours croissante des
variétés et la perpétuelle
divergence des formes nouvelles.... L'origine de
nouvelles espèces par la division du travail, la
divergence ou différenciation des
variétés, résulte
nécessairement de la sélection
naturelle.» La deuxième
conséquence de la sélection naturelle est
la loi de progrès ou de
perfectionnement (teleosis)
selon laquelle on constate qu'à toutes les
époques de la vie organique sur la terre, il y a
eu progression dans le degré de perfection des
êtres organisés.
|
|
« Les deux séries de
développement organique, l'ontogenèse de
l'individu et la phylogenèse du groupe, auquel il
appartient, sont étiologiquement liées de
la façon la plus intime. ... Comme je l'ai dit
alors (Morphologie générale, II, 110-147,
371), l'ontogenèse ou
l'évolution individuelle, est une courte
et rapide récapitulation de la
phylogenèse, ou du développement
du groupe correspondant, c'est-à-dire de la
chaîne ancestrale de l'individu, et cette
ontogenèse s'effectue conformément aux lois
de l'hérédité et de
l'adaptation» . Ces deux derniers termes faisant
référence à la sélection
naturelle : «action combinée,
générale de l'hérédité
et de la variabilité dans la lutte pour
l'existence ».
|
|
« Au fond de tous les
organismes il y a une communauté originelle ; au
contraire, la différence des formes provient des
rapport nécessaires avec le milieu
extérieur ; il faut donc admettre une
diversité originelle simultanée et une
métamorphose incessamment progressive, si l'on
veut comprendre les phénomènes constants et
les phénomènes variables »
(Goethe, La métamorphose des animaux, 1819,
cité dans ibid, p 79). Il y a une
« puissance formatrice intérieure de
l'hérédité, qui maintient
l'unité du type » (puissance de
spécification), et la « puissance
formatrice externe de l'adaptation, d'où
dérive la variété des formes
organiques » (ibid p 80).
|
Peut-on séparer dans la pensée d'Haeckel (même
si cela est sans aucun doute contraire à sa conviction
clairement exprimée) ce qui est théorie darwinienne
et l'idée selon laquelle les premières étapes
de la vie d'un être vivant sont le reflet de son
évolution ?
Et dans ce cas, quelle validité donner à cette
idée ?
Pierre-Paul Grassé, quant à lui, affirme (p 58):
« Ne lui en demandons pas trop, car dans un grand nombre
de cas l'ontogenèse est profondément modifiée
par des cénogénèses et des adaptations
à des modes de vie très particuliers ; nous pensons
aux larves pélagiques ou simplement planctoniques dont les
caractères, prétendus archaïques, ne le sont
peut-être pas ».
- les hétérochronies
Une théorie récente va dans le même sens. Et
même si je manque encore une fois de temps pour
détailler cette idée, il me semble important de
mentionner les hétérochronies. On peut en
donner la définition suivante : décalages dans le
temps des agencements spatiaux et temporels des
éléments de l'organisme lors de
l'ontogenèse. Pour ce que j'en sais, c'est notamment
Stephen Jay Gould qui fût l'un des instigateurs (1977,
Ontogeny and phylogeny, Harvard Univ. Press ed., Cambridge, 501
p... je n'ai pas encore lu ce livre ; on peut aussi certainement
citer deux auteurs importants sur cette question : Mc Kinney M.L.
et Mc Namara K.J., 1991, Heterochrony : the evolution of ontogeny,
Plenum Press ed., New York, 437 p). Cette théorie a donc
été présentée d'emblée dans la
perspective darwinienne mais je ne pense pas que cela soit la
seule possible.
Si je ne me trompe cette idée est issue
d'interprétations réalisées sur des fossiles
cherchant à expliquer des modifications subites des formes
fossiles par un mécanisme ontogénétique.
L'idée est sans aucun doute excellente et je pense que l'on
peut voir l'hétérochronie comme un retour de la
vision évolutive de l'ontogenèse par Haeckel
(l'ontogenèse récapitule la phylogenèse),
appliquée aux fossiles et qui revient au vivant. On se
trouve bien en face d'une théorie ontogénique,
expérimentale donc, issue (par induction puis
déduction) d'une théorie évolutive globale.
Cette idée peut très bien être utilisée
dans une perspective darwinienne (d'où elle est issue), que
dans une perspective différente, puisqu'elle est devenue
expérimentale. Il est à noter cependant que si de
nombreuses études paléontologiques ont
été faites avec le modèle des
hétérochronies, très peu d'études sur
le vivant ont cherché à démontrer la
validité du concept. Il est certain que si le modèle
des hétérochronies reste au niveau historique, il
n'apporte pas grand chose. Il faut absolument le tester sur le
vivant (voir page complémentaire -
octobre 2003).
Remarque :
Il faut préciser que si l'on pousse un peu la comparaison
entre ontogenèse et phylogenèse (développement
de l'être vivant dans le temps et évolution des
êtres vivants au cours de l'histoire), on en arrive
effectivement à considérer que l'avenir du monde vivant
est la mort car, une fois le stade adulte atteint, le seul devenir de
l'être vivant est la sénescence et la mort.
On ne peut donc pas pousser la comparaison trop loin sans tomber dans
la plus pure fiction, mais il y a tout de même une
séduction dans l'idée d'Haeckel dont on aimerait bien
trouver la cause.
A mon sens, elle vient de la considération que je vous
proposais plus haut sur l'évolution comme résultat du
travail du vivant. Ce travail est recommencé à chaque
être vivant qui se forme. Les mécanismes à
l'œuvre dans l'évolution sont visibles, accessibles lors
de cette embryogenèse.
J'en arriverai donc un peu plus loin donc à vous
présenter une théorie du vivant qui a été
proposée par une embryologiste expérimentale : Rosine
Chandebois, professeur à l'université de
Provence-Côte d'Azur.
a3. Caractères moléculaires,
génétiques et chromosomiques
La présentation ci-dessous est historiquement fausse, mais
elle est un choix pédagogique du fait de la culture
générale de la population scolaire.
a31. les ressemblances entre deux molécules
sont chiffrées par la
distance
génétique.
|
La distance génétique entre deux
molécules homologues (qui ont la
même fonction et la même origine, voir
ci-dessus)
estime le nombre minimal de différences de leurs
séquences en unités de distance
génétique.
Par exemple deux protéines dont la séquence
primaire (chaîne des acides aminés)
diffère de deux acides aminés, seront à
une distance génétique de 2 unités.
Deux gènes associés à la
synthèse de deux molécules quasi-identiques et
fonctionnelles mais différent de trois
nucléotides seront à une distance
génétique de 3 unités.
|
On construit alors des phylogénies par la méthode des
moindres distances génétiques. On place des organismes
actuels à l'extrêmité de rameaux dont la longeur
des branches est proportionnelle à la distance
génétique entre chacun des organismes pris deux
à deux. Les nœuds de l'arbre correspondant à des
ancêtres communs et étant placés de façon
logique afin de minimiser les écarts entre l'arbre et les
distances mesurées.
Exemple de construction de phylogénies
à partir de la comparaison de la séquence des
chaînes A et B de l'insuline de 4
vertébrés
(la myxine (M): un Agnathe, vertébré sans
mâchoire apparenté aux lamproies; le cobaye
(C); Mammifère rongueur; le boeuf (B):
Mammifère ruminant; et l'homme (H): Mammifère
Primate). (Bordas, p 342)
Remarque: la méthode
employée ici n'est qu'une illustration et n'est pas
utilisée par les
systématiciens...
|
|
arbre construit à partir de
l'évaluation des distances génétiques
entre les chaînes A
|
arbre construit à partir de l'évaluation
des distances génétiques entre les
chaînes B
|
arbre construit à partir de l'évaluation
des distances génétiques entre les
chaînes A et B
|
|
|
M
|
C
|
B
|
H
|
|
M
|
-
|
9
|
6
|
6
|
|
C
|
9
|
-
|
9
|
8
|
|
B
|
6
|
9
|
-
|
2
|
|
H
|
6
|
8
|
2
|
-
|
|
|
|
M
|
C
|
B
|
H
|
|
M
|
-
|
17
|
14
|
14
|
|
C
|
17
|
-
|
10
|
10
|
|
B
|
14
|
10
|
-
|
1
|
|
H
|
14
|
10
|
1
|
-
|
|
|
|
M
|
C
|
B
|
H
|
|
M
|
-
|
26
|
20
|
20
|
|
C
|
26
|
-
|
19
|
18
|
|
B
|
20
|
19
|
-
|
3
|
|
H
|
20
|
18
|
3
|
-
|
|
|
H
|
|
B
|
M
|
C
|
|
1\
|
/1
|
|
/1
|
/1
|
|
|
+
2\
|
/2
|
/
|
2
|
|
|
|
+
1,5\
|
/1,5
|
|
|
|
|
|
+
\
|
|
La distance entre l'H et le B est de 2
unités, donc la distance moyenne, supposée
identique est de1 unité entre un ancêtre commun
et chaque organisme;
La distance moyenne entre le groupe H et B et la M est de
(6+6)/2 = 6 ; en supposant la distance identique entre
chaque organisme actuel et l'ancêtre commun, on place
la M à 3 unités de l'ancêtre commun, ce
qui oblige à placer à 2 unités l'un de
l'autre les 2 ancêtres communs.
