les hétérochronies,
variabilité des
processus embryonnaires ou interprétations ontogéniques
évolutives
Sources
Pour en finir avec le darwinisme; une nouvelle logique du
vivant, Rosine Chandebois, 1993, Éditions Espaces
34, p 167
Les éléments de réflexion proposés par
Stéphane Jourdan à l'adresse
http://www.ifrance.com/tahitinui//pinus/glossaire.html
me semblent particulièrement pertinents; j'ai fini par
accepter comme vrai ce qui m'est d'abord apparu comme une erreur
historique: les termes d'hétérochronie (et de
néoténie par extension) doivent être
exclusivement utilisés dans un sens
phylogénétique; l'embryologie ne peut pas utiliser ces
termes dans un autre sens sans prêter à confusion.
Sciences de la Terre et de l'Univers, coll.,
Vuibert, 1999, p 552-555
* du point de vue théorique, les informations les plus
facilement accessibles en français sur le net sont issues de
The Shape of Life Genes, Development and the
Evolution of Animal Form, Rudolf. A. Raff, 1996, Chapitre 8: It's not
all heterochrony.
* Les figures issues de Reilly et al. (Biol. Journal of the Linnean
Soc., 60, 119, 1997) sont reprises, notamment dans La
Recherche (Quand l'évolution change le temps des
êtres, Jean Chaline et Didier Marchand, La Recherche, 316,
janvier 1999, p 56-59), dans le manuel scolaire Nathan
TS, 2002, p 137, ou encore sur les pages de
Planet-Terre.
* De nombreux laboratoires de recherche, principalement en
anthropologie s'intéressent aux hétérochronies
mais les publications ne sont pas facilement accessibles et les
résumés permettent de douter que le terme soit vraiment
pris dans son sens exact. L'article de Fernando V. Ramirez Rozzi,
Hétérochronies : êtat des
lieux, dans Primatologie, 2000, 3, 479-511 est assez
éclairant à ce sujet. http://www.ivry.cnrs.fr/
deh/ramrozzi/tap/
heterochronies.pdf
Le but de ces lignes n'est pas de
présenter une vue exhaustive des utilisations de ce concept
mais d'essayer de clarifier l'utilisation du concept pour un
élève curieux en proposant ainsi des
éléments mémorisables.
Plan de cette page:
1.
Définitions
2. Les
hétérochronies se classent en 6
modalités
3. Des illustrations
: exemple d'interprétations hétérochrones
théoriques imagées
4. Un exemple de
données fossiles interprétées à l'aide
d'hétérochronies
5. Quelques
hétérochronies utilisées en
anthropométrie et en paléontologie
humaine.
retour accueil, plan
du cours de Terminale S
1 Définitions
Du point de vue étymologique le terme
hétérochronie (du grec
hétéro = différent et chronos = le
temps) désigne un décalage dans le temps
mais n'est pas utilisé par tous dans le même sens.
Historiquement, le concept vient des paléontologues, mais son
utilisation en embryologie est délicate, même si elle me
paraît plus riche de promesses:
Cependant le concept d'hétérochronie mérite
bien toute l'attention des embryologistes et pas seulement animaux,
même si les ontogénies animales et
végétales diffèrent profondément. Au
contraire, l'utilisation du concept d'hétérochronie, en
respectant sa dimension phylogénétique, doit
déboucher sur une embryologie évolutive.
Couplé à des données embryologiques, un
processus hétérochrone devient un vrai mécanisme
évolutif.
Remarque:
pour R. Chandebois, embryologiste, dont j'ai
essayé de présenter quelques idées sur mes
pages, les
hétérochronies
désignent des décalages mineurs dans
l'amortissement de la croissance post-embryonnaire en fin de
développement observées chez différentes
espèces proches. Dans sa
théorie
évolutive, elles seraient à
l'origine de la diversification des espèces, au sein de
lignées évolutives.
