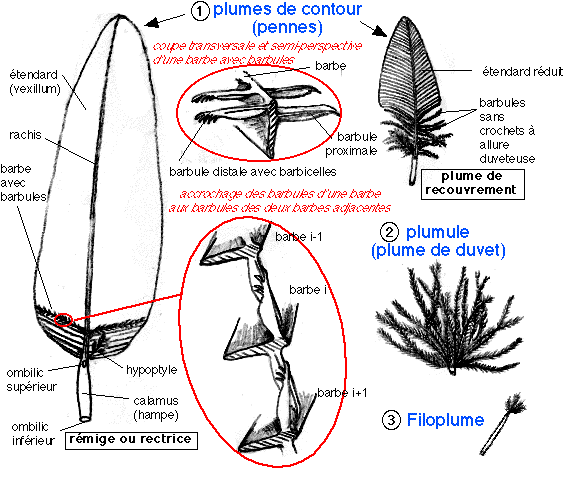
Plumes d'un oiseau adulte actuel (Dessins repris principalement de Beaumont et Cassier)
Il existe trois types de plumes chez l'oiseau adulte:
* les pennes ou plumes de contour, qui selon leur
position sont nommées rémiges (plumes de contour
des ailes), rectrices (plumes de contour de la queue) ou
plumes de couverture. Elles comportent toutes un axe rigide,
partagé en calamus (ou hampe, cylindre creux dont la
cavité est cloisonnée transversalement) proximal et
rachis (plein mais creusé d'un sillon ventral) distal,
ce dernier supportant l'étendard ou vexillum,
composé de barbes (lames parallèles) portant
deux rangées de barbules, les barbules proximales d'une
barbe s'entrecroisant avec les barbules distales de la barbe voisine,
grâce aux barbicelles, petites excroissances crochues
des barbules distales (les barbules proximales sont lisses).
Rémiges et rectrices portent des barbules accrochées
étroitement sur toute leur surface de l'étendard alors
que les plumes de couverture non portantes présentent des
barbes relâchées sur les parties recouvertes par
l'empilement des plumes sur le corps de l'oiseau: les barbules sont
plus longues et sans crochets, à allure duveteuse. La partie
ventrale et supérieure du calamus porte un étendard
secondaire duveteux, plus ou moins développé, chez un
même oiseau, selon les parties du corps et chez les
différents oiseaux.
*les plumules ou plumes de duvet sont de structure
remarquablement uniforme chez tous les oiseaux. Une touffe de barbes
s'insérant sur un court calamus (rachis réduit ou
absent). Chaque barbe porte deux rangées de barbules lisses,
toujours plus développés vers la base que vers le
sommet. Le duvet, très abondant chez les oiseaux aquatiques,
couvre le corps, sous la couverture des plumes de contour. Son
rôle dans l'isolation thermique est essentiel.
* et les filoplumes, qui sont réduites à un
rachis filiforme, portant parfois quelques barbes à son
extrémité. Elles sont situées à la base
des plumes de contour, se formant sur la paroi latérale de
leur follicule (voir ci-dessous). Ce sont les filoplumes qui restent
souvent sur les volailles plumées et que le cuisinier doit
flamber.
La répartition des plumes varie selon les espèces: absence de duvet chez les Ratites (Autruches, Casoars, Émeus, Nandous et Kiwis), très grandes rectrices lâchement groupées chez les autruches (servant à la confection de plumeaux ou de plumes à chapeau...), absence de rectrices chez les Nandous, réduction des rémiges et rectrices chez les Casoars et Émeus, uniques plumes de revêtement longues et étroites chez le Kiwi, très grand développement des 10 rémiges primaires des apodes (Martinets et Colibris) qui en font d'excellents voiliers, extraordinaire développement des rectrices du mâle aussi bien chez les oiseaux-lyres que le paradisier ou encore le paon, plumage très particulier des impennés (Manchots) avec notamment des rectrices transformées en baguettes cornées et courtes...
Ainsi on peut interpréter la structure et la localisation
des plumes comme concourant à la réalisation de
plusieurs fonctions:
- des plumes pour voler: rémiges portantes, rectrices
stabilisatrices, plumes de contour facilitant la
pénétration dans l'air... l'isolation thermique et de
l'humidité est aussi un élément important du
vol
- des plumes pour isoler de l'eau : du fait des
sécrétions huileuses associées qui en font une
couverture hydrofuge (qui préserve de l'eau)...