Pour C la distance est environ de 9 unités de chacun
des autres organismes, ce qui fait 9-3 =6 unités de
l'ancêtre commun de ceux-ci.
Dans cet arbre, à but strictement
pédagogique, le cobaye se serait séparé
avant les agnathes de la lignée des
vertébrés, ce qui n'a aucun sens
paléontologique.
|
|
H
|
|
B
|
C
|
M
|
|
0,5\
|
/0,5
|
|
/0,5
|
/0,5
|
|
|
+
4,5\
|
/4,5
|
/
|
4,5
|
|
|
|
+
5 \
|
/5
|
|
|
|
|
|
+
\
|
|
La distance entre l'H et le B est de 1
unités, donc la distance moyenne, supposée
identique est de 0,5 unité entre un ancêtre
commun et chaque organisme;
la distance moyenne entre le groupe H et B au C est de
(10+10)/2 = 10, donc si l'on tient compte des 0,5 entre
l'ancêtre commun de l'homme et du bœuf, il reste
une distance de 9,5 que l'on sciende en 4,5 et 5 afin de
placer les organismes actuels à la même
duistance de l'ancêtre commun le plus ancien. De la
même manière la M est à 14 unités
de l'H et du B et à 17 du C donc une distance moyenne
de (14+14+17)/3=15 unités que l'on sépare en
10 et 5 unités comme
précédemment.
Dans cet arbre encore à but strictement
pédagogique les agnathes se séparent bien des
gnathostomes (vertébrés à
mâchoire) avant les rongeurs et les mammifères
puis les primates.
|
|
H
|
|
B
|
C
|
M
|
|
1,5\
|
/1,5
|
|
/1,5
|
/1,5
|
|
|
+
7,8\
|
/7,8
|
/
|
7,8
|
|
|
|
+
1,8 \
|
/1,8
|
|
|
|
|
|
+
\
|
|
La distance entre l'H et le B est de 3
unités, donc la distance moyenne, supposée
identique est de 1,5 unité entre un ancêtre
commun et chaque organisme;
la distance moyenne entre le groupe H et B au C est de
(19+18)/2 = 18,5, que l'on scinde en (18,5-1,5*2)/2=7,8 et
(18,5-1,5*2)/2+1,5=9,2; la distance moyenne de la M avec les
trois autres organismes est de (26+20+20)/3 =22 et
scindé de la même manière en 9,2, 3,6 et
11.
Dans cet arbre toujours à but strictement
pédagogique les agnathes se séparent bien des
gnathostomes (vertébrés à
mâchoire) avant les rongeurs et les mammifères
puis les primates.
|
|
Aucun de ces arbres phylétiques n'est VRAI,
historiquement, il traduisent chacun l'idée de
filiation (posée comme une hypothèse) en
fonction du degré de ressemblance entre deux
chaînes d'acides aminés d'une de leur
protéine hormonale chez des organismes actuels. En ce
sens ce sont des ILLUSTRATIONS de la ressemblance
moléculaire (phénotypique)
regardée dans une optique évolutive.
|
Si l'on souhaite utiliser ces arbres comme arguments d'un
modèle évolutif on fait de l'évolution
moléculaire phénotypique. Cette méthode
repose sur des postulats autrement plus hypothétiques
que la paléontologie classique. En voici un essai de
délimitation (en plus du principe de l'actualisme):
* limites : les molécules chimiques évoluent
et sont le reflet de l'évolution des formes des êtres
vivants : si l'évolution des êtres vivants est une
interprétation soutenue par d'innombrables arguments
paléontologiques, l'évolution des molécules
n'a aucun argument paléontologique ; elle est une
hypothèse dans le cadre de l'interprétation
évolutive ; elle n'apporte aucun argument à
l'évolution. Donc les conclusions obtenus ne sont pas des
arguments évolutifs se sont des arguments d'évolution
moléculaire. Étant bâtie sur une
hypothèse, ses conclusions ne peuvent dépasser cette
hypothèse. Dans l'exemple étudié ci-dessus,
le fait de retrouver des molécules de même fonction et
de séquence voisines dans différents groupes de
vertébrés ne suggère pas l'évolution
mais, dans le cadre d'une interprétation évolutive, on
peut les expliquer en les faisant dériver les unes des autres.
Leur filiation établie n'est en aucun cas un argument
évolutif puisqu'il est posé au départ. Le fait
de retrouver deux gènes identiques codant pour deux produits
identiques chez deux espèces actuelles ne donne aucun argument
évolutif. Mais dans le cadre d'une interprétation
évolutive, on peut les interpréter comme issues par
exemple d'un gène unique ancestral. Mais si l'on veut utiliser
cette interprétation comme argument d'évolution - et
donc dire que ces deux espèces ont donc probablement un
ancêtre unique -, on tombe dans le sophisme: la conclusion est
contenue dans la prémisse.
Par contre on peut utiliser le critère de ressemblance mais
dans ce cas, non pas comme argument évolutif mais comme
argument pour classer les espèces. Les arguments
moléculaires sont des critères de classification mais
non évolutifs. Une classification évolutive repose elle
aussi sur la même hypothèse et l'on risque de tomber
dans le même sophisme que précédemment.
L'évolution posée comme hypothèse, on classe les
espèces avec des critères uniquement
considérés dans le cadre de cette hypothèse. La
valeur de ces classifications ne dépasse pas les limites de
l'hypothèse de départ.
* spécificité : les molécules
chimiques se ressemblent plus entre espèces qu'elles ne
différent entre individus ou chez un même individu : on
peut dire qu'il existe une signature moléculaire de
l'identité de l'individu, de l'espèce et des groupes
d'organismes d'ordre supérieur. Les espèces
actuelles reflètent, dans leur composition moléculaire,
les similitudes entre individus d'une même espèce, d'une
même classe, d'un même embranchement (on gomme les
variations phénotypiques individuelles ou on suppose qu'on
peut les isoler).
* stabilité : les molécules utilisées
n'ont pas évolué dans les groupes
considérés (espèce et donc population,
embranchement) et les différences observées
actuellement sont les traces des différences
hypothétiques passées. La seule évolution
qui est acceptée est celle, discontinue donc, entre les
groupes. Il ne peut être considérée
d'évolution individuelle ou intraspécifique. Bien
entendu, l'évolution individuelle au cours de la vie de chaque
individu ne peut pas non plus être prise en compte.
Cela fait beaucoup d'hypothèses pour justifier autrement
que par un phénomène de mode et d'entraînement
politique et financier, l'importance donnée aux
phylogénies moléculaires. Cette méthode est
intéressante mais délicate et chère, et doit
être toujours présentée avec ses postulats car
ils ne sont pas simples à mettre en évidence car ils
sont souvent implicites.
Remarque 1:
il est très contestable de comparer des séquences d'ADN
(empreintes génétiques par exemple) sans faire
référence à des molécules associés
aux gènes car la majorité du génome des
eucaryotes est constitué d'ADN repété qui n'est
pas utilisé par la cellule et qui semble de pas avoir de sens
d'ADN pour des molécules (information
génétique).
Remarque 2:
la notion d'horloge moléculaire est indissociable de la
théorie neutraliste des mutations (Kimura, 1990 ; voir
par exemple L'évolution, des faits aux
mécanismes, Louis Allano et Alex Clamens, ellipses, 2000,
p 52-60) et est très discutable.