En effet, les décalages majeurs dans le temps, dus
à des facteurs accidentels au cours de l'embryogenèse,
conduisent le plus souvent à un arrêt de
développement et, dans certains cas, à des formes
monstrueuses. Dans d'autres cas ils ont été
rapportés comme des éléments essentiels de la
genèse de lignées. Un des exemples les mieux
documentés 2 est la sirénomélie, qui
se traduit par la fusion puis l'atrophie des membres
postérieurs, le développement exagéré de
kla boîte crânienne. A partir d'une étude
embryologique minutieuse sur des fœtus humains monstrueux
présentant cette malformation R. Chandebois propose comme
cause unique (mais avec des modalités variées)
«un déroulement trop rapide de la progression
autonome (PA - qui désigne l'automatisme embryonnaire chez
cet auteur - voir page
sur le développement) dans
l'ébauche de la chorde, au stade où elle fonctionne
comme centre organisateur de l'embryon». L'auteur va
jusqu'à proposer des modalités similaires de
modification du métabolisme mésodermique pour la
tendance à la réduction des pattes postérieures
et l'augmentation du nombre des vertèbres chez de nombreux
Vertébrés (anguilles chez les Poissons, Siren chez les
Amphibiens, serpents, Scinque et orvets chez les Reptiles...). Elle
ajoute qu'«il est fort possible que
l'adhésivité différentielle soit touchée,
d'un côté supprimant dans le mésenchyme des
flancs la compétence pour donner un bourgeon de membre, de
l'autre augmentant le nombre de rosettes (ébauches de somites)
par unité de volume».
Les hétérochronies ne désignent pas de tels
décalages mais des décalages de moindre amplitude qui
se passent en fin de développement. On sait que la
différenciation des tissus est un phénomène
progressivement amorti. En fin de développement, les tissus
prolifèrent (croissance post-embryonnaire) sous le
contrôle de nombreuses hormones. Mais cette croissance
s'amortit progressivement, probablement du à une perte de
compétence des tissus différenciés. Rosine
Chandebois2 refuse le statut d'interprétation
évolutive aux hétérochronies. Pour elle ce sont
des modalités bien connues de la diversification des
espèces et non des mécanismes évolutifs (au sens
de mécanismes majeurs de la variation qui conduit à
l'apparition des embranchements par exemple).
2. Les hétérochronies se
classent en 6 modalités
On classe THÉORIQUEMENT les
hétérochronies (ou processus
hétérochrones) de la façon suivante
- taux de développement (ou de croissance)
accéléré (accélération) ou ralenti
(décélération)
- début de développement plus précoce
(prédéplacement) ou plus tardif
(postdéplacement)
- fin du développement plus précoce (progenèse)
ou plus tardive (hypermorphose).
|
peramorphoses
("sur-développements" ) - - - - -
- pædomorphoses*
("sous-développements"
)
|
|
|
accéléré ou
précoce
|
ralenti ou
tardif
|
|
taux de
développement
(= vitesse ou/et
intensité de
développement selon les cas)
|
accélération
|
décélération
(néoténie)
|
|
.
|
.
|
|
début de
développement
|
prédéplacement
|
postdéplacement
|
|
.
|
.
|
|
fin de
développement
|
progenèse
(hypomorphose**)
|
hypermorphose*
|
Remarques:
* pædomorphose
s'écrit aussi paédomorphose ou
paedomorphose ce qui aide à éviter de confondre
les lettres "æ" et "œ".
** L'hypermorphie et l'hypomorphie sont des
états de sur- et de sous-développement morphologique
alors que l'hypermorphose et l'hypomorphose sont des
processus aboutissant aux états précités et qui
entrent, dans l'acception qui est ici soutenue, dans le cadre d'un
processus hétérochrone.
3. Des illustrations : exemple
d'interprétations hétérochrones
théoriques imagées
On ne peut pas donner d'exemple d'hétérochronies,
comme quelque chose de connu, mais bien des exemples
d'interprétations à l'aide
d'hétérochronies ce qui rend beaucoup plus difficile
leur présentation pédagogique.
Voici celle de Sciences de la Terre et de l'Univers,
coll., Vuibert, 1999, p 553-554 (fig 13-54,
légèrement modifiée), qui me
paraît nettement plus claire, une fois l'effort de
compréhension fait, que les habituelles courbes. Les
schémas et le texte de l'encadré de la p 57 de
L'évolution, Belin-Pour La Science,
Bibliothèque Pour La Science, 1998 (Les
hétérochronies du développement par Didier
Néraudeau), sont aussi tout à fait corrects.