- des plumes pour isoler thermiquement : le duvet des oiseaux
aquatiques en est un exemple démonstratif, c'est l'air
emprisonné entre les barbes et barbules qui sert de matelas
isolant
- des plumes pour nager: du fait de leurs
propriétés d'isolants thermiques, hydrofuges et aussi
de la flottabilité due au matelas d'air emprisonné
entre les plumes; sans oublier de profondes modifications de
structure et de formes comme les petites plumes écailleuses
recouvrant les ailes nageuses des impennés (Manchots)...
- des plumes décoratives qui interviennent dans la
recherche et le choix des partenaires reproducteurs...Les
pigments de la plume se déposent lors de la formation des
crêtes des barbes. Ils sont noirs (mélanines), jaunes
(porphyrines) ou jaunes-rouges (caroténoïdes). Les
couleurs vertes ou bleues sont parfois dues à des pigments
spéciaux mais plus souvent à des interférences
(la lumière est décomposée lors du passage
à travers les minces lamelles de kératine des barbules:
on sait par exemple que les barbules des rectrices du Paon (Pavo)
sont limitées par 3 couches fines (0,4 µm) de
kératine). L'accumulation des pigments varie avec l'âge
de l'oiseau.
Les plumes sont des phanères, c'est-à-dire
des productions épidermiques fortement
kératinisées.
« Une plume naît d'un bourgeon épidermique
que soulève une papille dermique vascularisée.
Le bourgeon s'allonge en un cylindre épidermique oblique
entourant un axe dermique ou pulpe, puis s'enfonce
progressivement par sa base sous la surface de la peau,
entraînant l'épiderme qui s'invagine en un follicule
plumaire.»
Par comparaison un poil, caractéristique des
Mammifères, est un filament kératinisé issu de
l'enfoncement d'un bourgeon épidermique dans le derme,
entourant une papille dermique pulpeuse à sa base.
De même, une écaille d'Amniote est un
épaississement corné de l'épiderme (elles
existent notamment sur les pattes des oiseaux et sur la queue ou les
flancs de divers mammifères comme la Tatou) .
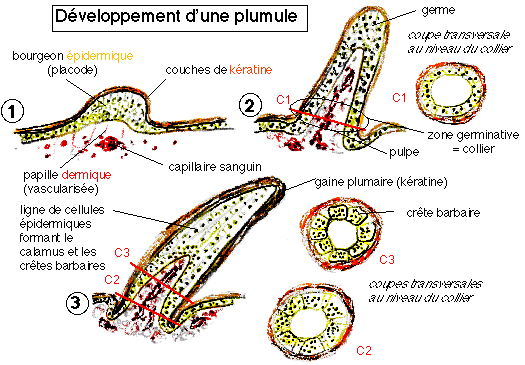
« Au sein de l'épiderme épaissi de la partie apicale du bourgeon, des files longitudinales de cellules poussent à partir d'une zone germinative annulaire ou collier, se vacuolisent, se kératinisent et constituent les crêtes barbaires. Des crêtes barbulaires se différencient de la même façon de part et d'autre des crêtes barbaires. L'ensemble de l'ébauche plumaire est revêtu d'une gaine kératinisée, différenciée à la périphérie du bourgeon épidermique. »
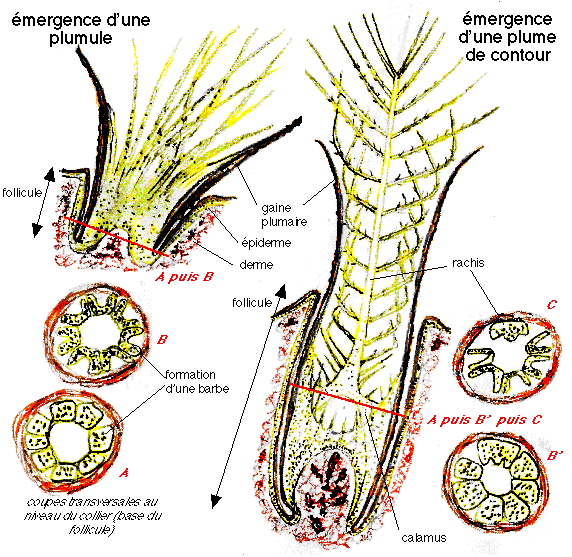
« L'évolution de l'ébauche est simple dans le
cas du duvet, complexe dans le cas d'une plume de contour:
- dans le cas d'une plume de duvet, les crêtes barbaires
sont disposées régulièrement autour du collier
qui engendre un court calamus kératinisé;
- dans le cas d'une plume de contour, le collier
prolifère dans sa région dorsale et s'allonge en une
tige pleine, ébauche du rachis, qui entraîne les
crêtes barbaires à son extrémité. De
nouvelles crêtes barbaires se différencient à la
face ventrale du collier et, subissant le même
déplacement, viennent s'ajouter en arrière des
précédentes le long du rachis qui s'allonge
régulièrement. Au cours de cette évolution, les