Le taux de substitutions d'acides aminés par site (position
d'un acide aminé dans la chaîne) et par année est
assez significativement constant . Une famille de
protéines homologues peut ainsi servir d'horloge
moléculaire de l'évolution: connaissant le nombre de
substitutions de la protéine d'un organisme par rapport
à une organisme supposé ancestral, on eut
déterminer son âge. Il faut cependant de nombreux et
importantes hypothèses pour arriver à ce
résultat:
* on doit d'abord ramener chaque différence d'acide
aminés au nombre minimal de mutations ponctuelles par
substitution nécessaire pour produire chaque changement
d'acide aminé (du fait des autres types de mutations et des
redondances du code génétique, cette méthode
n'est pas sans nouvelles approximations et hypothèses);
* on doit ensuite caler dans le temps les phylogénies
ainsi obtenues à l'aide des données
paléontologiques: le fossile le plus ancien permettant
d'approcher au mieux le nœud qualifié "d'ancêtre
commun" dans les phylogénies précédentes (ce qui
empêche toute utilisation paléontologique de l'horloge
réalisée).
L'exemple classiquement donné (Bordas, p 353) est celui de la
famille des globines. La critique principale, en plus des
hypothèses soulignées plus haut, est que le taux de
substitutions par site est constant par année et non par
génération, ce qui n'a aucune signification
biologique étant donné que les mutations ne semblent
pouvoir progresser dans les populations que par la reproduction.
a32. la comparaison des génomes associés
à la synthèse de molécules homologues chez
des organismes apparentés permet de retracer une
histoire (dans le cadre de l'hypothèse
évolutive)
L'exemple le plus fréquemment utilisé est celui des
hémoglobines dans la série des vertébrés.
(Bordas p 354-355) En voici un exposé simplifié dans un
but de mémorisation.
|
Les données de la génétique moderne
permettent d'imaginer l'action de deux types de
mécanismes principaux: les duplications de
gènes et les transpositions de gènes. Ces deux
mécanismes peremttant d'obtenir des groupes de
gènes homologues situés sur différents
chromosomes et formant ce que l'on appelle une famille
multigénique (de molécules homologues). Pour
les chaînes de la globine (partie protéique de
la molécule d'hémoglobine) on distingue deux
groupes: les chaînes du groupe alpha (alpha, tseta...)
et les chaînes non alpha (beta, epsilon, delta...).
Les gènes associés aux chaînes du groupe
alpha sont portés par le chromosome 16 et les
gènes associés aux chaînes du groupe non
alpha par le chromosome 11.
|
|
Chez les Oiseaux et les Mammifères, les
deux groupes de gènes sont portés par deux
chromosomes différents, ce qui pourrait être la
conséquence de duplications faisant suite à
uen transposition.
|
groupe non alpha
chromosome 11
/\
groupe alpha
chromosome 16
/\
|
DUPLICATIONS
|
|
étape intermédiaire ne
correspondant pas à un génome
existant
|
/\
|
TRANSPOSITION
(= translocation)
|
|
Chez certains Amphibiens deux gènes sont
portés par le même chromosome, ce qui peut
s'interpréter comme la conséquence d'une
duplication génique.
|
/\
|
DUPLICATION
|
|
Chez les Agnathes, un seul gène est
associé à 4 chaînes identiques. On
qualifie de gène d'ancestral.
|
gène ancestral unique
|
|
exercice Bordas, n°7, p 368
a33. la comparaison des caryotypes chez des organismes
apparentés permet de proposer des hypothèses
concernant la spéciation (formation de nouvelles
espèces) par isolement reproducteur
L'exmple le plus classique est celui de l'homme et du
chimpanzé (Bordas, p 382).
Pour la technique de coloration des chromosomes et d'obtention du
caryotype, voir cours de
spécialité. Je rappelle sommairement que les bandes
ne correspondent pas du tout à des unités
fonctionnelles (gènes...) et que l'on ignore la raison du
très faible polymorphisme de cette coloration au sein d'une
espèce (bandes quasi-spécifique du génome d'une
espèce). Cette technique permet ainsi de faire des comparaison
entre des chromosomes de taille et de répartitions semblables
de bandes et donc de mettre en évidence :
* 13 chromosomes identiques (sur une formule chromosomique de 22
+X(X,Y) pour l'homme et 23 + X(X, Y) chez le chimpanzé)
* un chromosome n°2 de l'homme qui pourrait résulter de
la fusion de 2 chromosomes du chimpanzé ou inversement 2
chromosomes n°2 du chimpanzé issus de la
séparation du chromosome n°2 de l'homme
* 9 possibles inversions péricentriques (simples ou doubles)
sur les chromosomes 2, 4, 5, 9, 12, 15, 17, 18
* 1 possible addition de matériel chromosomique, sur le
chromosome 1 humain ou une déletion sur ce même
chromosome chez le chimpanzé
* 2 possibles délétions de matériel
chromosomique sur les chromosomes 13 et 18 humains ou des additions
sur les mêmes chromosomes du chimpanzé
* des modifications encore plus complexes sur les chromosomes 9 et
15.
Quelle est l'interprétation que l'on peut faire
actuellement de ces ressemblances et de ces
différences ?
* on connaît des mécanismes chromosomiques
actuels fins qui pourraient rendre compte d'une telle
évolution génomique.
* les ressemblances sont toujours interprétées
en terme de parenté: plus le génome de deux
espèces est proche plus on considère que ces
espèces ont une origine voisine. Cet argument repose sur le
principe d'unité du monde vivant appliqué au nombre,
à la forme, la taille et les bandes de coloration des
chromosomes.
MAIS parenté ne veut pas dire "ancêtre
commun". La parenté peut se faire entre groupes voisins,
même si, à l'origine, un (ou deux) des membres du groupe
a du s'isoler (reproductivement) et donner naissance à un
autre groupe. La notion d'ancêtre commun fait
référence à des phylogénies non cladistes
alors que la cladistique refuse cette notion (voir plus
bas).
*ACTUELLEMENT les différences sont
- principalement interprétées en terme
d'accidents ou d'anomalies. Les anomalies sur le nombre de
chromosomes (trisomies, monosomies...) sont toujours gravement
invalidantes et l'on estime que la plupart des trisomies (sauf 21,
18, 17...) sont non viables.
- parfois interprétées en termes de modifications
"neutres", sans modification de la fonctionnalité du
génome Les translocations sans ajout ou perte de
matériel (dites équilibrées), par exemple, sont
considérées comme neutres, même si elles sont
rarement dépistées.
- enfin, certaines modifications sont interprétées
comme responsables de la mise en place d'une
barrière reproductrice d'ordre
génétique. Les duplications de génome,
naturelles ou provoquées sont considérées comme
donnant naissance à des individus d'une autre espèce.
Un cas souvent rapporté est celui de l'espèce
Myosotis alpestris , qui se répartit en fait 2
espèces: l'une à 24 chromosomes et l'autre à 48
chromosomes, espèces qui ne s'hybrident pas (pour donner des
individus à 36 chromosomes). Ce mécanisme est
considéré comme important dans la spéciation
par isolement reproducteur. Cependant, dans le cas de
reproduction sexuée, il faut imaginer un COUPLE d'individus
atteints tous deux simultanément des mêmes modifications
FIXÉES HEREDITAIREMENT (c'est à dire non
réparées par la cellule) pour donner naissance à
une descendance fertile.
Les cas de reproduction sexuée entre individus de 2
espèces différentes et donnant naissance à des
hybrides de première génération stériles
est aussi considéré comme la preuve d'une
barrière reproductrice. On pense que les deux lots
chromosomiques n'ont pas d'homologie suffisante pour qu'il y ait
appariement (par exemple, de la stérilité du mulet,
hybride entre l'âne et le cheval). Mais cette notion de
barrière génétique n'a pas vraiment de base
expérimentale cytologique et l'explication pourrait
être d'un tout autre ordre (physiologique, cytologique...).
On verra dans la partie d4 que l'on a par contre des
données écologiques, comportementales, physiologiques
et moléculaires de plus en plus précises sur les
mécanismes de spéciation (par exemple pour la Souris,
voir L'Évolution, des faits aux mécanismes, p
31-33).
Donc, il ne s'agit pas de faire dériver l'homme d'un
chimpanzé double monosomique au caryotype profondémment
altéré. Il semble plutôt que ces modifications
chromosomiques puissent être la trace de
phénomènes cellulaires qu'il nous reste à mettre
en lumière. C'est pourquoi on se contente souvent
d'utiliser ces ressemblances chromosomiques comme des
caractères génétiques sans les
considérer comme un mécanisme réel de
macroévolution. L'exemple d'un arbre
phylogénétique ainsi obtenu et limité à
une seule paire de chromosome est donné dans le Bordas p 383,
n°4. (celui du chromosome n°7 extrait de
Dutrillaux et présenté dans L'évolution des
faits aux mécanismes, p 51 (fig. 2-10), est autrement plus
parlant, mais peut-être est-ce le même, simplifié
?).
