De la biodiversité à
l'histoire des êtres vivants
(2ème
partie)
retour accueil, plan
du cours de terminale S
Introduction (voir partie
précédente)
Plan de cette page:
2. Phylogénie des
vertébrés (un cours de terminale)
- a. comparer deux
vertébrés suppose plusieurs théories
notamment: une théorie du vivant
(de l'unité du vivant), une théorie de la
forme et une théorie de
l'évolution...
- b. classer des
vertébrés à l'aide de
caractères
- b1. les plumes, caractères de
vertébrés
- b2. généralisation: homologies et
analogies
- c. deux phylogénies et une histoire des
vertébrés
- a1 Un cladogramme SIMPLIFIÉ des
vertébrés
- b2 Une phylogénèse naturelle des
vertébrés
3. l'origine des
vertébrés
2. Phylogénie des
vertébrés
Le programme nous propose un vaste groupe d'organismes,
l'embranchement des Vertébrés, dont nous allons
comparer phylogénétiquement les espèces,
unité de classification la plus simple. Nous reportons la
discussion de l'origine des Vertébrés à la
partie suivante, bien qu'il apparaîtra que la question de
l'origine doit toujours être traitée en premier.
une phylogénie
(du grec phulon = race, et
genèse ou génie) c'est une
classification évolutive
la phylogénèse c'est l'étude
ou l'établissement des phylogénies
(pour une approche historique,
voir chapitre
précédent)
|
a. Comparer deux vertébrés (qui se ressemblent)
suppose plusieurs théories: notamment:
une théorie du vivant (de l'unité du vivant),
une théorie de la forme et une théorie de
l'évolution...
Soyons simpliste avec un "petit" exemple... qui va nous permettre
de poser les questions fondamentales et d'y apporter des
réponses provisoires (les caractères
présentés ici ne sont qu'un exemple de ceux qui peuvent
être proposés par les élèves, ou
l'enseignant, le tout étant qu'ils recouvrent les quatre
domaines exposés ici: caractères écologiques,
caractères morphologiques ou anatomiques ou embryologiques,
caractères physiologiques, caractères
moléculaires:
|
Comparons
sommairement un thon
et un
dauphin.


|
des ressemblances...
... mais des différences
|
... qui permettent de poser
des questions...
|
... et d'y apporter des
réponses, souvent provisoires et parfois sous forme
d'hypothèses
|
|
une même forme
hydrodynamique ...
... mais on reconnaît la
silhouette de ces deux animaux au premier coup
d'œil
(caractères
morphologiques, anatomiques, physiologiques,
embryonnaires...)
|
La forme (d'un organisme mais aussi
d'un organe, d'une cellule...) est un caractère
morphologique, à quoi est-elle due ?
D'où cette ressemblance
tire-t-elle son origine ?
Cette ressemblance (entre deux
adultes) se met-elle en place lors du développement ?
D'une façon encore plus
générale sur quelle causalité repose la
forme des vivants ?
Quelle est la cause la plus proche
qui est déterminante pour la forme d'un être
vivant ?
|
Ils se ressemblent parce que ce
sont tous deux des êtres vivants. Le principe de
l'UNITÉ du
vivant fait reposer la
genèse de la forme sur des structures et
mécanismes fonctionnels communs (que j'appelle
travail): la structure unité c'est la cellule
(toutes les formes vivantes sont composées de
cellules, c'est la théorie cellulaire
vue en seconde); les mécanismes fonctionnels communs
sont appelés travail du vivant: travail de
relation, de reproduction et de nutrition
(voir
cours
de seconde ou
page
sur la diversité du
vivant)
Mais...
Les ressemblances et différences de forme
peuvent être causées par:
* des contraintes
EXTÉRIEURES, du milieu,
qui ORIENTERAIENT le développement, qui modifieraient
les interactions entre populations cellulaires
(viscosité de l'eau, pression de la colonne
d'eau...); ce qui suggère, que placé dans un
autre milieu, l'embryon pourrait se développer
autrement....ce qui n'est pas le cas (ce n'est pas l'embryon
qui a une forme hydrodynamique, c'est l'adulte...les
caractères de l'adulte résultent de ceux de
l'embryon mais ne sont pas ceux de l'embryon)... On pense
donc que la forme de l'espèce est
innée,
transmise héréditairement, stable;
même si des contraintes au cours du
développement peuvent provoquer des anomalies, des
monstruosités... on ne pense pas qu'elles puissant
jamais causer cette adaptation de l'animal avec son
milieu de vie, qui est donc une donnée dont on
doit rechercher la causalité à
l'intérieur du vivant.
Mais la forme de l'adulte n'est pas non plus celle de
l'embryon. La forme de l'embryon pourrait-elle être
sous la dépendance de facteurs externes ? Cette fois
la réponse est
affirmative.
On peut, en
augmentant par exemple la salinité du milieu,
provoquer une exogastrulation chez la gastrula de grenouille
(la gastrulation est l'étape où la couche de
cellules la plus externe de l'embryon se déplace pour
aller tapisser la cavité interne de la gastrula (voir
photos
de l'hermelle
dans le cours de seconde); dans le cas d'une
exogastrulation, les mouvements des cellules au lieu de
provoquer l'invagination des tissus, provoquent leur sortie
vers
l'extérieur).
La forme qu'il faut envisager
n'est donc pas unique. Chaque forme, à chaque instant
du développement, celle du zygote après la
fécondation dans l'eau, celle de l'embryon au stade
gastrula au milieu de ses tissus protecteurs, celle du
fœtus dans les annexes fœtales, celle du jeune
à l'éclosion libéré ou non dans
le milieu extérieur..., peut être sous la
dépendance des conditions extérieures qui
maintiennent celle-ci tant que l'être vivant
possède la capacité à changer de forme:
période que l'on pense être limitée au
développement pour un organisme dans son ensemble,
mais qui peut être certainement beaucoup plus longue
pour un organe. Ainsi la forme de nos organes pourrait
dépendre des interactions, au cours de leur
croissance, des populations qui les composent avec le milieu
qui les environne ? Il s'agit là
d'une théorie
dynamique de l'hérédité de la
forme. Selon le mot de S.
F. Gilbert, un embryologiste de renom: « il n'y a
pas de prédiction du phénotype par le
génotype.»
(Biologie du
développement, De Boeck Université, 1996, p
74)
* un déterminisme
INTERNE; deux grandes
réponses sont proposées:
- soit un programme
génétique de développement,
transmis sous forme d'ADN, stable mais fragile
(mutations, délétions...), molécule
qui stockerait non seulement l'information pour les
molécules du vivant mais aussi pour l'ordre dans
lequel elles doivent être
synthétisées, utilisées...certains
(matérialistes du point de vue philosophique) y
voient même un programme qui déterminerait
la totalité des processus de l'être vivant,
ce qui est le sens "grand public" de "programme
génétique"
(voir discussion
de ce mot dans les commentaires
du programme de
seconde);
- soit une mémoire
cytoplasmique de l'espèce (qui comprend des
éléments chimiques transmis d'organisme
maternel à la cellule sexuelle maternelle ou
ovocyte, à nulle autre pareille: ARN
maternels, a.a., protéines....mais aussi des
éléments non chimiques, propres à
cette cellule, comme sa compartimentation cytoplasmique
originale...), à laquelle s'ajoute bien sûr
l'information génétique qui ne
serait qu'une information pour des molécules et en
aucun cas un programme; dans ce cas la vie reste un
mécanisme original dont l'autonomie
s'exprime par le travail du vivant.
|
|
CONCLUSION:
Pour comparer des formes vivantes il nous faut une
théorie des formes et une théorie du
vivant. Comme plusieurs sont actuellement disponibles,
la moindre des choses pour un enseignant est d'en esquisser
les contours... vous n'êtes pas obligés de
choisir, même si l'enseignant peut vous exprimer son
choix...
|
|
leurs cellules musculaires
synthétisent une myoglobine...