crêtes barbaires initialement insérées
verticalement sur le collier s'insèrent transversalement sur
le rachis, en demi-cercles légèrement obliques qui
s'affrontent par leurs extrémités le long d'une ligne
médio-ventrale. Quand le cylindre plumaire émerge
à la surface de la peau, la gaine plumaire se déchire
et le cylindre se fend selon cette ligne médio-ventrale,
libérant les barbes qui s'étalent de chaque
côté du rachis en un étendard. A la face ventrale
du collier un mécanisme identique met en place un
étendard secondaire (hypoptyle). Quand l'étendard
principal est achevé, le collier prolifère
régulièrement et édifie un cylindre de
kératine dure, le calamus, ouvert à ses deux
extrémités.»
Du point de vue moléculaire, les données ne sont pas très vulgarisées et seul l'article cité au début de cette page (Les plumes des dinosaures, Richard Prum et Alan Brush, Pour La Science , 305, mars 2003, p 24-32) y fait directement allusion. L'un des auteurs a particulièrement étudié l'expression de deux gènes: Shh (sonic hedgehog) et Bmp2 (Bone Morphogenetic Protein 2) au cours du développement des plumes. Il a mis en évidence que les protéines Shh et Bmp2, résultant de l'expression de ces gènes, étaient produites à des taux variés et spécifiques lors de toutes les étapes et dans toutes les ébauches plumaires. Son hypothèse de départ était que les plumes modernes, portées par un oiseau actuel, reflètent les différents stades de développement, qui pouvaient être séparés chez des organismes fossiles. Il faut donc que du point de vue moléculaire, chaque stade puisse correspondre à une étape, résultant du stade précédent et dépassée dans le stade suivant. Il semblerait que ses résultats puissent s'interpréter dans cette optique, mais l'article est un peu court à ce sujet.
Au cours de sa vie, l'oiseau porte 3 types de plumages:
- le duvet (duvet néoptile ou plumage du poussin,
porté quelques semaines habituellement),
- le plumage juvénile (plumes ne différent
guère de celles de l'adulte si ce n'est par la taille, la
forme, la pigmentation, la structure plus lâche due à un
nombre de barbes plus faible),
- et, après la mue juvénile, rarement totale, le
plumage adulte (plumage définitif progressivement
atteint par des mues successives). C'est ce dernier qui
présente souvent un dimorphisme sexuel et parfois un
dimorphisme saisonnier (très marqué par exemple chez le
vanneau ou les cormorans).
La mue assure le renouvellement périodique
annuel (lors du repos sexuel, après la reproduction) ou
bisannuel (mue supplémentaire juste avant la
période de reproduction) du plumage. La plume ancienne se
décolle de sa gaine et tombe. Elle est remplacée par
une nouvelle plume qui peut parfois pousser en s'engageant dans le
calamus de l'ancienne plume (Casoars, Émeus, Manchots). La
formation d'une plume résulte toujours de la reprise
d'activité d'une papille plumaire ancienne dont le nombre et
la disposition sont donc définitifs chez l'adulte. Les
papilles plumaires sont probablement toutes formées chez
l'embryon. Le déterminisme de la mue n'est pas encore
complètement élucidé mais fait intervenir
l'hormone thyroïdienne, l'hypophyse et le système
nerveux.