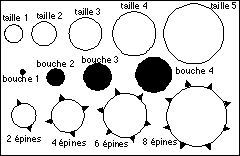
L'ontogenèse est illustrée par 3
caractères: la taille de l'organisme (taille 1
à 5), la taille de la bouche (de 1 à 4) et le
nombre d'épines (0,2,4, 6 ou 8).
|
Le développement normal de cet "organisme"
pédagogique comporte 4 étapes A,
B, C et D, représentées ci-dessous. Dans la
figure originale les durées croissantes de chaque
phase (A, B,C et D) étaient
représentées par des rectangles de longueur
croissante, ce qui n'a pas toujours été
respecté ici.
|
Pour chaque type d'hétérochronie deux exemples sont
proposés selon une modalité globale (taille
globale, taille de la bouche et nombre d'épines augment ou
diminuent corrélativement = c'est une allométrie
positive) et selon une modalité locale
(dissociée) où seule la taille de la bouche
augmente plus ou moins vite selon le type
d'hétérochronie; la longueur des rectangles des
étapes de développement est représentative de
leur durée.
|
peramorphoses
("sur-développements" ) - - - - -
- pædomorphoses
("sous-développements"
)
|
|
|
précoce
|
tardif
|
|
début de
développement
|
|
|
A
|
B
|
C
|

|

|

|
|
hétérochronie
globale
|
|
|
Pour une durée de
développement allongée, la taille globale de
l'animal, la taille de sa bouche et le nombre
d'épines sont tous les trois
augmentés
|
Pour une durée de
développement diminuée, la taille globale de
l'animal, la taille de sa bouche et le nombre
d'épines sont tous les trois
diminués
|
|
prédéplacement
|
postdéplacement
|
|
|
A
|
B
|
C
|
D
|
|
|
|

|

|

|
|
hétérochronie
locale (dissociée) limitée à
la bouche
|
|
|
|
Pour une durée de
développement allongée, la bouche est plus
grande car sa taille a augmenté
précocement
|
Pour une durée de
développement identique, la bouche est plus petite
car sa taille a augmenté
tardivement
|
|
développement de l'organisme
ancestral de
référence
|
|
|
|
peramorphoses
("sur-développements" ) - - - - -
- pædomorphoses
("sous-développements"
)
|
|
|
précoce
|
tardive
|
|
fin de
développement
|
|
A
|
B
|
C
|
|
|
|
|
|
hétérochronie
globale
|
Pour une durée de
développement diminuée, la taille globale de
l'animal, la taille de sa bouche et le nombre
d'épines sont tous les trois
diminués
progenèse
(hypomorphose)
|
A
|
B
|
C
|
D
|
|
|
|
|

|
|
hétérochronie
locale (dissociée) limitée à
la bouche
|
Pour une durée de
développement identique, la bouche est plus petite
car sa taille a cessé d'augmenter
précocement
|
Pour une durée de
développement augmentée, la taille globale de
l'animal, la taille de sa bouche et le nombre
d'épines sont tous les trois
augmentés
hypermorphose
Pour une durée de
développement augmentée, la bouche est plus
grande car sa taille a continué d'augmenter
tardivement
|
|
développement de l'organisme
ancestral de
référence
|
|
|
4. Un exemple de données fossiles
interprétées à l'aide
d'hétérochronies
Un exemple d'interprétation "hétérochrone" de
formes actuelles (il serait préférable de dire: un
exemple d'interprétation mettant en jeu des processus
hétérochrones): Cardium plicatum et Cardium
fittoni (in Sciences de la Terre et de l'Univers, coll.,
Vuibert, 1999, p 554, fig 13,54 repris d'après Skelton,
1993).
|
Chez Cardium
fittoni, l'adulte mesure environ 35 mm de large
et son ornementation ressemble très fortement
à celle de C. plicatum juvénile
|
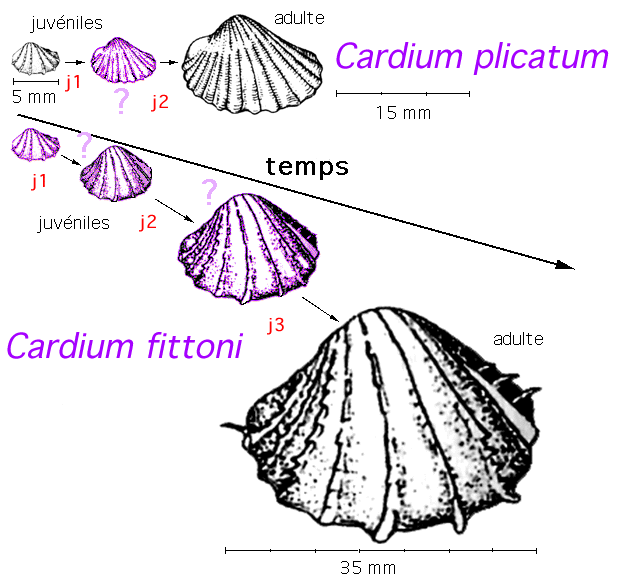
|
Cardium
plicatum juvénile mesure environ 5 mm de
large et atteint près de 17 mm à l'état
adulte mais son ornementation change (le nombre de
côtes augmente et celles-ci sont moins
marquées.