... mais la séquence des
acides aminés composant cette molécule unique
de 153 aa - associée à au même
groupement ferreux (hème) que chacune des 4
chaînes de l'hémoglobine - est
différente. De plus la quantité de
myoglobine dans les cellules musculaires du dauphin
est extrêmement importante (les muscles du dauphin
comme celui du cachalot ou du phoque ne sont pas rouges mais
bruns - couleur due à l'abondance de cette
molécule). Cette abondance est mise en relation avec
la capacité, pour cet animal, de rester en immersion
de façon prolongée, malgré sa
respiration pulmonaire, qui ne peut donc se faire
qu'à l'émersion.
Le thon possède bien
sûr une respiration branchiale et sa respiration est
aquatique.
|
Sur quoi repose un caractère
moléculaire comme la séquence primaire d'une
protéine (comment et sous quel contrôle est
synthétisée cette molécule) ?
Quelle est, pour une
protéine, la relation entre la séquence
primaire, secondaire, tertiaire, quaternaire et la fonction
(deux séquences différentes peuvent-elles
correspondre à des protéines ayant la
même fonction et la même efficacité) ?
Dans le cadre d'une
interprétation évolutive comment peut-on
envisager de passer de la synthèse d'une
molécule à une autre ?
|
L'unité
du vivant au niveau
moléculaire peut être qualifiée
d'unité chimique.
En atteignant le niveau moléculaire, on pense
s'affranchir de la vie. Est-ce possible ? La matière
vivante diffère-t-elle de la matière
inorganique ? Est-elle constituée de molécules
inanimées
(avec les mots de
Principes de Biochimie de A. Lehninger, D. Nelson et M. Cox,
Médecine-Sciences, Flammarion, 1994, p
3) ? Pour un philosophe, il est
clair que la matière vivante n'échappe pas au
problème du vivant. Tout biochimiste sérieux
n'est pas forcément matérialiste.
En effet, comme toute biologie, la biochimie repose sur une
théorie du vivant, peut-être plus difficile
à formuler au niveau moléculaire, mais la
question ne doit pas être évacuée.
Les résultats de la biologie moléculaire
forment actuellement un ensemble cohérent que l'on
pourrait qualifier de
biologie moléculaire
du gène et qu'il est
indispensable de séparer d'une théorie du
vivant particulière.
Un gène est une unité de fonction qui, dans sa
définition la plus simple, est un fragment d'ADN
associé à la synthèse d'une
molécule, principalement une protéine. Un
même caractère moléculaire chez deux
individus résulte donc probablement du fonctionnement
d'un même gène, du fait de l'unité du
vivant. Les mêmes mécanismes de synthèse
de protéines intervenant chez les deux individus, il
est très probable que les gènes soient
très voisins, si ce n'est identiques. Plus qu'une
unité génétique il s'agit d'une
véritable unité biochimique,
métabolique. A l'échelle moléculaire,
les molécules du vivant sont toujours
supposées avoir une même origine, une
même fonction, les mêmes
propriétés. La matière organique
obéit aux lois de la chimie. Mais la biochimie n'est
pas qu'une extension de la chimie. Dans le détail il
existe des spécificités du vivant, et surtout
de chaque individu, ou plus encore de chaque espèce,
qui, sans violer les lois de la chimie, dépassent une
vision unifiée simpliste du vivant
considéré comme un composé
organique...
|
Quelques
"rappels" liés au programme de 1ère
S
* L'ADN et l'ARN
(acides nucléiques) contiennent
l'information pour les molécules
(information génétique)
sous la forme de triplets de bases azotées;
elle peut passer de l'ADN à l'ARN par
transcription ou de l'ARN à l'ADN par
transcription inverse. L'information
génétique est exprimée
(traduite) par la machinerie cellulaire (qui
comprend des ARN (messagers, ribosomiaux, de
transfert...), de nombreux acides nucléiques
monomèriques à rôle
énergétique (ATP, GTP...), de
nombreuses protéines...).
* La synthèse des protéines
est la mieux connue mais surtout c'est elle qui
soutient la vision la plus unitaire dans le monde
vivant. Les protéines ont un rôle
à la fois structural (comme constituant du
vivant) et fonctionnel (Elle commence par la
formation des ARNm (messagers) par des processus
complexes de maturation (l'ARN issu de la
transcription d'une chaîne d'ADN (transcrit
primaire) est souvent découpé,
modifié par l'ajout de séquences
répétées en début et en
fin de molécule...). Les acides
aminés sont soit issus de l'alimentation
protéique de l'organisme soit
synthétisés par l'organisme (cas le
plus général). La synthèse des
protéines se continue par l'assemblage des
acides aminés de chaque polypeptide
(structure primaire) au sein de ribosomes
(composés d'ARNr et de protéines)
grâce à la correspondance (c'est le
code génétique, quasi universel)
entre chaque triplet de bases de l'ARNm (messager)
et un acide aminé transporté sous
forme de complexe avec un ARNt. Les polypeptides
ainsi formés subissent de très
nombreuses transformations de maturation (ajout ou
excision de groupements, par exemple les nombreuses
méthylations, l'ajout de chaînes
glucidiques pour les glycopeptides...) avant de
former des protéines, la plupart du temps
issues de l'assemblage de plusieurs chaînes
polypeptidiques.
* Les glucides ainsi que les lipides et les autres
molécules apparentées aux lipides
comme les alcools ou stéroïdes, sont
réputés pour avoir une
spécificité
(originalité au niveau de
l'espèce, ou au niveau de l'individu)
moindre, voire inexistante. Ainsi, rien ne
différencie une molécule de glucose
sanguin du thon de celle du dauphin. Mais cela
n'est pas vrai pour un glucide complexe: nombre de
glucides ramifiés forment des motifs de
reconnaissance à la surface des membranes
(on les qualifie de lectines dans le cas de la
paroi bactérienne), associés à
des protéines (glycoprotéines) ou
à des lipides (glycolipides). Les
molécules simples peuvent aussi avoir des
rôles spécifiques selon leur
localisation: un acide aminé comme la
glycine peut être un neurotransmetteur: il
sera alors stocké dans des vésicules
synaptiques et libéré au voisinage de
récepteurs postsynaptiques
spécifiques. Les molécules simples
sont aussi transportées par des
molécules complexes protéiniques qui
déterminent alors des
spécificités supplémentaires,
par exemple les protéines associées
aux lipides (VDL, HDL et autres gouttelettes
lipidiques).