En conclusion
Les comparaisons moléculaires de séquences de
protéines réalisées sur le vivant (et
éventuellement sur des restes fossilisés avec une forte
incertitude sur leur âge) ne permettent que de parler de
ressemblances chimiques ou moléculaires, entre deux individus
ou plus, si les analyses ont été
réalisées sur plusieurs spécimens. L'extension
à l'espèce est souvent non justifiée.
L'extension dans le temps complètement
hypothétique.
Les comparaisons sur l'ADN, molécule instable, doivent
être encore plus prudentes. La séquence d'un gène
d'un individu n'a jamais permis d'établir des
caractères d'espèce, tout au plus la capacité
à la synthèse éventuelle d'une molécule.
Comment peut-on continuer de présenter avec la même
valeur un caractère anatomique, lié à par
exemple une articulation crânio-dentaire de plusieurs fossiles
et un séquençage de l'ADN de quelques spécimens
appartenant à deux groupes distincts ? Les arguments ne sont
pas du même ordre.
Je me garde bien aussi d'utiliser les termes "séquence
homologues" ou pire encore "gène homologue". Voir à ce
sujet la discussion sur l'homologie plus haut
dans cette page.
b. Les phylogénies cladistes
(d'après la méthode de Willi Hennig) classent des
caractères polarisés
(cette partie est clairement hors programme
mais incontournable: en effet il me semble que le cladisme fait
encore partie de ces théories cachées volontairement
dans les programmes : il est fréquent d'entendre dire que
seules les cladogrammes sont de véritables classifications
évolutives : c'est totalement faux (on trouvera une telle
affirmation, par exemple, dans le livre de Guillaume Lecointre et
Hervé Le Guyader: Classification
phylogénétique du vivant, Belin, 2000 et bien
sûr tous les textes repris de celui-ci, des documents
d'accompagnements du nouveau programme de TS à certains
nouveaux manuels de cette classe). D'autre part, étant
donné la difficulté de revenir aux hypothèses et
aux théories philosophiques supposées lors de la mise
en oeuvre de ces phylogénies, le travail de mise à
niveau de l'enseignant est à la limite de l'acceptable;
d'où l'intérêt de travailler avec des philosophes
qui nous aideraient à voir les non-dits de ces
théories. Tout ceci pour dire que je ne maîtrise pas la
question et que ce que j'en ai compris n'est certainement que
très partiel.)
Il y a au moins deux lectures du cladisme :
- la première chronologiquement semble celle que l'on
retrouve dans Comprendre et enseigner la planète
terre, Ophrys, 1989 ou dans le Dictionnaire de
géologie, A. Foucault et J.F. Raoult, Masson, 1992,
dont voici les définitions :
clade : nom masculin [du grec klados, rameau] -
Taxon monophylétique, c'est-à-dire unité
systématique comprenant exclusivement une espèce
ancestrale prise comme référence et toutes les
espèces qui en descendent.
cladistique : adjectif [du grec klados,
rameau] - Qui se rapporte aux clades. Analyse
cladistique : recherche des caractères des êtres
vivants permettant de mettre en évidence leur appartenance
à un clade. Systématique cladistique :
classification des êtres vivants basée sur la
filiation des espèces (plutôt que sur de simples
ressemblances).
cladogenèse : nom féminin [du grec
klados, rameau et de genèse] - Apparition de
lignées divergentes au cours de l'évolution à
partir d'une même espèce ancestrale. Antinomie :
anagenèse.
[anagenèse : nom féminin [du grec
ana, vers le haut et de genèse ] - Evolution
continue d'une lignée d'êtres vivants par
transformation progressive de divers caractères, sans
création de lignées divergentes. Antinomie :
clagogenèse.adjectif :
anagénétique.]
cladogramme : nom masculin [du grec klados,
rameau et gramma, écrit] - Représentation
graphique des relations de parenté entre taxons.
La cladogenèse est considérée comme une
hypothèse d'interprétation évolutive qui
s'oppose à l'anagenèse. Il me semble que cette
acception n'est plus utilisée en 2002.
- la seconde lecture est ce que l'on peut appeller
l'école universitaire française cladiste avec des
références accessibles facilement pour un enseignant
du secondaire comme par exemple : La construction de
phylogénies, Guillaume Lecointre,
Biologie-Géologie (Bulletin de l'APBG), n°1-1995, pp
109-136, ou l'article de Pascal Tassy : Un arbre pas comme les
autres, La Recherche, 296, mars 1997, pp 76-81.
Le cladisme est une nouvelle méthode de construction
d'arbres phylogenétiques: elle se pose en unique et seule
vraie méthode scientifique.
Pour une approche voir une page sur le cladisme.
Il me semble important que dans plusieurs ouvrages récents
(notamment "Le livre de la vie", p 221) il soit affirmé que
- au moins pour l'homme, mais c'est une idée que l'on
retrouve pour d'autres phylogénies - on ne peut pas
représenter d'arbre. Si la cause principale en est due
à la présence d'énormes lacunes dans nos
archives fossiles, il est aussi suggéré que il n'y a
aucune preuve de relation entre les lignées : ce n'est pas
un précurseur-ancêtre qui donne par adaptation un
individu plus évolué. Chaque étape
évolutive, marquée par une espèce fossile
correspond à une population adaptée à un
milieu donné à une époque donnée. La
notion de primitif conduisant à l'évolué est
souvent refusée pour laisser la place à celle de
lignées présentant des filiations. On pourrait
croire que cette idée est nouvelle, mais il me semble que
ce n'est qu'une conséquence du cladisme. Une des
idées centrales du cladisme étant en effet de
considérer les groupes frères à partir de
parentés établies pour chaque caractère, il
n'y a donc pas un ancêtre mais bien des groupes parents.
L'ancêtre n'existe pas pour un cladiste (ce qui n'est
pas très bien exposé par nombre de manuels
scolaires...). En s'inspirant de l'article de Claudine Cohen
(Nos ancêtres dans les arbres, La Recherche,
345, septembre 2001, p 33-37) on pourrait ainsi dire que
«certes, toute espèce, actuelle et fossile, à
des ancêtres », mais, il est impossible du retrouver
parmi tous les fossiles cet ancêtre
précisément, du fait du phénomène
d'évolution. Cette vision relativiste de la
paléontologie, sous prétexte de ne s'appuyer que sur
des critères scientifiquement déterminés,
refuse à l'évolution biologique le statut de
déterminisme.
Pour une approche de la construction de cladogrammes voir le
manuel de terminale S de Nathan, 1994, p326-327.
Ou le manuel de1ES : Bordas, 2001, p 290-291 et bien sûr,
depuis le nouveau programme de TS en 2002, les nouveaux manuels de
cette classe.
c. On peut faire des classifications évolutives qui ne
soient pas des cladogrammes
Actuellement trois écoles se partagent le devant de la
scène:
- la phénétique : qui ne prend en compte
que les caractères visibles d'un organisme
(phénotype), méthode qui remonte au
XVIIIe siècle avec le botaniste Michel Adanson. La
phénétique recherche le plus grand nombre de
caractères possibles (c'est pour cela qu'on parle parfois
de "classification naturelle"). Une classification
phénétique se traduit sous la forme d'un graphe
branchu, un dendrogramme (ou
phénogramme), sans racine, qui
traduit les degrés de ressemblance entre organismes et non
une généalogie. La hiérarchisation des
caractères repose sur l'anatomie et l'embryologie et seule
l'unité du monde vivant est prise comme hypothèse de
départ.
- ces deux dernières méthodes sont historiques ou
généalogiques
(phylogénétiques) et reposent sur
l'établissement de groupes apparentés.
- la cladistique construit des
cladogrammes qui traduisent graphiquement
les clades ou lignées évolutives
construites à partir de caractères
polarisés (dont on décide s'ils sont primitifs ou
évolués à l'aide d'hypothèses
évolutives paléontologiques et embryologiques et
donc ainsi, à mon avis, relativisées
(voir paragraphe b, ci-dessus))
- la systématique évolutive construit
des phylogrammes qui traduisent des
parentés déduites principalement à partir
d'homologies (ressemblance entre deux structures qui ont
la même position anatomique et ont la même origine
embryonnaire).
Un article intéressant mais qui appelle de
nombreux commentaires : De la phylogénie... sans bouillir
ou les bases de la reconstitution phylogénétique,
Bruno Chanet, Biologie-Géologie (Bulletin de l'APBG)
n°3-1997, pp 505-527; L'essentiel de l'article est une
présentation de la méthode cladistique (avec une
brève explication de la méthode
phénétique, toujours à partir de l'exemple de P.