|
Une illustration
de l'ontogénie comparée à partir de
deux espèces fossiles supposées dériver
l'une de l'autre phylogénétiquement;
au stade juvénile j1 les deux
espèces seraient morphologiquement semblables et
commenceraient à différer au stade j2; C.
fittoni atteint à l'âge adulte une taille
supérieure à celle de C.
plicatum
(les dessins intermédiaires
en violet
sont
INVENTÉS
et donc FAUX d'après
Sciences de la Terre et de l'Univers, coll., Vuibert, 1999,
p 554, fig 13,54, eux-mêmes d'après Skelton,
1993, très modifiés).
|
|
Les "données"
paléontologiques que l'on ne remet pas en
question dans cet exemple:
Remarque:
certaines, pour lesquelles je n'ai pas trouvé
d'information, sont des hypothèses qui me semblent
nécessaires
|
* on considère comme acquis, même si ce
n'est qu'une hypothèse paléontologique, que la
petite forme est bien la forme juvénile de la grande
forme, cette dernière étant la plus
ornementée des deux, et qu'ils forment une seule
espèce paléontologique: Cardium
plicatum.
* on considère comme acquis que la forme
représentée ici est une forme adulte qui
constitue une espèce paléontologique :
Cardium fittoni, sans que l'on connaisse la forme
juvénile, à moins qu'elle ne soit identique
à la forme adulte en plus petit (?).
* il existe une assez forte ressemblance morphologique entre
la forme juvénile de C. plicatum et C.
fittoni (ce qui n'est pas trop évident sur ces
dessins).
* C. fittoni dérive
phylogénétiquement de C. plicatum (par
exemple du fait de l'âge auquel il appartient -
au sens paléontologique de division du temps
correspondant à un étage: C. fittoni
est une espèce postérieure (à quelques
millions d'années près) à C.
plicatum).
* le taux de croissance est supposé identique chez
les deux espèces paléontologiques.
|
règles
|
règle
1. En paléontologie la règle
est de considérer comme
"sous-développée" (pædomorphe)
la forme adulte qui ressemble à un
ancêtre juvénile.
|
|
règle
2. Ensuite on attribue cette morphologie
à une néoténie si le
descendant a, au moins, la taille de
l'ancêtre adulte et à une
progenèse si le descendant a la taille de
l'ancêtre juvénile.
|
|
Mais...
l'état adulte (capacité à se
reproduire) peut être atteint avant que la taille
maximale des individus ayant donné des fossiles
connus soit atteinte ou, au contraire, après
celle-ci, si les fossiles des individus les plus grands
n'ont pas été retrouvés.
|
|
Si l'on suit les règles données, cette
ressemblance doit donc être interprétée
comme une pædomorphose qui peut prendre
deux aspects: C. fittoni dérive de C.
plicatum par une ralentissement du développement
global (décélération ou
"néoténie") et/ou une
hypermorphose. En effet, le ralentissement
n'empêche pas que - ou même induit le fait que -
le développement se poursuive plus longtemps chez
C. fittoni que chez C. plicatum, ce qui peut
être considéré comme une hypermorphose,
la taille de C. fittoni étant nettement
supérieure à celle de C. plicatum
On observe également une augmentation locale du
développement des côtes (qui sont donc plus
marquées chez C. fittoni que chez C.
plicatum) mais une réduction de leur nombre.
Cette morphologie ne peut pas résulter d'un
mécanisme d'hypermorphose seul (puisqu'il devrait
s'accompagner d'une augmentation du nombre de côtes)
alors qu'il est beaucoup plus simple de la rapporter
à une décélération
(néoténie) locale qui touche principalement le
nombre de côtes.
|
5. Quelques
hétérochronies utilisées en
anthropométrie et en paléontologie humaine.
Des études de biométrie réalisées par
exemple par Anne-Marie Guihard-Costa (Directeur de
recherche au C.N.R.S., UPR 2147 "Dynamique de l'Évolution
Humaine : Individus, Populations, Espèces", voir
http://www.ivry.cnrs.fr/deh/guihard/guihard.htm) ont permis de
mettre en évidence l'existence d'une modulation des vitesses
de croissance au cours de la vie fœtale avec une
décélération marquée en fin de
gestation. De même cette auteur a développé
la caractérisation d'une "période de transition
adaptative" autour de la naissance, débutant 5 à 6
semaines avant la naissance, au cours de laquelle on observe une
augmentation considérable de la variabilité
individuelle, chez les sujets normaux.