On pense que toutes ces spécificités
résultent de l'activité de
protéines (enzymes..) et d'acides
nucléiques (ribozymes...). En dernier
ressort, toutes les réactions du
métabolisme (comprenant le catabolisme, qui
regroupe les dégradations, et l'anabolisme,
qui regroupe les synthèses), sont
réputées être
contrôlées par les gènes, ou au
moins par les acides nucléiques et leurs
produits. C'est ainsi que l'on a une vision
unifié de la biochimie du vivant, que l'on
peut qualifier de biologie moléculaire du
gène.
|
* Les molécules complexes comme les protéines
ne sont presque jamais totalement identiques chez deux
individus d'espèce différente. Par contre
elles sont souvent semblables au sein de la même
espèce. Dans la théorie du vivant la plus
répandue, les gènes définissant les
caractères de l'espèce, l'espèce se
définit donc par l'ensemble de ces gènes
(chaque individu ne possédant qu'une partie des
gènes de l'espèce...devenue ainsi une notion
relative). On retombe donc ici sur une théorie du
vivant discutée plus haut.
* Deux molécules
différentes peuvent provenir de la traduction d'un
même gène du fait de la complexité des
étapes de la maturation des ARNm. La correspondance
une protéine-un gène n'est donc pas simple
à mettre en évidence car elle repose sur une
grand nombre de processus. L'hypothèse d'un
gène semblable lorsque l'on a deux molécules
semblables, ne sera toujours qu'une hypothèse,
raisonnable, mais jamais une certitude.
* Deux molécules
différentes mais proches peuvent toujours avoir la
même fonction mais cette fonction n'est pas
forcément remplie avec la même
efficacité. Le postulat selon lequel tout
système vivant optimise son fonctionnement, est
certainement critiquable.
Une modification de la structure primaire de la
protéine peut avoir des conséquences
variables:
- dans le cas de la myoglobine; je ne connais pas
précisément les modifications de
séquence entre les deux molécules de
myoglobine du thon et du dauphin et je n'ai trouvé
aucune information sur une éventuelle modification de
la fonction de la molécule qui est pourtant si
importante dans l'oxygénation du muscle de
vertébré. Cependant, comment imaginer qu'une
substitution d'acide aminé ne modifie pas les valeurs
de la diffusion facilitée du dioxygène pour
les pressions partielles de dioxygène du muscle au
repos et en activité ? Les données disponibles
sont trop souvent partielles et surtout limitées
à quelques organismes.
- dans le cas du collagène, des cas
d'anomalies dans la structure tertiaire de la
molécule (structure hélicoïdale
altérée) ont été
attribuées à des substitutions simple d'un
acide aminé. Ces substitutions sont rapportées
à des maladies graves: la substitution d'une glycine
par une cystéine a été rapportée
à l'osteogeneis imperfecta et celle d'une
autre glycine par une sérine a été
rapportée au syndrome d'Ehlers-Danlos. La
première conduit à une formation osseuse
anormale chez les bébés et la seconde à
une distension des articulations. Les deux peuvent
être létales.
Que dire des autres anomalies: n'existent-elles pas ?
Sont-elles trop graves pour être conservées
chez l'embryon ?
* On peut enfin faire des
hypothèses évolutives lorsque l'on compare des
molécules. Le raisonnement de départ est assez
séduisant: plus deux molécules se ressemblent,
plus leurs gènes sont proches, et du fait de
l'unité du vivant, plus il est probable qu'ils soient
apparus sous une forme unique qui se serait ensuite
diversifiée. Dans un tel raisonnement, en plus d'une
théorie évolutive, posée comme
hypothèse de départ, les postulats logiques
sont très nombreux (voir aussi l'ancien
cours de TS, &4a3, dont
quelques éléments sont repris ici):
- le plus important me semble être l'hypothèse
logique de l'optimisation des processus (voir
l'encadré sur le
finalisme dans la
première partie de ce cours). Il se traduit par
l'hypothèse d'un gène unique, évoluant
de façon importante, plutôt que
l'hypothèse de deux gènes, plus stables.
- une phylogénie basée sur des similitudes
moléculaires (séquence d'acides
nucléiques ou d'a.a.) n'apporte aucune PREUVE de
filiation mais est une ILLUSTRATION d'une filiation
posée comme hypothèse de départ: c'est
ce que l'on peut appeler de l'évolution
moléculaire phénotypique. En voici
quelques éléments logiques:
* 1er postulat (postulat évolutif): les
molécules chimiques évoluent et sont le reflet
de l'évolution des formes des êtres
vivants.
En effet, si l'évolution des êtres vivants est
une interprétation soutenue par d'innombrables
arguments paléontologiques, l'évolution des
molécules n'a aucun argument paléontologique ;
elle est une hypothèse dans le cadre de
l'interprétation évolutive ; elle n'apporte
aucun argument à l'évolution. Donc les
conclusions obtenus ne sont pas des arguments
évolutifs se sont des arguments d'évolution
moléculaire. Étant bâtie sur une
hypothèse, ses conclusions ne peuvent dépasser
cette hypothèse.
Par contre on peut utiliser le critère de
ressemblance mais dans ce cas, non pas comme argument
évolutif mais comme argument pour classer les
espèces. Les arguments moléculaires sont des
critères de classification mais non évolutifs.
Une classification évolutive repose elle aussi sur la
même hypothèse et l'on risque de tomber dans le
même sophisme que précédemment.
L'évolution posée comme hypothèse, on
classe les espèces avec des critères
uniquement considérés dans le cadre de cette
hypothèse. La valeur de ces classifications ne
dépasse pas les limites de l'hypothèse de
départ.
* 2ème postulat (postulat de
spécificité) : les molécules
chimiques se ressemblent plus au sein d'un même groupe
qu'elles ne différent entre individus ou chez un
même individu. En effet, on peut dire qu'il existe
une signature moléculaire de l'identité de
l'individu, de l'espèce et des groupes d'organismes
d'ordre supérieur, selon les groupes comparés.
Les espèces actuelles reflètent, dans leur
composition moléculaire, les similitudes entre
individus d'une même espèce, d'une même
classe, d'un même embranchement (on gomme les
variations phénotypiques individuelles ou on suppose
qu'on peut les isoler).
* 3ème postulat, le plus paradoxal (postulat de
stabilité) : les molécules
utilisées n'ont pas évolué dans les
groupes considérés et les différences
observées actuellement sont les traces des
différences passées
(hypothétiques). La seule évolution qui
est acceptée est celle, discontinue donc, entre les
groupes. Il ne peut être considérée
d'évolution individuelle ou intraspécifique.
Bien entendu, l'évolution individuelle au cours de la
vie de chaque individu ne peut pas non plus être prise
en compte.
|
|
le collagène est une
protéine qui compose principalement les tendons et la
matrice des os...
... mais chaque molécule de
collagène est habituellement constituée de 3
hélices enroulées les une autour des autres et
formant une superhélice
(qui détermine
la résistance de la molécule à la
tension mais pas à
l'étirement). La
séquence des acides aminés composant chaque
hélice peut changer légèrement (une
molécule de collagène contient habituellement
35% de glycine, 11% d'alanine, 21% de proline et
hydroxyproline - un
aa dérivé du précédent et
portant un OH sur le carbone
n°4): en effet, la forme
en hélice de chaque molécule (structure
tertiaire) est due à une séquence
répétée de 3 acides aminés
(glycine-X-proline) ou (glycine-X-hydroxyproline), qui
détermine un tour d'hélice.
Les molécules
de collagène peuvent s'associer en fibres
constituées d'empilement de molécules de
tropocollagène elles-mêmes formées par
des hélices de trois molécules de
collagène...
|
|
caractères
moléculaires
|
|
Conclusion:
Pour comparer des molécules du vivant, il nous faut
l'outil de la biologie moléculaire mais encore
et toujours une théorie du vivant et une
théorie de l'évolution. Là
encore, les options sont multiples.
|
|
un même milieu de vie
...