Tassy de l'article de La Recherche sur l'Evolution cité plus
haut). Mais une idée me semble à souligner : "on
peut établir des phylogénies sans fossiles". Je
suis tout à fait d'accord mais dans ce cas, on ne fait pas de
la paléontologie, on ne sort pas de l'essai de classification
et sans fossile, il n'y a pas d'évolution. Faire une
reconstitution cladistique (et donc utiliser la paléontologie
et la loi biogénétique fondamentale pour polariser les
caractères) et dire que l'on utilise pas les fossiles est une
tromperie. On utilise des théories qui n'ont de sens que dans
une certaine interprétation évolutive
réalisée à partir des fossiles. Une affirmation
comme "toute espèce actuelle est le résultat d'une
histoire, d'une évolution..." n'est pas simple : dans une
phrase comme celle-ci le terme d'«espèce» prend deux
significations successives et différentes :
l'«espèce biologique» actuelle (sujet du
verbe "est") et l'«espèce
paléontologique» qui est population isochrone fossile
(espèce historique en évolution). De la même
manière un caractère ne peut pas être
défini de la même façon chez un être vivant
et un fossile. Quand à dire que «seuls les
caractères déterminés
génétiquement ("sous contrôle
génétique") sont utilisables», c'est aller
un peu vite (voir le chapitre de génétique
et la liaison gène-caractère...), car le moins qu'on
puisse dire c'est que, à part le déterminisme de
quelques caractères moléculaires de quelques
espèces, les gènes ont un pouvoir de contrôle
qu'il reste à établir.
Pour montrer l'étonnante actualité
des présentations du XIXème, voici un arbre
phylétique des vertébrés tiré de
l'ouvrage d'Haeckel de 1874 : Histoire de la création des
êtres organisés d'après les lois naturelles
par Ernest Haeckel, professeur de zoologie à
l'université de Iéna, C. Reinwald et Cie,
libraires-éditeurs à Paris, consultable à la
bibliothèque du Muséum d'Histoire Naturelle d'Angers :
contrairement à ce qu'affirme certains auteurs, les
critères d'établissement de cet arbre sont tout
à faits transparents : ils font un chapitre complet du livre
et je ne peux donc pas les recopier ici.
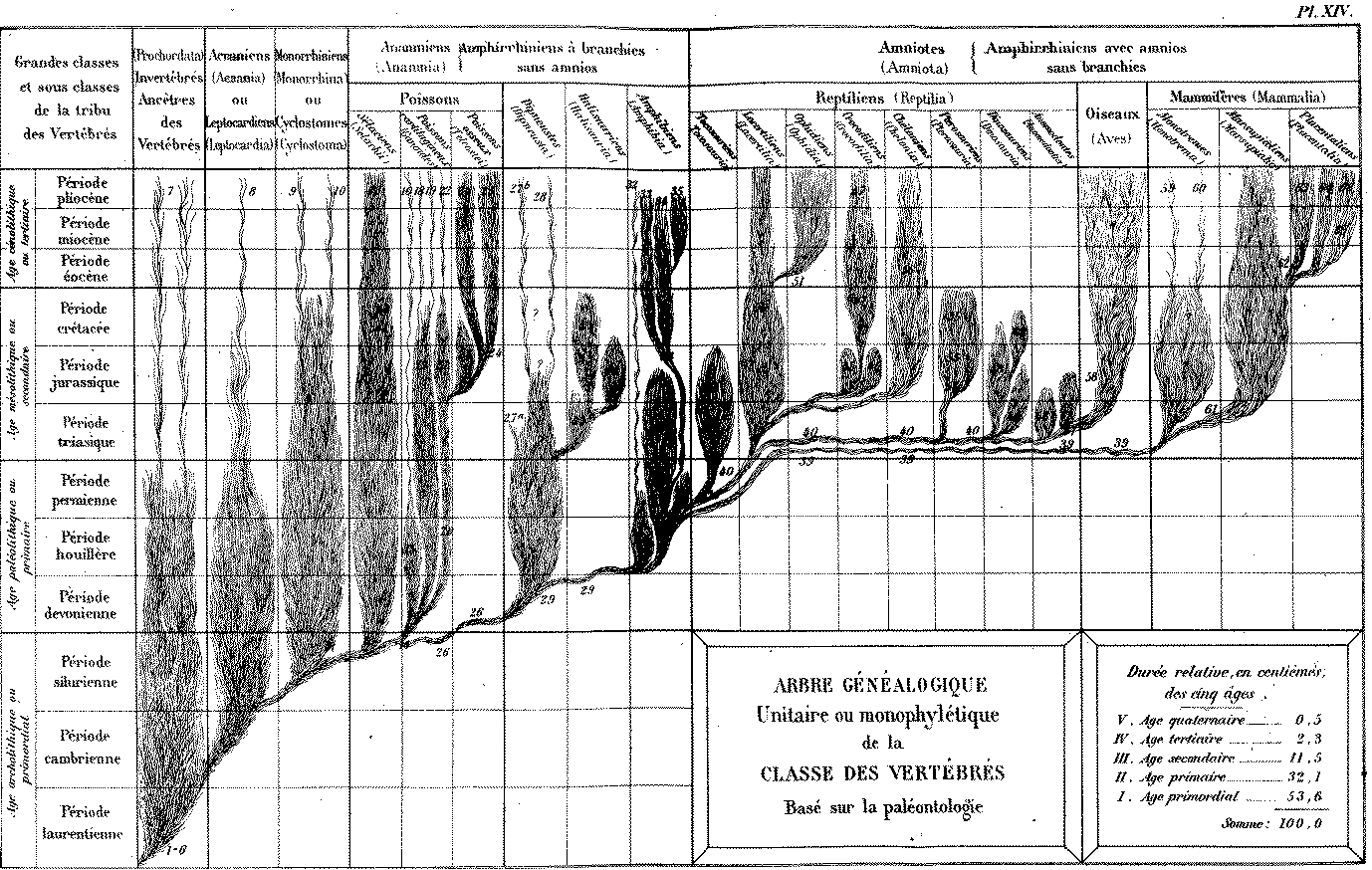
(image lisible, et utilisable avec des élèves
avec les légendes ci-contre, si elle est ouverte dans
une fenêtre séparée ou imprimée
au format paysage à 85%...)
|
voici la légende de cette Planche
XIV:
«Arbre généalogique
unitaire ou monophylétique de la tribu des
vertébrés, figurant l'hypothèse de
la commune origine de tous les vertébrés et
l'évolution historique de leurs différentes
classes durant les périodes géologiques. Les
lignes horizontales indiquent les périodes de
l'histoire organique terrestre, durant lesquelles se sont
développées les couches fossilifères.
Les lignes verticales séparent les unes des autres
les classes et sous-classes de
vertébrés.
|
|
Les lignes ramifiées indiquent par
leur nombre plus ou moins grand et d'une manière
approximative le plus ou moins grand degré
d'évolution, de diversité et de perfection que
chaque classe a du atteindre dans chaque période
géologique. Quant aux classes qui, à cause du
peu de consistance de leur corps, n'ont pu laisser
derrière elle de débris fossiles (par exemple
les procordates, les acrâniens, les monorrhiniens et
les dipneustes), le cours hypothétique de leur
évolution a été indiqué
d'après les données fournies par les trois
sources de documents sur la création organique, par
l'anatomie comparée, l'ontogénie et la
paléontologie. Pour combler hypothétiquement
les lacunes paléontologiques, on s'est surtout
appuyé, ici comme partout, sur la loi
biogénétique fondamentale, reposant sur
l'intime union étiologique entre l'ontogénie
et la phylogénie (voir les planches VIII et XIII, et
p 273 et 359). Partout on doit considérer
l'évolution individuelle comme une courte et rapide
récapitulation de l'évolution
paléontologique. Cette récapitulation a pour
cause fondamentale les lois de
l'hérédité ; mais elle est
modifiée par celle de l'adaptation. Cette proposition
est le «Ceterum senso» de notre théorie de
l'évolution.
Les données relatives à l'apparition
première, à l'époque originelle de
chaque classe et sous-classe des vertébrés,
telles qu'elles sont indiquées sur la planche XIV,
sont, autant que possible, strictement déduites des
faits paléontologiques, en exceptant, bien entendu,
les compléments hypothétiques indiqués.