La mue est rarement incapacitante. Étant rarement
totale et toujours progressive, l'oiseau n'est pas privé de
ses capacités de vol, sauf exceptions: Canards, Oies, Cygnes,
Flamants, perdant brusquement toutes leurs rémiges et
étant incapables de voler pendant plusieurs jours. Font aussi
exception, les Manchots, très amaigris lors de la mue, leurs
pertes thermiques augmentant fortement, et n'allant pratiquement plus
nager, du fait qu'ils "se mouillent".
Les plumes ne sont pas disposées uniformément
à la surface du corps sauf chez les Ratites (qui ont cependant
de larges surfaces dénudées) et les Manchots. Les
plumes s'insèrent sur des aires nommées
ptérylies, séparées par des zones nues ou
duveteuses, les aptéries.
On dénombre entre 2.550 et 2.700 plumes en février chez
le Bruant à gorge blanche (Zonotrichia albicollis) et
seulement 1500 à 1550 en octobre. Un Colibri (Archilochus
colubris) n'en porte que 940 (en juin) alors que le Cygne
américain (Cygnus columbianus), nettement plus gros, en
porte 25.216 (en octobre). Le poids du plumage est très
élevé comparativement à la masse corporelle: par
exemple il représente le double de celui du squelette (soit
14% de la masse corporelle) chez le Pygargue chauve
(Haliaëtus leucocephalus).
La première empreinte dans les calcaires lithographiques du Jurassique supérieur de Bavière attribuée à une plume isolée a été rapportée en 1861 sous le nom d'Archaeopteryx lithographica (on donne en effet à tout fossile un nom de genre et d'espèce dans la nomenclature binominale, même si l'on ne dispose que d'une plume; d'ailleurs cette bonne habitude trouve sa justification dans les trouvailles qui suivirent). Cette même année, un squelette de cet animal est découvert à proximité. Depuis lors, on connaît six autres squelettes de cet oiseau primitif provenant tous de cette région d'Allemagne. Le dernier en date, découvert en 1992, a été décrit en 1993 comme une nouvelle espèce, Archaeopteryx bavarica.
La découverte de Sinosauropteryx prima en
Chine (formation de Yixian, dans le nord-ouest de la Chine) au cours
de l'été 1996 (Un dinosaure à
plumes ?, Echos de la science, La Recherche, 293,
décembre 1996, p 16) dans des sédiments
lacustres datés alors d'environ 140 millions d'années
(Jurassique terminal) ou de 128 à 124 millions d'années
selon des datations plus récentes (donc Crétacé
inférieur), a fait sensation. On connaît deux
exemplaires de ce petit dinosaure carnivore (Théropode,
cœlurosaure), bipède, à longue queue, long d'un
mètre (mais dont le corps a la taille, ou plutôt le
volume, de celui d'un poulet) présentant des traces de
phanères évoquant un duvet (5 mm) sur la tête et
le cou, le long du dos et de la queue (recouvrant probablement tout
le corps). Dans la même formation on a trouvé des
mammifères placentaires, des oiseaux anciens (possédant
des empreintes de phanères que l'on peut rapporter à
des plumes de contour), des plantes à fleur et de nombreux
autres dinosaures théropodes. Parmi les oiseaux anciens on
peut citer Caudipteryx, de la taille d'une dinde,
présentant des plumes de contour à
l'extrémité de ses membres antérieurs. D'autres
fossiles de dinosaures à plumes comme ceux de
théropodes dromæosaures Microraptor et
Sinornithosaurus ont été trouvés
avec ce que l'on peut interpréter comme des plumes de
contour.
Un de ces Dromaeosauridés nommé Microraptor gui
et récemment découvert dans la formation de
Liaoning datée du Crétacé inférieur (120
millions d'années), possède même des plumes de
contour non seulement le long des membres antérieurs mais
aussi le long des membres postérieurs et de la queue. Un
dinosaures à 4 ailes en quelques sorte.