... mais un mode de vie
différent: le dauphin revient à la surface
pour respirer et se reproduire, même s'il reste dans
l'eau (animal à respiration aérienne
adapté à la plongée et au
déplacement en milieu aquatique), le poisson ne vit
que dans l'eau (animal aquatique); le thon est un animal qui
vit en bancs (ce qui est bien pratique pour les
pêcheurs) alors que le dauphin est plus solitaire,
même s'il peut former des groupes de plusieurs
individus
(caractères
écologiques et éthologiques, voir
physiologiques)
|
Le mode vie de l'animal est-il
fixe, déterminé, ou bien adaptable, modifiable
?
Par quelles éléments
héréditaires est-il transmis ?
|
Des réponses
provisoires:
Si un animal semble adapté à son milieu de vie
et a perdu toute capacité de changer brutalement de
milieu (un poisson marin ne peut pas survivre en eau douce
pour de multiples raisons liées à ses
métabolismes), il existe cependant une certaine
plasticité comportementale qui diffère selon
les espèces et les individus. La diversité
écologique des populations et l'adaptation de
celles-ci reposent plus sur une théorie de
l'évolution que sur une théorie du
vivant étant donné la faible
compréhension que l'on a des liens existant entre le
niveau cellulaire et le niveau comportemental d'un
métazoaire. On a l'habitude de poser comme une
donnée l'adaptation de l'animal au milieu, ce qui est
en quelque sorte que extension de la loi logique
d'optimisation des processus citée plus haut mais
peut aussi être qualifiée de finalisme
(interne): un être vivant est adapté à
son milieu de vie, parce que "vivre est la
finalité interne (le but biologique) de tout
être vivant". (voir finalisme
dans le chapitre précédent)
Notre compréhension de
l'hérédité comportementale repose
à ma connaissance sur des mécanismes
innés, qui seraient en quelque sorte la
conséquence de la structure (anatomie) et du
fonctionnement (physiologie) des organes, et des
mécanismes acquis (plus proprement
éthologiques) par apprentissage individuel ou par
l'exemple de parents ou congénères.
|
Idéalement, il ne faudrait comparer que des individus. Mais
les comparaisons utilisent des caractères. Et les
classifications nés de ces comparaisons classent bien des
individus. D'où le caractère imparfait et provisoire de
toute classification. Nous reviendrons sur ce problème.
Nous allons essayer de creuser un exemple de caractère, les
plumes, qui est propre aux oiseaux actuels.
b. Classer des
Vertébrés à l'aide de caractères
b1. Un exemple de caractère(s): les
plumes
L'article de Pour La Science du n°305 de mars 2003
(Les plumes des dinosaures, Richard Prum et Alan Brush,
p 24-32) est l'occasion rêvée de présenter une
étude évolutive d'un caractère porté par
certains vertébrés: les oiseaux, mais aussi, par des
fossiles.
L'exemple est détaillé dans une page
annexe sur les plumes. Voici un
résumé de ce que de nombreux paléontologistes
pensent aujourd'hui, d'après les articles cités.
b1.1. les plumes actuelles et fossiles sont variées et ont
sans doute de nombreux rôles
Les plumes sont actuellement considérées comme des
structures variées (plumes de duvet, plumes de contour)
dont les rôles sont tout aussi divers: protection
thermique, protection hydrique, vol, nage, reproduction,
camouflage... Ce n'est que depuis une vingtaine d'années que
l'on peut affirmer avec des arguments paléontologiques solides
qu'il en a été de même au cours des temps
géologiques. Les différents types de plumes et les
différents rôles, qui restent toujours des
hypothèses paléontologiques, non
expérimentables, sont probablement apparus progressivement au
cours du temps. Si seuls les oiseaux actuels ont des plumes, tous
les organismes fossiles qui ont porté des plumes
n'étaient pas des oiseaux.
b1.2. les plumes sont apparus dans le groupe des dinosaures
théropodes
Les plumes seraient apparues vers la fin du Jurassique, ou plus
probablement vers le début du Crétacé (vers 120
millions d'années, comme les oiseaux !!!), chez des dinosaures
coureurs, les théropodes. Ces phanères se sont
différenciées (plumes tubulaires non ramifiées,
plumes de duvet, plumes planes) dans ce groupes avec des rôles
variés (difficiles à reconstituer mais dépendant
étroitement du milieu de vie reconstitué pour chacun
des fossiles). Certains de ces dinosaures à plumes sont
à l'origine des oiseaux, qui volent et dont les plumes
interviennent de façon primordiale dans ce moyen de
locomotion.
Du point de vue phylogénétique, le fait de
posséder des plumes est donc un caractère trop
imprécis pour être qualifié d'une façon
évolutive (polarisé, comme disent les cladistes) sans
discussion. On pense qu'il a existé de nombreux types de
plumes avec probablement de nombreux rôles. Si l'on
connaît de nombreux dinosaures à plumes, ils sont pour
l'instant tous classés dans le groupe des Théropodes.
Certains n'ont que du duvet, d'autres de magnifiques plumes planes,
certains même ont des plumes sur leurs quatre membres et leur
queue (Microraptor gui). Certains couraient probablement,
d'autres planaient, d'autres enfin avaient peut-être un vol
battu.
b1.3. les oiseaux prennent naissance vers 120 millions
d'années dans ce groupe de dinosaures théropodes
emplumés
L'origine des oiseaux ne se comprend qu'en regroupant de nombreux
autres caractères essentiels comme la présence d'un
petit nombre de vertèbres caudales (- de 26) ou la
présence d'une fourchette (issue de la soudure des deux
clavicules en un os unique), ou encore le retournement du premier
orteil. Avec un vocabulaire cladiste, on peut affirmer que la
présence de plumes de contour sur les membres
antérieurs est ainsi une homoplasie (analogie au sens
classique) entre un Microraptor gui (dinosaure
théropode du groupe des dromæosauridés, non
directement affilié aux oiseaux) et un Archeopteryx (un
ancêtre direct des oiseaux). La présence de plumes ne
constitue une homologie de filiation qu'au sein de la
lignée des oiseaux (Aves).
b2. Généralisation: les caractères des
vertébrés
Certains en opposant une classification ascendante (qui regroupe
par les ressemblances) et une classification descendante (qui divise
par les différences) ne font que mettre en avant les deux
groupes de références principaux:
l'espèce pour celui qui unit (on regroupe les
espèces en ordres puis classes puis
embranchements puis règnes...) et le monde
vivant pour celui qui sépare (on divise le monde vivant en
règnes puis embranchements puis classes puis ordres puis
espèces...). Par exemple l'état pluricellulaire est un
caractère commun à trois règnes et qui
sépare les eucaryotes du règne des Protistes de tous
les autres eucaryotes.
Par commodité, ressemblances ou différences sont
appelées des caractères mais n'oublions pas
qu'ils sont définis pour un individu (taxon =
élément de la classification) relativement à un
groupe. Comme il est évident que certains
caractères sont de peu d'importance (couleur d'iris) ou de peu
d'intérêt systématique (poids), il nous faut
hiérarchiser ces caractères. Par exemple on peut
distinguer les caractères éthologiques,
embryologiques, morphologiques, physiologiques,
anatomiques, histologiques, moléculaires
...
En taxonimie (science de la classification), il est donc
fréquent de rechercher dans un premier mouvement ce qui unit.