Néanmoins, je dois faire remarquer que ,
vraisemblablement, l'origine de la plupart des groupes est
antérieure d'une ou de plusieurs périodes au
moment indiqué par nos fossiles actuels. A ce sujet,
je partage l'opinion d'Huxley ; cependant, autant que
possible, je me suis attaché, sur les planches V et
XIV, à ne pas trop m'écarter des faits
paléontologiques.
|
|
Voici la signification des chiffres.
(Voir la XXe leçon).
1. Monères animales. 2, Amibes animales. 3,
Synamibes. 4, Planæa. 5, Gastræa. 6,
Turbellaria. 7, Tunicata. 8, Amphioxus. 9, Myxinoida. 10,
Petromyzontia. 11, Formes transitoires inconnues entre les
monorrhiniens et les poissons primitifs. 12, Poissons
primitifs siluriens (Onchus, etc.). 13, Poissons
primitifs actuels (requin, chimère, raie). 14,
Poissons cartilagineux très anciens (Silurien
Pterapsis). 15, Pamphracti. 16, Sturiones. 17,
Rhombiferi. 18, Lepidosteus. 19, Polypterus. 20,
Cœloscopes. 21, Pycnoscolopes. 22, Amia. 23,
Thrissopida. 24, Poissons osseux avec conduit
aérifère de la vessie natatoire (Physostomi).
25, Poissons osseux sans conduit aérifère de
la vessie natatoire (Physoclysti). 26, Types
intermédiaires inconnus entre les poissons primitifs
et les dipneustes. 27, Ceratodus. 27a, Ceratodus
éteint du trias. 28, Dipneustes d'Afrique
(Protopterus). 29, Formes intermédiaires
inconnues entre les poissons primitifs et les amphibies. 30,
Ganocephala. 31, Labyrinthodonta. 32, Cæciliæ.
33, Sozobranchia. 34, Sozura. 35, Anura. 36,
Proterosaurus.37, Formes inconnues intermédiaires aux
amphibies et aux protammiens. 38, Protammiens (forme
ancestrale commune à tous les amniotes). 39,
Promammalia. 40, Proreptilia. 41, Thecodontia. 42,
Simosauria. 43, Plesiosauria. 44, Ichtyosauria. 45,
Amphicœla. 46, Opisthocœela. 47, Prosthocœla.
48, Dinosauriens carnivores, Harpagosauria. 49, Dinosauriens
herbivores, Therosauria. 50, Mososauria. 51, Forme
ancestrale commune des serpents (Ophidiæ). 52,
Cynodontia. 53, Cryptodontia. 54, Rhamphorynchi. 55,
Pterodactylii. 56, Chersita. 57, Tocornithes. Formes
intermédiaires aux reptiles et aux oiseaux. 58,
Archæopteryx. 59, Ornithorynchus. 60, Echidna. 61,
Formes intermédiaires inconnues entre les
monotrèmes et les marsupiaux. 62, Formes
intermédiaires inconnues entre les marsupiaux et les
placentaires. 63, Villiplacentaires. 64, Zonoplacentaires.
65, Discoplacentaires. 66, Homo pithecogenes, improprement
appelé par Linné homo
sapiens.»
|
Je manque de temps pour proposer une utilisation
pédagogique de cet arbre et je ne crois pas que mes
élèves y soient sensibles mais il serait à mon
avis très intéressant de faire une comparaison avec un
arbre moderne (par exemple celui proposé dans Comprendre et
enseigner la planète terre, Ophrys, 1989, p. 248). Montrer
quels sont les groupes nouveaux ? Quels sont les critères
retenus actuellement ? etc. On peut bien sûr le comparer
à un cladogramme comme celui présenté dans La
construction de phylogénies, Guillaume Lecointre,
Biologie-Géologie (Bulletin de l'APBG), n°1-1995, p 110
et légendes p 126-128.
Dernière question : l'évolution
est-elle complexifiante ?
Pierre-Paul Grassé répond affirmativement et il utilise
cette observation pour justifier de la concordance entre les
classifications du monde vivant et les phylogénies :
l'évolution va du simple au complexe, du primitif à
l'évolué.
Je ne reprends pas ici la discussion cladistique (voir plus haut)
mais il faut signaler que les cladistes présentent
actuellement leur méthode comme la seule capable de
présenter une classification évolutive, mais le
raisonnement est trompeur et je ne suis pas sûr d'arriver
à exprimer clairement mon opinion, il faudra peut-être
plus de temps pour que tout cela apparaisse au grand jour. Ce
à quoi arrivent les cladistes, ce sont des phylogénies
à partir de n'importe quel caractère observé
dans des espèces actuelles et polarisés
(qualifiés de primitif ou dérivé grâce
à des hypothèses issues de la paléontologie et
de la loi biogénétique fondamentale) et donc sans faire
référence directement à une théorie
évolutive. On est si loin de la théorie
évolutive qu'on est en droit de se demander s'il y en a une.
Dans ce cas, dire que l'on propose la seule interprétation
évolutive sans faire référence à une
théorie évolutive relève de
l'artifice.
En fait je me demande si, finalement, ce qui justifie le
mélanges des fossiles et des espèces actuelles, n'est
pas seulement l'idée qu'il n'y a qu'UNE histoire des
êtres vivants, une seule phylogénie, ordonnée,
hiérarchisée. Si l'évolution n'était
qu'un possible alors comment en retrouver ses mécanismes dans
l'ontogenèse et ses traces dans les fossiles ?
Pour être déchiffrée, l'évolution doit
être un déterminisme, une causalité
expérimentale. L'est-elle ? Nous essaierons de proposer
des réponses dans le chapitre suivant.
Je recommande aussi fortement la lecture de l'article de Claudine
Cohen: Nos ancêtres dans les arbres (La Recherche, 345,
septembre 2001, p 33-37) qui démonte bien des
présupposés masqués lors de la construction des
arbres généalogiques humains et qui recommande
d'attendre prudemment d'en savoir plus pour oser en construire.
d. L'espèce est à la base de toute classification
ou dénombrement du monde vivant
Pour les néodarwiniens comme pour de nombreux
transformistes actuels l'évolution du vivant c'est
l'évolution des espèces ("L'origine des
espèces" est le titre de l'ouvrage majeur de Darwin mais
il ne contient pas de définition de l'espèce). La
définition de l'espèce est donc à la base de
toute classification évolutive ou toute approche de la
diversité biologique car celle-ci est évaluée en
nombre d'espèces.
Mais une espèce reste un concept biologique lié
à la reproduction sexuée, qui est loin de toucher tous
les règnes (les bactéries se reproduisent par division
binaire et la reproduction asexuée est extrêmement
répandue chez les unicellulaires ou les plantes) . La
taxonomie (science de la classification des êtres vivants
ou taxinomie du grec "taxis = rangement " et
"nomos = loi - correct") continue actuellement de nommer
tout être vivant par deux noms: un nom de Genre (avec
une majuscule) et un nom d'espèce (avec une minuscule),
selon l'usage hérité de Linné (Escherichia
coli, Homo sapiens...). Ce naturaliste suédois, dans son
Systema naturae , dont la dixième édition,
publiée en 1758, fonde la classification des plantes (1753)
exclusivement sur un nombre limité de caractères
sélectionnés (ceux liés à la
reproduction: les étamines et le pistil). Pour la
première fois, il codifie sous une forme condensée la
description des plantes et des animaux et introduit la
nomenclature binominale: les espèces y sont
désignées par un nom double, en latin, véritable
nom propre à chaque espèce. On a gardé cette
dénomination binôminale mais il est évident que
les critères de classification ont changé (voir
ci-dessus, paragraphe c). Seuls les plantes et les animaux peuvent
être nommés avec une certaine
homogénéité mais très souvent un nouveau
nom d'espèce est donné sur des critères
morphologiques en attente de confirmations et d'observations
ultérieures de la biologie de l'organisme (on utilise parfois
le terme de parataxinomie pour désigner cette
classification provisoire).
d1. le problème philosophique de l'espèce
Voici 2 formulations d'un même problème vraiment
philosophique et non pas seulement logique:
* l'espèce est-elle une classification artificielle,
élaborée par l'homme ou est-elle réellement une
entité du monde vivant ?
* est-ce l'homme qui ordonne, qui range la nature ou la nature
est-elle ordonnée indépendamment de l'esprit humain
?
Personnellement je suis certain de
l'ordre de la nature et que l'espèce est une entité qui
est le reflet d'une réalité (voir les pages sur la
science).