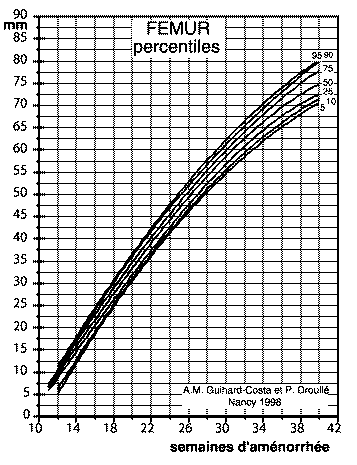
Courbe de croissance standard du
fémur
(http://www.ivry.cnrs.fr/deh/guihard/fig1.htm)
(ces courbes standard sont établies à
l'aide de très nombreuses données recueillies sur 490
fœtus et nouveau-nés "normaux" (sujets issus de
grossesses uniques, non malformés, ne présentant pas
d'anomalies chromosomiques), provenant de l'hôpital de
Port-Royal, Paris (Dr J.C. Larroche) et 673 fœtus et
nouveau-nés "normaux", provenant de l'hôpital Robert
Debré, Paris (Dr. A.L. Delezoide) et de données
échographiques (359 fœtus, mesurés entre 10 et 42
semaines d'aménorrhée à la Maternité
régionale de Nancy (Dr. P. Droullé) pour les
données transversales, et 24 sujets suivis dans les
périodes pré- et périnatale pour les
données longitudinales).
Une telle courbe permet de déterminer les limites entre la
croissance "normale" et la croissance retardée, pathologique
et est utilisée couramment dans plusieurs grandes
maternités françaises
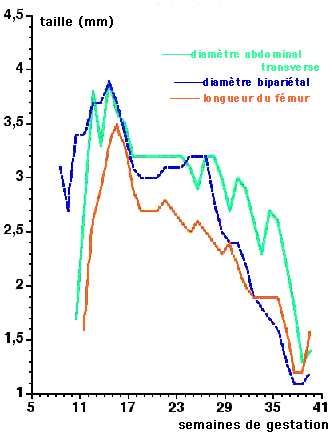
Courbes moyennes de vitesse de croissance
comparées
entre les diamètres bipariétal et abdominal transverse
et la longueur du fémur
(http://www.ivry.cnrs.fr/deh/guihard/fig2.htm)
Ces courbes montrent une modulation des vitesses de croissance au
cours de la vie fœtale avec une
décélération marquée en fin de gestation.
Attention ces courbes ne représentent pas toutes le même
type de croissance (une croissance en longueur et deux croissances en
diamètre), les organes sont différents (un os long, des
os crâniens formant la boîte crânienne et des
viscères) et n'atteignent pas une maturité identique en
même temps.
En tant que telle une décélération de
croissance, comparativement au rythme de la croissance
déjà irrégulier au cours de la vie fœtale,
n'est aucunement un processus hétérochrone. Pour
le qualifier ainsi il faut une comparaison avec des organismes
reliés phylogénétiquement. Des
comparaisons doivent donc être menées sur des
embryons de Primates dans la lignée humaine.
Ces résultats, replacés dans une perspective
évolutive, permettent cependant d'intéressantes
hypothèses. Par exemple on peut considérer cette
période d'instabilité de fin de développement
fœtal comme une possibilité d'accroissement des
potentialités adaptatives du fœtus. De même
l'analyse et la comparaison des rythmes de croissance pré- et
post-nataux chez les Primates, replacées dans leur contexte
d'environnement écologique, peut sans aucun doute aider
à définir le rôle adaptatif de ces processus de
croissance. Ces travaux sont développés par exemple
dans l'équipe UPR 2147 du CNRS, Dynamique de
l'Évolution Humaine (http://www.ivry.cnrs.fr/deh/themes.htm#theme2)
Processus ontogéniques et hétérochronies
(A.-M. Guihard-Costa, G.Berillon, Y.Deloison, P.Droullé,
J.-C.Pineau, H.Pucciarelli, F.Ramirez-Rozzi). Les processus
ontogéniques étudiés morphométriquement
sont: d'une part le développement fœtal et
néonatal humain, en général, et d'autre part,
des points plus précis comme le développement dentaire,
la morphologie crânienne ou la dimension longitudinale du
visage ou encore la morphogenèse postnatale du pied au cours
de l'apprentissage de la marche. Les mécanismes
hétérochroniques sont par exemple appliqués
à l'étude des dents des Hominidés
plio-pleistocènes.