On recherche d'abord des déterminismes communs car
l'unité du vivant est un principe premier. C'est cette
unité qui fonde l'évolution dynamique, un
phénomène qui a commencé dans le temps et qui
se prolonge.
Ensuite on sépare pour classer, pour montrer le second
principe qui est la diversité du vivant , signe de
l'évolution qui s'étend. Plus
précisément il s'agit souvent de montrer
l'évolution diversifiante, ce que l'on peut appeler la
variation.
2. classer des caractères ou bien des organismes, deux
compréhensions différentes de l'évolution
On cherche en vain dans les manuels scolaires une
définition de l'évolution (qui font tous cependant
référence à une diversité née
d'une origine commune, mais sans affirmer clairement:
l'évolution c'est ....), sauf dans celui de
l'éditeur Didier qui affirme: l'évolution, c'est la
modification des organismes au cours des
générations. Le terme modification fait,
à mon avis, presque référence aux
caractères (voir partie
précédente sur le transformisme, qui est le
mot historiquement premier pour désigner l'évolution).
Le question centrale est là:
l'évolution, pour un néodarwinien,
c'est la transformation au cours du temps des
caractères, vus comme l'expression du
génotype;
l'évolution, pour un partisan d'une phylogénie
naturelle, c'est la transformation des organismes vivants,
sans référence à une causalité
particulière.
|
En effet, le terme caractère est employé par
presque tout le monde comme un équivalent de
phénotype, sous le contrôle du génotype.
Il véhicule donc une théorie, néodarwinienne, du
vivant.
Pour le partisan d'une théorie non darwinienne, un
caractère observé chez l'adulte peut très bien
correspondre à la réponse de l'organisme à une
contrainte du milieu. De même, un caractère embryonnaire
majeur peut très bien donner naissance à des structures
variées en fonction des environnements de
développement. Deux caractères différents chez
l'adulte, par exemple, peuvent provenir du même
caractère embryonnaire... Bref, la notion de caractère
chez l'adulte (où chez l'embryon) n'est pas sous la
dépendance d'un déterminisme unique.
Si le caractère désignait un élément
classificatoire univoque (avec un seul sens), cela permettrait
d'établir une phylogénie universelle. Ce qui
était peut-être le rêve du cladiste. Mais il est
clair que la classification cladiste la plus habituellement
présentée participe de la théorie
néodarwinienne de l'évolution.
Pour les chercheurs, cela ne pose pas trop de problèmes, on
peut espérer qu'ils puissent prendre conscience de ces
implicites; mais pour les élèves, il en est autrement.
Leur faire classer des caractères en leur présentant
par exemple la plume des oiseaux comme une écaille
très modifiée (Bordas p 33) est dangereux,
même si ce n'est pas faux, quoique cela reste à
démontrer. Combien d'enseignants prendront-ils la peine (et le
temps !) de préciser que dans le cadre d'une anatomie
comparée, ce sont, non pas les productions finales qui
permettent d'établir l'homologie, mais la
présence de bourgeons épidermiques, leur croissance et
différenciation, dont les caractéristiques constituent
justement la théorie évolutive: sous le contrôle
d'un programme de développement pour les néodarwiniens,
et sous le contrôle d'un automatisme (comportement social
élémentaire de chaque cellule engagée dans une
progression autonome au sein d'une population) et de
réarrangements liés aux interactions entre populations,
pour les partisans d'une théorie
téléologique.
J'avoue avoir passé énormément de temps sur
cette partie sans avoir trouvé de solution satisfaisante. Je
vais cependant m'efforcer de présenter un cladisme non
darwinien... dont l'occasion m'est fournie par l'article sur les
plumes.
1.1 Des ressemblances qui unissent :
homologies et analogies et l'unité du vivant
Intéressons-nous d'abord aux ressemblances ou
similitudes qui font référence à la
stabilité de l'évolution.
La question pourrait aussi être formulée ainsi:
l'évolution se
répète-t-elle ?
La question fondamentale pour des phylogénies est de savoir
si une ressemblance ou similitude :
- est l'expression d'une réelle parenté
(cette ressemblance résulte d'un
phénomène qui n'est apparu qu'une fois dans
l'histoire de la vie et qui depuis se répète
dans une lignée, un groupe d'organismes tous
apparentés phylogénétiquement - que l'on
appelle groupe monophylétique ; on peut dire aussi
que les deux vertébrés ont un ancêtre
commun pour ce caractère. Une telle ressemblance est
qualifiée
d'homologie.
Ex: la présence du squelette
vertébral de tous les vertébrés est un
caractère toujours homologue chez tous les
vertébrés.
- ou si elle est fortuite (non pas dans le sens ou elle
serait le fruit du hasard, car cette ressemblance repose toujours
sur des mécanismes du vivant voisins, si ce n'est
identiques) et donc plutôt secondaire (dans le sens
où cette ressemblance résulte d'un
phénomène qui est apparu plusieurs fois dans
l'histoire de la vie et qui se répète donc
depuis dans plusieurs groupes non apparentés
phylogénétiquement, que l'on qualifie donc de
polyphylétique; on peut dire que les deux
vertébrés qui se ressemblent n'ont pas
d'ancêtre commun pour ce caractère). Une telle
ressemblance est qualifiée
d'analogie.
Ex: la palette natatoire du membre
antérieur du dauphin n'a qu'une analogie de forme avec la
nageoire pectorale du thon. On parle aussi de
convergence.
Le choix ne peut être définitif car il est logique et
comporte des arguments principalement paléontologiques mais
qui dépendent de la théorie évolutive à
laquelle on se réfère. Nous verrons des exemples pour
les Vertébrés par la suite. C'est un travail que l'on
ne peut pas demander à un élève de faire. Mais
vous pouvez avoir à définir ce que sont ces deux types
de caractères.
En phylogénie ont s'efforce de n'utiliser que des
caractères de ressemblance qui traduisent une réelle
parenté, donc des homologies.
Pour répondre à la question de la
répétition de l'évolution on peut donc dire
oui, l'évolution se répète
toujours, c'est même ce qui fait qu'elle est
évolution, dans le temps. Si elle se répète de
façon continue (héréditairement si l'on veut),
la ressemblance issue de cette répétition se trouve au
sein d'un groupe monophylétique et est une homologie. Si elle
se répète à des moments différents de
l'histoire, les ressemblances issus de cette répétition
seront qualifiées d'analogie. Ceci dit deux résultats
identiques n'ont pas forcément la même cause.
remarque 1:
Les cladistes préfèrent le terme d'homoplasie pour
désigner les non homologies et distinguent les analogies
(homoplasies de fonction), la convergence, le parallélisme et
la réversion, qui sont toutes des relations logiques qui ne
seront pas traités ici.
Il est toujours nécessaire de s'assurer d'abord du
monophylétisme d'un groupe (ou de le poser comme
hypothèse !) avant de parler d'homologie au sein de
celui-ci,.
remarques:
2. une comparaison pour les collègues
habitués à l'analyse mendélienne des arbres
généalogiques: il est indispensable de
considérer la récessivité ou la dominance du
caractère avant d'envisager sa liaison à un type de
chromosome; en phylogénie, c'est pareil: il est vain de
vouloir étudier la pertinence de tel caractère
classificatoire dans un groupe dont on a pas auparavant
déterminé le mono ou le polyphylétisme.