Il y a un ordre dans la nature qui dépasse l'homme mais qui en
quelquesorte le sert. La nature est ordonnée à l'homme
dans le sens où elle le révèle. Elle est le lieu
et l'occasion de la découverte et de l'expression de son
humanité. Elle n'est pas soumise mais révèle
l'humanité de l'homme. La classification que fait l'homme de
la nature est à la fois catégorisation
extérieure (nommer) et manifestation de ce qui est
caché. L'homme est à la fois partie de la nature (en
profonde harmonie avec cette vie dont il participe) et à la
fois responsable d'elle (du fait que l'homme expérimente un
pouvoir qu'il a sur la nature qui dépasse celui de tous
les autres êtres vivants). Cette vision est certes
inséparable de ma foi chrétienne.
d2. l'espèce vivante (biologique)
Voici deux définitions de l'espèce biologique
(et donc actuelle) :
* « rassemblement d'individus morphologiquement plus ou moins
semblables entre eux, ayant un stock commun de gènes et un
même genre de vie. Très généralement, ils
sont interféconds et leur progéniture est
également féconde.» (Cuénot, 1951)
* « groupe de populations naturelles capables potentiellement
d'intercroisements, mais qui sont reproductivement isolées
d'autres groupes semblables » (Mayr, 1963) ; ou sa version
modifiée : « une espèce est une
communauté reproductive de populations (reproductivement
isolée d'autres communautés) qui occupe une niche
particulière dans la nature » (Mayr, 1989,
cité dans l'article "A propos de la notion
d'espèce", Louis Allano et Alex Clamens,
Biologie-Géologie (Bulletion de l'APBG), n°3-1996,
471-472 ); la notion de niche écologique la plus
utilisée actuellement est celle de Hutchinson (1957) :
«ensemble des conditions dans lesquelles vit et se
perpétue la population».
Depuis cet article ces auteurs ont publié un petit ouvrage
dense à recommander (L'évolution, des faits aux
mécanismes, Louis Allano et Alex Clamens, Ellipses,
2000). En voici quelques éléments
particulièrement clairs «les espèces sont les
véritables catégories du Vivant, les autres taxons
n'ont qu'une utilité pratique... l'espèce est donc le
produit de l'évolution biologique....la spéciation,
devient le thème majeur de l'étude de
l'évolution biologique». De plus j'y ai trouvé
la suite du raisonnement sur la niche écologique que
j'ignorais: en effet si une espèce occupe une niche
écologique et une seule, chaque niche en retour n'a qu'une
seule espèce d'un taxon donné (p 35).
Je suis par contre beaucoup moins convaincu par les
conséquences de l'isolement reproductif qu'ils trouvent dans
l'espèce : pour eux le déterminisme des
caractères de l'espèce étant
génétique, et la reproduction étant
elle-même une recombinaison de gènes, l'isolement
reproductif est un isolement génétique (ils parlent
d'une «définition génétique de
l'espèce»). C'est ainsi que pour eux, le
problème de la spéciation devient le problème de
l'isolement reproducteur, première étape de
l'évolution permettant la conquête d'une nouvelle niche
écologique. Aussi, d'une façon générale,
dans le premier chapitre, il me semble facile de remplacer
"génétique" par "héréditaire" et tous
leurs exemples sont très pertinents. D'autre part, ils
justifient le caractère naturel de l'espèce comme taxon
du Vivant en faisant remarquer que les noms scientifiques modernes
des espèces animales et végétales se superposent
quasiement aux noms traditionnels donnés par des ethnies
anciennes. Mais cette superposition n'est à mon avis que le
reflet de la classification anthropomorphique des espèces,
naturelle à l'homme. Ce qui n'empêche pas son
caractère naturel, bien au contraire, car c'est bien l'homme
qui classe et pour moi il n'y a qu'une nature humaine.
Une remarque personnelle :
si l'espèce n'est pas un concept typologique (voir ci-dessus
ma position philosophique) et n'ignore pas le polymorphisme des
individus, la catégorie espèce présente des
limites qui peuvent être fluctuantes du fait de nos
connaissances. Je signale aussi que l'espèce, subdivision du
genre, est étymologiquement rapportée à la forme
: en latin species signifie "ce qui se voit avec les yeux"...
J'ai retrouvé le texte d'une conférence
de Mayr au Collège de France en 1978 qui éclaire sa
position philosophique sur l'évolution et qui la remplace dans
l'histoire des sciences avec ce qu'il appelle la "pensée
populationnelle". C'est assez intéressant, même si je ne
conçois pas du tout la réalité comme lui. On
peut aussi rapporter la conception de Lamarck de l'espèce,
variable, même si elle représentait pour lui une
réalité certaine : «un ensemble» ou
«une collection d'individus semblables ou presque
semblables», «qui furent produits par d'autres
individus pareils à eux»(La
naissance du transformisme, Lamarck, entre Linné et
Darwin, Goulven LAURENT, Collection inflexions, Vuibert/Adapt,
2001, p 19).
d3. l'espèce morte (paléontologique)
Une espèce paléontologique
est définie comme la forme vivante ayant conduit
aux fossiles classés dans une population isochrone.
(ex: Bordas p 310-311: les Ammonites)
La notion d'espèce paléontologique ne peut
que s'appuyer strictement sur des caractères morphologiques ou
au moins déduits des données morphologiques (voir plus
haut la notion de "fait
paléontologique"), ou d'espèce isochrone
lorsque les caractéristiques morphologiques des individus
supposés d'âge identique (population isochrone) et
regroupés dans une même espèce se
répartissent graphiquement selon une courbe approximativement
gaussienne.
Il y a malheureusement d'innombrables espèces
paléontologiques définies simultanément par
plusieurs paléontologues sous des noms différents,
certaines formes étant souvent rapprochées en
considérées comme deux sexes d'une même
espèce, d'autres formes sont au contraire scindées en
plusieurs espèces distinctes... bref, les critères
morphologiques n'étant ni universellement définis ni
indépendants d'autres découvertes, il est certain que
toute arbre présentant la diversité des espèces
fossiles, tout essai de dénombrement des espèces
fossiles, reste une présentation personnelle de chaque auteur,
sans qu'il puisse y avoir le consensus d'une science
expérimentale. De la même façon, le rattachement
de telle espèce à tel ou tel embranchement ou grand
groupe actuel est toujours une question d'opinion, les arguments
n'étant jamais définitifs.
Que l'on ne se méprenne pas, une fois encore, je ne veux pas
dire que l'histoire est inaccessible, mais qu'elle n'est pas une
science expérimentale.
Il est probable qu'il sera bientôt nécessaire de faire
un tri dans toutes ces espèces et genres fossiles, essayer de
simplifier comme l'avait déjà fait Lamarck, en
établissant des filiations entre espèces successives ou
dérivées et en réunissant des espèces
séparées en autant de variétés d'une
même espèce pour ne conserver que des différences
majeures. Ceci ne pourra se faire que si l'on s'accorde à une
définition unifiée, évolutive de
l'espèce.
d4. la spéciation: vers une définition
évolutive de l'espèce
Exemple: Bordas p 360: Larus fuscus et Larus
argentatus... un exemple pédagogique de E. Mayr à
l'intérêt historique, dont la critique est
particulièrement soignée par les deux
spécialistes d'ornithologie: Louis Allano et Alex Clamens dans
leur ouvrage: L'évolution, des faits aux
mécanismes (Ellipses, 2000), p 25 et suivantes.
Un exemple beaucoup mieux documenté et plus moderne est celui
de la souris domestique (même ouvrage, p 31-33).
Dans les livres d'écologie "moderne" cités en
référence, l'unité d'étude des
systèmes vivants n'est pas l'espèce mais la
population (ou plutôt le système
population-environnement). La population étant
définie comme un ensemble d'individus de la même
espèce. Les limites géographiques
n'étant pas fixées, on peut étudier la
population mondiale d'une espèce de criquet ou bien la
population de cette espèce sur l'île de Madagascar. Il
est évident que l'on retombe en fait sur la notion
d'espèce.
Mais le fait principal est que la population est une
sous-catégorie de l'espèce et donc que dans une
population d'individus de la même espèce les individus
n'ont pas forcément l'intégralité du
génome de l'espèce.
En paléontologie, le raisonnement est inversé, c'est
ce que l'on considère comme une population (population
statistique ou population isochrone), qui nous permet de
définir une espèce isochrone ou
paléontologique.