3. je corrige aussi les lignes de
l'ancien
cours de TS où je
citais l'article de Corinne Fortin (revue de l'APBG: Classification
et évolution, Biologie-Géologie (Bulletin de l'APBG),
n°3-2000, p 525-537) qui me semble maintenant plus clair: une
homologie s'affirme au sein d'un groupe monophylétique, et
donc entre individus ayant un ancêtre commun.
Ce niveau de ressemblance suffit pour une phylogénie
approximative du vivant mais depuis les années 1980-1990 on
cherché à préciser le vocabulaire, surtout au
sein de l'école cladiste (une des deux méthodes
utilisées en phylogénétique) qui est devenu
très complexe. Le paragraphe qui suit n'en est qu'un
aperçu.
Il est hors de propos de faire ici une
étude complète du cladisme. Ce survol, imposé
par le programme, rend difficile la légitime et indispensable
comparaison avec l'autre méthode
phylogénétique.
1.2 Les différences qui
séparent les ressemblances: les dissimilitudes entre
homologies et la diversité du vivant
Toutes les homologies sont-elles
équivalentes ?
Cela dépend de la méthode
phylogénétique et donc en définitive du sens que
l'on accorde à l'homologie.
En effet une parenté réelle (donc une homologie,
située au sein d'un groupe monophylétique) peut
remonter très loin dans l'histoire de la vie.
* Pour les tenants d'une phylogénie naturelle, toutes les
homologies sont pertinentes pour la phylogénie puisque toute
homologie résulte d'une innovation acquise et
conservée par l'automatisme de l'évolution dans la
lignée.
* Pour les cladistes, une homologie ayant une origine ancienne n'a
pas de valeur classificatoire. Si l'ancêtre commun
remonte à des centaines de millions d'années ce n'est
plus un ancêtre, c'est un mythe. Ils nomment une telle
homologie une homologie primaire (homologie de position
anatomique). Par contre, une homologie affirmée entre deux
taxons (éléments de la classification
phylogénétique) ayant un ANCÊTRE COMMUN DIRECT ou
EXCLUSIF est dite homologie de filiation. Nous
n'utiliserons pas ce vocabulaire, en accord avec le programme.
Ex: tous les membres chiridiens des
tétrapodes (membres dressés dont l'étymologie du
mot (du grec cheiros = la main) rappelle la forme habituelle
à 5 doigts de l'extrémité terminale) sont
homologues selon une homologie primaire. Seules les homologies entre
deux membres chriridiens appartenant à deux taxons
réunis par des liens de filiation directe comme, chez les
équoïdés, Hyracotherium et Mesohippus
(voir ancien
cours de
TS),
sont acceptées par les cladistes comme des homologies de
filiation.
Remarque 4:
Et pourquoi ne pourrait-on pas avoir des caractères
identiques et non seulement des ressemblances ?
C'est avant tout une question de vocabulaire: deux caractères
identiques chez deux taxons différents ne sont pas ...
identiques absolument... puisqu'ils n'appartiennent pas toujours au
même taxon. Comme je l'ai précisé au début
de la partie, la notion de caractère est relative à
un individu ou à un groupe. Les taxons d'un même
groupe partagent le même caractère relativement à
ce groupe mais n'ont pas des caractères identiques.... C'est
tout le problème d'enseigner l'évolution avec des
caractères et non pas seulement avec des théories. On y
gagne en précision mais pas forcément en
compréhension.
Pour les cladistes les homologies de filiation ne sont pas non
plus toutes équivalentes. Ils distinguent entre elles des
différences
Intéressons-nous maintenant aux différences .
Les différences doivent être LÉGÈRES, car
on n'a aucune utilité à comparer deux caractères
qui n'ont aucune similitude; on pourrait plutôt parler de
faible
dissimilitude.
Un caractère est dissemblable d'un autre caractère mais
il n'y a pas de temps, c'est une relation logique (voir la
page sur le
cladisme). Les faibles
dissimilitudes font référence à la
l'évolution diversifiante.
On recherche cette fois encore la parenté mais non plus
dans une ressemblance mais dans une faible dissimilitude (mais pas
dans une différence trop importante -
par exemple la différence entre la symétrie
bilatérale et la symétrie radiaire de deux organismes
n'est pas une différence phylogénétique, car ce
ne serait plus une parenté que l'on rechercherait alors mais
un mécanisme ontogénique beaucoup plus
général).
Si une faible dissimilitude entre deux caractères,
déclarée pertinente pour la construction d'une
phylogénie :
- est considérée comme l'expression d'une
parenté et donc d'une
évolution (transformation d'un
caractère présent chez un ancêtre direct ou
exclusif), on dit que l'un des caractères est
dérivé
par rapport à l'autre (il n'y a pas de temps dans cette
relation logique mais une théorie de l'évolution ou
des données paléontologiques peuvent y mettre le
temps);
Ex: dans la cohorte des Glires (groupe de
Mammifères) les Rongeurs (ordre comprenant les souris,
écureuils et autres cobayes... que l'on nomme aussi les
duplicidentés) portent des incisives doubles (2
incisives par demi-mâchoire) alors que les Lagomorphes
(ordre comprenant les lapins et les lièvres..; que l'on
nomme aussi les simplicidentés) portent des
incisives simples (une seule incisive par demi-mâchoire). Le
nombre d'incisives par demi-mâchoire est pour chaque ordre
un caractère dérivé.
De même les plumes des premiers reptiles à plumes
peuvent être considérées comme un
caractère dérivé des phanères
(productions épidermiques kératinisées) d'un
taxon qui engloberait la lignée aviaire (relative aux
oiseaux) des reptiles dans un groupe
monophylétique.
- ne constitue pas l'expression d'une parenté ni d'une
évolution mais au contraire d'une
stabilité (c'est-à-dire que l'on
considère que le caractère a si peu
évolué qu'il est presque identique chez les
deux taxons; on dit que le nouveau caractère est
primitif (à
l'état primitif) ou
ancestral (dans le même
état que chez les deux taxons).
Ex: c'est le cas de l'absence de canines
aussi bien chez les Rongeurs que chez les Lagomorphes qui est un
caractère des Glires (qui possèdent une formule
dentaire très particulière... ce qui est un
caractère dérivé pour ce taxon).
La présence de plumes dans la lignée des oiseaux est
aussi un caractère primitif pour ce taxon qui n'englobe que
des animaux à plumes.
Ce vocabulaire, issu du cladisme, est déroutant (car
les mots ne sont pas pris dans le sens courant) mais cependant
précis et donc, en ce sens, il présente une
amélioration par rapport à ce qui pouvait se faire
avant le cladisme.
Pour être franc ce vocabulaire a probablement
été développé pour intégrer les
caractères moléculaires dont la parenté est rien
de moins qu'évidente (mais se prête facilement à
un traitement probabiliste): le cladiste réalise, avant tout
essai de représentation (cladogramme), la polarisation des
caractères qu'il utilise, c'est-à-dire qu'il
décide de l'état dérivé ou
ancestral des caractères. Il établit ainsi une matrice
de caractères présentant leur état (0
représente l'état primitif et 1 l'état
dérivé). Pour les molécules qu'il choisit comme
homologues, il procède de la même manière, en
considérant chaque site de mutation possible d'une
chaîne nucléotidique comme un caractère, ou
chaque acide aminé d'une chaîne polypeptidique....
La méthode de polarisation et les postulats évolutifs
qui y sont attachés (liés à la théorie
choisie) dépassent le propos de ce cours mais influent bien
sûr fortement sur les cladogrammes obtenus
(voir page
sur le cladisme). C'est là
que se décident les grandes lignes des
phylogénèses. Ce lieu est inaccessible à notre
niveau, ce qui rend les cladogrammes bien fades. Pour preuve
l'utilisation des logiciels comme phylogène que je me
garderai bien de conseiller à qui veut comprendre
l'évolution.
Le vocabulaire dérivé-primitif a été
retenu dans le programme. Nous l'emploierons donc. Mais suis-je
capable de l'enseigner ? Il y a de nombreux
équivalents et nuances à ces mots que nous
n'emploierons pas comme synapomorphie, apomorphie,
plésiomorphie.... (voir cladisme).
La recherche de caractères dérivés et
primitifs exige toujours, comme dans le cas des homologies et
analogies, à se placer dans un groupe monophylétique
(clade) avant tout travail. (voir
remarque 2 ci-dessus)
Les ressemblances et les différences sont les deux facettes
de la phylogénie établies à l'aide de
caractères. Le cladisme a eu le mérite de clarifier
fortement le vocabulaire des phylogénies qui sont donc
devenues plus scientifiques. Mais il a aussi introduit des
postulats logiques qui peuvent occulter la recherche d'une
phylogénie universelle. Je ne détaillerai pas cette
idée dans ce cours mais j'ai essayé de le faire dans la
page annexe sur le cladisme pour les
esprits curieux (les opinions qui y sont exprimées n'engagent
que moi).
2. deux phylogénies et une histoire des
vertébrés
Si les deux classifications phylogénétiques qui
suivent, reposent chacune sur une théorie évolutive,
elles ne sont bien sûr pas définitives ou
certaines, scientifiquement et paléontologiquement. Une
seule est "calculable" au sens d'une réduction à un
algorithme, c'est la première (du moins dans un premier temps
car l'optimisation se finit presque toujours "à la main", sur
le papier ; et d'autant moins que la première étape,
celle de la polarisation des caractères, n'a rien de
mathématique). Une seule est réelle ou
naturelle, c'est la seconde. Philosophiquement, la
première est relativiste (à mon
avis, voir la page
sur le cladisme), la seconde est plus
réaliste mais ouverte aussi bien à un
matérialisme qu'à un vitalisme (voir
page sur l'évolution
téléologique).
La première, cladiste,
construite à partir des caractères, n'est que logique,
elle ne présente pas des organismes qui auraient existé
(même pas virtuellement contrairement à ce que disent
certains exégètes du cladisme, car il faut, avec les
cladistes et de façon claire, affirmer que les ancêtres
des classifications cladistes n'existent pas et n'ont pas
existé). Une classification cladiste REPRÉSENTE les
RELATIONS LOGIQUES que l'on considère comme LES PLUS PROBABLES
(réfutables en fonction des découvertes de la
paléontologie ou d'un consensus sur les relations logiques les
plus pertinentes) qui auraient donné naissance aux INDIVIDUS
ACTUELS OU FOSSILES placés à l'extrémité
des rameaux de l'arbre cladiste ou cladogramme, seuls RÉELS et
seuls représentés. Le fait que les ancêtres ne
sont pas représentés dans les classifications cladistes
ne veut pas dire qu'ils n'aient pas existé, mais seulement
qu'ils sont inaccessibles au cladiste. Les dessins
présentant des intermédiaires paléontologiques
ne cadrent pas avec des classifications cladistes. Pour pouvoir
cependant prévoir des améliorations de leurs
classifications ils ont introduit la notion d'intermédiaire
structural tout aussi théorique et placé au bout
d'une branche quelque part entre deux nœuds, représentant
deux innovations. En fait cet intermédiaire structural n'est
que la représentation d'espèces qui ne rentrent pas
dans l'arbre présenté avec les choix logiques
réalisés et qui, provisoirement sont placés sur
un calque, hors de la classification, le plus près possible de
la position que le cladiste souhaite lui donner .
La seconde,
téléologique,
construite à partir d'une théorie évolutive, se
veut NATURELLE et donc représenter UNE HISTOIRE
(voir partie
1 de ce cours). Elle prétend
représenter réellement les ANCÊTRES successifs
que l'on peut donc rechercher dans les archives
PALÉONTOLOGIQUES (elle se soumet aux découvertes de la
paléontologie et est donc réfutable), même si
leur existence a été éphémère.
Elle les relie au moyen d'une théorie et donc aussi des
relations logiques, tout aussi scientifiques que dans le
cladisme. (Pour mieux appréhender ce qu'est
une phylogénie je renvoie à la partie précédente
de cette page).
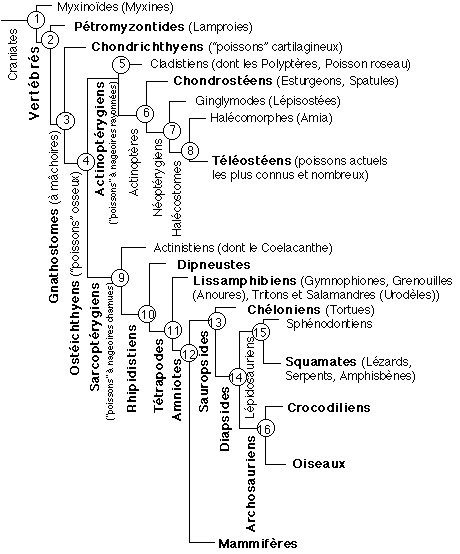
2.1 Un cladogramme SIMPLIFIÉ et illustré des
vertébrés
|
|
|

myxine
|
|

spatule
|
|
|
|

cœlacanthe
|

gymnophione
|

triton
|

tortue
|
|

lézard
|
|

crocodile
|

archeoptéryx
|

autruche
|
|
|
|
|
|
|
|
|
|
|
Un cladogramme des
Vertébrés:
il se lit de la façon suivante: les Craniates
possèdent tous un caractère
dérivé exclusif qui est la présence
d'un crâne osseux; ce clade (groupe
monophylétique) est composé de deux clades:
les Vertébrés (dont le caractère
primitif exclusif est la possession de vertébres mais
dont la possession d'un crâne est un caractère
primitif) et les Myxinoïdes (qui
possèdent un crâne, ce qui leur est un
caractère primitif mais qui présentent
d'autres caractères propres (dérivés)
comme leur appareil branchial, particulier). Les
vertébrés sont à leur tour
composés de deux clades: les
Pétromyzontides (qui possèdent donc
comme caractère primitif (ancestral) un crâne
et des vertébres mais comme caractère
dérive exclusif par exemple leur bouche en forme de
ventouse) et les Gnathostomes (qui possèdent
comme caractères primitifs leur crâne, leurs
vertèbres et comme caractère
dérivé exclusif leurs mâchoires); ces
derniers sont constitués à leur tour de deux
clades: les Chondrichtyens et les
Ostéichthyens... et ainsi de suite.
(modifié d'après G. Lecointre,
La construction de phylogénies,
Biologie-Géologie (Bulletin de l'APBG), 1-1995,
pp110-111-128 (les nœuds 12, 13 et 14 ont
été modifiés)
et Classification phylogénétique du
vivant, G. Lecointre et H. Le Guyader, Belin,
2001)
|
Quelques
caractères polarisés
considérés comme
dérivés exclusifs
d'après G.