Lamarck, fondateur de la paléontologie des
Invertébrés, n'a as éludé le
problème de l'espèce et il donne notamment cette
définition: « Ainsi, parmi les corps vivans, la
nature, comme je l'ai déjà dit, ne nous offre, d'une
manière absolue, que des individus qui se succèdent les
uns aux autres par la génération, et qui proviennent
les uns des autres; mais les espèces, parmi eux, n'ont qu'une
constance relative, et ne sont inavariables que temporairement.
Néanmoins, pour faciliter l'atude et la connaissance de tant
de corps différens, il est utile de donner le nom
d'espèce à toute collection d'individus semblables,
que la génération perpétue dans le même
état, tant que les circonstances de leur situation ne changent
pas assez pour faire varier leurs habitudes, leur caractère et
leur forme.» (Philosophie zoologique, t 1, 1809,
P. 74-75, in La naissance du transformisme, G. Laurent,
Vuibert/Adapt, col. inflexions, 2001, p 96) Lamarck donne ici une
véritable définition évolutive de
l'espèce qui est à la fois une espèce
paléontologique par la référence à la
collections d'individus semblables (que l'on appelle maintenant
population) et une espèce biologique par
référence à la génération, mais
aussi enfin une espèce changeante de par les circonstances de
leur situation (ce qui permet d'envisager à la fois des
facteurs internes que des facteurs externes à ces
changements). Lamarck ne me semble pas avoir été
dépassé.
Le but actuel est toujours d'unir ces deux visions de
l'espèce en une espèce dynamique ou
évolutive.
5. Les grandes lignes de l'histoire de la vie
a. L'échelle des temps géologiques
L'échelle internationale des temps géologiques est
une échelle stratigraphique
( la stratigraphie est la science des
dépôts sédimentaires, c'est-à-dire des
sédiments ayant donné naissance à des roches
sédimentaires). (nouvelle échelle
internationale de la Société géologique de
France et du Bureau de Rercherches géologiques et
minières, publiée en 1990, Bordas p 323; sur internet
au format PDF vous pouvez accéder aux dernières
moutures (mise à jour tous les 4 ans, la dernière en
2004) de l'échelle internationale sur le site de
l'ISC
(International Stratigraphic Chart) avec les conventions de la
Commission de la Carte Géologique du Monde (CCGM - CGMW):
http://www.stratigraphy.org/cheu.pdf
pour l'échelle des temps géologiques et pour le code
des couleurs: http://www.stratigraphy.org/codeu.pdf
ce qui est fort utile car avec les composantes en RGB on peut
représenter des couleurs exactes et non pas approximatives;
fait rarissime et remarquable:
cette échelle
présente des écarts d'incertitudes, pour les limites
situées avant le
Paléogène).
L'échelle est en fait scindée en trois
composantes:
- la lithostratigraphie (qui étudie la nature
minérale des couches sédimentaires) : les couches
sédimentaires sont alors regroupées selon leur
composition lithologique en membres, formations et
groupes.
- la biostratigraphie (qui étudie le contenu en
fossiles des roches sédimentaires) : les couches
sédimentaires étant habituellement divisées
en biozones. Les fossiles par comparaison avec les
espèces actuelles et en accord avec le faciès
lithologique (faciès déterminé à
partir de l'étude des minéraux de la roche
sédimentaire seuls) permettent de reconstituer le
faciès biologique et donc d'avoir une idée de
ce que pouvait être le milieu de vie de ces animaux
lorsqu'ils vivaient (c'est la
paléogéographie) : en ce sens ce sont des
fossiles de faciès (un
"bon" fossile de faciés doit être spécifique
d'un milieu le plus restreint possible (le mieux défini du
point de vue des ses paramètres environnementaux) mais par
contre avoir une extension chronologique maximale,
c'est-à-dire pouvoir être retrouvé pendant un
intervalle de temps le plus long possible). Mais ils permettent
aussi de réaliser des corrélations de datations de
couches parfois très éloignés
géographiquement. Un fossile
stratigraphique parfait aurait ainsi une extension
géographique mondiale (ou horizontale, maximale) pour une
extension chronologique (ou verticale) minimale
(c'est-à-dire qu'on ne le retrouverait que dans une seule
couche d'âge très précis).
Quelques exemples de groupes utilisés en
stratigraphie sont présentés p 322.
- la chronostratigraphie regroupe les données
lithostratigraphiques et biostratigraphiques en intervalles de
temps:
|
division de
temps
|
couches
|
ordre de grandeur de la
durée
|
définition
géologique
|
exemple
|
|
âge
|
étage
|
5 à 6 Ma
|
stratotype
*
|
Le nom de l'étage est le plus souvent
dérivé du nom du lieu où est
défini le stratotype, auquel on ajoute le suffixe
"-ien". Ainsi Lutèce (nom ancien de
Paris) a donné le lutétien.
|
|
époque
|
série
|
10 à 50 Ma
|
ensemble d'étages
|
Miocène
|
|
période
|
système
|
25 à 100 Ma
|
|
Crétacé
|
|
ère
|
érathème
|
60 à 500 Ma
|
|
Cénozoïque
(ère tertiaire)
|
|
éon
|
éonothème
|
> 500 Ma
|
|
Protérozoïque
|
|
* affleurement de référence dont le
contenu lithologique et biologique doit être le
plus riche possible (épaisseur de la couche,
nombre de faciès (le faciès d'une roches
sédimentaires est l'ensemble des conditions de
dépôt du sédiment qui a donné
naissance à cette roche: par exemple: un
faciès de haut de plage, un faciès de
récif tropical, un faciès de zone de
balancement des marées, un faciès de grand
fond d'une mer froide...), nombre d'espèces
différentes...)
|
Les limites entre les étages sont donc des coupures
chronologiques qui rendent compte à la fois de
phénomènes sédimentaires (par exemple fin d'une
période de transgression (avancée de la mer) et
début d'une période de régression (la mer se
retire)) et de l'évolution biologique: apparition de nouvelles
espèces et disparition d'autres espèces, du fait du
renouvellement des faunes et flores, au moins partiel entre chaque
étage. Les limites entres les intervalles de temps plus grands
sont elles aussi à rapporter à des
phénomènes géologiques globaux (montée ou
descente du niveau des mers, glaciation, émergence de
continents, forte période d'érosion...). Du point de
vue biologique les coupures chronologiques majeures sont souvent
associées à des crises, disparitions massives de
nombreuses espèces, suivies par des périodes de
radiation (développement brutal d'espèces nouvelles ou
d'espèces qui ont survécu au-delà de la limite
théorique).
b. Une application : la crise
Crétacé-Paléocène (ou la limite K-T)
Cet exemple à été choisi dans le programme
pour présenter la science paléontologique en cours
d'élaboration. Le fait que ses traces soient accessibles sur
le sol français et que les débats soient suivis par les
mass-média, prouve la pertinence du choix.
La véritable question est donc bien la limite K-T, la
notion de limite en stratigraphie, les moyens d'étude,
les arguments, l'historique du débat... il ne s'agit pas ici
de clore une question qui restera toujours du domaine de l'histoire
mais bien de montrer un exemple de discussion. Je précise que
ce qui est en discussion ce ne sont pas uniquement les causes, mais
la notion même de limite, le problème de la crise en
paléontologie, qui est loin d'être une question
définitivement tranchée.
Sur cette question le manuel scolaire Nathan est une
référence : la question y est très bien
traitée à mon avis (je salue ici le
travail de Stéphane Guellec), les principes clairement
exposés. Il me semble donc inutile d'essayer de mieux faire
(et puis surtout, c'est là que l'on voit les limites de la
géologie : cette science demande tellement de documents qu'il
est très difficile de présenter un cours qui soit un
résumé...).
c. Les grandes dates de l'évolution du vivant
Il faut bien connaître pour le bac quelques groupes
(Foraminifères, Trilobites et Ammonites incontournables pour
les invertébrés et pour les vertébrés les
Agnathes/Ganthostomes, Poissons, Amphibiens, Reptiles, Oixseaux et
Mammifères) et quelques dates clés (-450Ma : premiers
poissons; -420 Ma : premières plantes terrestres; -300 Ma:
premiers reptiles; -200 Ma : premiers oiseaux et premiers
mammifères; -120 Ma : premières plantes à
fleurs). Voir par exemple les p 312-313 et 325 du
Bordas et la synthèse p 329.
Introduction: Les objets et les
méthodes - 1. Les origines: un
problème à part - 2.Des
fossiles aux modèles - 3.L'évolution
- 4.Phylogénies - 5. Les grandes
lignes de l'histoire de la vie - 6. La
lignée humaine
retour plan cours
{kind=link}
{kind=link}