Lecointre et H. Le Guyader dans Classification
phylogénétique du vivant, Belin,
2001
|
|
n° du nœud
|
pour les clades situés en
dessous du nœud dont le numéro est
référencé (dont le nom est noté
en parenthèses)
|
pour les clades situés au-dessus
du nœud dont le numéro est
référence (dont le nom est noté en
parenthèses)
|
|
On notera que étant
donné qu'un cladogramme n'est jamais fixé au
niveau des nœuds qui sont des points de rotation, la
présentation qui en est faite ici n'est qu'une
variante. De plus la polarisation des caractères
n'est pas discutée. Les choix sont ceux des auteurs
cités.
|
|
1
|
(Vertébrés) la présence de
vertébres, de 2 canaux semi-circulaires dans
l'oreille interne, une musculature extrinsèque de
l'œil, une régulation nerveuse du cœur,
rate et pancréas individualisés
|
(Mixynoïdes) appareil branchial très
particulier (poches en forme d'oignons), tête
terminée par un orifice nasopharyngien unique
s'ouvrant dans le pharynx, six tentacules tactiles entourant
la bouche, présence d'un conduit
œsophago-cutané gauche, appareil lingual
|
|
2
|
(Gnathostomes) la présence de 2
mâchoires (du grec gnathos = mâchoire),
squelette branchial interne par rapport aux branchies,
troisième canal semi-circulaire, axones
myélinisés, molécule
d'hémoglobine formée de 4 sous-unités
identiques deux à deux
|
(Pétromyzontides) squelette branchail
particulier (corbeille branchiale), organe olfactif
s'ouvrant dans un tube fermé à une
extrémité, bouche entourée d'une
ventouse
|
|
3
|
(Ostéichthyens) os enchondral,
éléments osseux dermiques, ceinture scapulaire
portant aussi des os dermiques, arcs branchiaux 1 et 2
articulés sur une même pièce osseuse,
sacs aériens connectés au tube digestif
|
(Chondrychthiens) couche de cartilage
calcifié prismatique tapissant le cartilage du
squelette, organes spéciaux d'accouplement chez les
mâles
|
|
4
|
(Sarcoptérygiens) squelette interne des
nageoires monobasal (s'attache aux ceintures par un seul
élément), émail vrai sur les dents, le
5ème et dernier arc branchial s'attache ventralement
sur l'avant-dernier
|
(Actinoptérygiens) chaque dent porte un
petit capuchon de tissu minéralisé
(l'acrodine), perte de la nageoire dorsale
antérieure, écailles articulées
|
|
5
|
(Actinoptères)
|
(Cladistiens)
|
|
6
|
(Néoptérygiens)
|
(Chondrostéens)
|
|
7
|
(Halécostomes)
|
(Ginglymodes)
|
|
8
|
(Téléostéens)
|
(Halécomorphes)
|
|
9
|
(Rhipidistiens)
|
(Actinistiens)
|
|
10
|
(Tétrapodes)
|
(Dipneustes)
|
|
11
|
(Amniotes)
|
(Lissamphibiens)
|
|
12
|
(Mammifères)
|
(Sauropsides)
|
|
13
|
(Diapsides)
|
(Chéloniens)
|
|
14
|
(Lépidosauriens)
|
(Archosauriens)
|
|
15
|
(Squamates)
|
(Sphénodontiens)
|
Quelques commentaires:
* c'est nouveau, cela fait pas mal de noms à apprendre
* les noms des clades reprennent des noms de classes, d'ordres, et
autres divisions classiques, parfois modifiés mais surtout
DANS UN AUTRE SENS (les clades définis ne contiennent pas les
mêmes organismes que les divisions du même nom )....
comment espérer une future unité ?
* on voit aisément le côté provisoire d'une telle
classification; il suffit de changer un caractère
dérivé principal pour changer un nœud et donc tout
l'arbre: les comparaisons entre arbres sont aussi rendues plus
difficiles étant donné la rotation possible au niveau
de chaque nœud.
* la question la plus pertinente concerne plutôt
l'évolution: où est représentée
l'évolution ? Elle ne l'est pas, c'est une classification qui
respecte l'évolution possible mais qui ne la représente
pas. LE TEMPS NE DOIT PAS ÊTRE REPRÉSENTÉ : il
n'existe pas, sauf si l'on s'efforce de classer des fossiles selon
leur âge déterminé par la paléontologie.
Ce n'est pas le cladisme qui peut déterminer un âge. Par
contre il est évident qu'un âge peut être
opposé ou au contraire aller dans le même sens qu'une
relation logique présentée par l'arbre.
* contrairement à ce que les cladistes affirment, certains
caractères sont définis négativement (absence
d'une nageoire par exemple)...
* comme toute classification débutante elle est très
incomplète et donc il nous est impossible de comparer cette
classification (seule accessible aisément aux enseignants du
secondaire) avec celles que nous avons dans des ouvrages classiques
comme le Précis de zoologie de P.P. Grassé. Comment
utiliser des fossiles avec ces classifications incomplètes,
sauf à prendre bien sûr pour argent comptant ce que nous
disent les cladistes.... ; par exemple, la classe des Agnathes
correspond peut-être aux deux clades Myxinoïdes et
Petromizontides; cette classe était d'une grande richesse dans
la classification "classique": les Céphalapsidomorphes
comprenaient les Osteostracés et les Anapsides en plus des
actuels Pétromizonoïdes; venaient ensuite les
Ptéraspidomorphes avec les Hétérostacés
et les Myxinoïdes; enfin les Chélodontes... où
sont-ils tous passés, ont-ils tous été
utilisés dans les classifications cladistes ? Cette
classification incomplète est frustrante. Je souligne que dans
le cas d'un cladogramme le fait qu'une classification soit
incomplète prend une toute autre importance que dans une
classification naturelle. Ce sont les caractères qui
définissent pour le cladiste les groupes qui peuvent
être profondément modifiés avec un seul fossile
original alors que le "phylogéniste naturel" se contente de
chercher le groupe où faire entrer son fossile et, à
défaut, il ajoute une division.
* il n'y a pas que les cladistes pour être capables de discuter
de la position systématique des oiseaux.
2.2 Une phylogénèse naturelle des
vertébrés
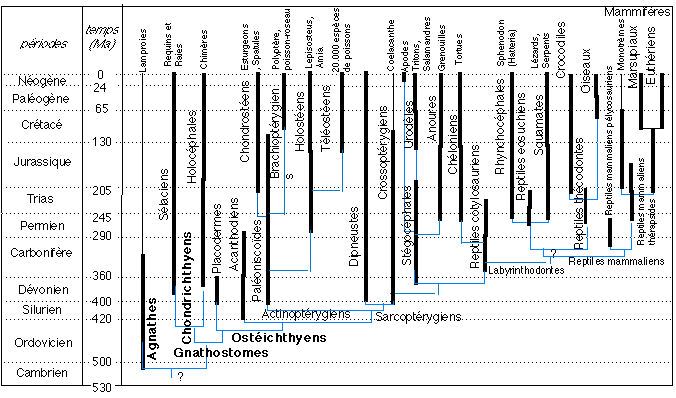
Une phylogénèse des vertébrés
(d'après Biologie animale, , les
Cordés, anatomie comparée des
Vertébrés, A. Beaumont et P. Cassier, Dunod
Université, 1987, p 71, modifié)
Parentés: les caractères communs des
vertébrés, les classes de vertébrés, le
problème du passage à la classe
3. l'origine des
vertébrés