 |
de Microraptor gui et reconstitutions avec silhouette classique (en haut) et silhouette en vol plané (en bas) (in Les plumes des dinosaures, Richard Prum et Alan Brush, Pour La Science , 305, mars 2003, p 32, modifié) |
Ce gisement chinois comporte aussi des dizaines d'oiseaux primitifs dont le plus fréquent est Confuciusornis sanctus. Cet oiseau, de la taille d'une pie, présente encore bien des ressemblances avec Archaeopteryx , mais il possède une queue beaucoup plus courte, et des mâchoires dépourvues de dents. Des oiseaux plus modernes, appartenant à la sous-classe des Ornithurae, qui comprend aussi les oiseaux actuels, sont aussi présents dans les gisements chinois. Liaoningornis , de la taille d'un moineau, est le plus ancien oiseau connu à posséder un bréchet sur lequel pouvaient s'insérer de puissants muscles servant à mouvoir les ailes. Chaoyangia est un autre oiseau chinois présentant des caractères modernes. La présence de ces formes assez évoluées aux côtés de représentants du groupe plus archaïque des Énantiornithes, comme Sinornis et Cathayornis , montre que les oiseaux s'étaient notablement diversifiés dès le début du Crétacé.
Les calcaires lacustres de Las Hoyas (près de Cuenca, centre de l'Espagne), un peu plus récents (environ 115 Ma) que les sites chinois, ont livré les restes d'Iberomesornis , Concornis et Eoalulavis hoyasi , tous attribuables aux Énantiornithes. Les traces de plumage bien conservées chez Eoalulavis hoyasi montrent que cet oiseau possédait une alule, plume insérée sur le premier doigt de l'aile, qui chez les oiseaux actuels joue un rôle important dans le vol en améliorant la stabilité et en facilitant les manœuvres.
10/2010 ("Des plumes toujours plus anciennes, questions à
Ronan Allain, La Recherche, 446, novembre 2010, 20-21)
Concavenator corcovatus est un dinosaure carnivore du groupe
des allosaures. L'os de l'avant-bras porte un
alignement de petites protubérances qui ressemblent aux points
d'attache des plumes chez certains oiseaux actuels. On pense donc que
ce petit dinosaure très primitif portait des plumes sur ses
deux avant-bras (F. Ortega et al., Nature, 467, 203, 2010).
Du point de vue du squelette Sinosauropterix est proche de
Compsognathus, un autre petit Théropode fossile du
Jurassique supérieur d'Europe et semble éloigné
phylogénétiquement de l'origine des oiseaux. Les
plumes pourraient donc être un caractère apparu
chez d'autres dinosaures indépendamment de la lignée
des oiseaux.
De façon similaire, Caudipteryx, avec ses plumes de
contour à l'extrémité des membres, est
considéré comme un dinosaure coureur, de la
lignée des oiseaux. De même, les
Sinornithosaurus, toujours de cette lignée, ont des
plumes de contour qui peuvent difficilement être
rapportées au vol. Les plumes de contour pourraient donc
être apparues dans la lignée des oiseaux, sans
être liées directement à la fonction du vol.
On a proposé dernièrement que les jeunes poussins
d'oiseaux primitifs duveteux se réfugient sur des pentes et
faisant ainsi leur apprentissage d'un vol de fuite,
intermédiaire entre leur vol couru et le vol plané
(Une troisième façon de voler,
Philippe Taquet, La Recherche, 362, mars 2003, p
14).
Mais il y a encore plus fort, - et cette hypothèse invalide la
précédente - Microraptor gui , cité plus
haut, est un dinosaure qui planait probablement. Or l'Archeopteryx
volait déjà probablement à cette
époque. Le vol plané serait donc apparu
indépendamment chez plusieurs dinosaures à
plumes.
En effet, on a longtemps considéré la plume comme une
phanère dont le rôle principal était le vol. Le
fait de posséder des plumes pouvaient donc être
interprété comme une homologie reposant sur une
même origine embryonnaire et une même fonction.
L'origine des plumes a longtemps été rapporté
aux écailles. Le raisonnement reposait probablement plus sur
l'origine reptilienne des oiseaux, les caractères des oiseaux
devant donc être des caractères reptiliens
transformés. Cette vision est totalement abandonnée. Le
développement des plumes peut être embryologiquement
séparé de celui des écailles des amniotes,
même si ce sont évidemment des phanères de
vertébrés. Dans leur article (Les
plumes des dinosaures, Richard Prum et Alan Brush, Pour La
Science , 305, mars 2003, p 24-32) Prum et Brush proposent
une évolution des plumes en 5 stades illustrés
ci-dessous:
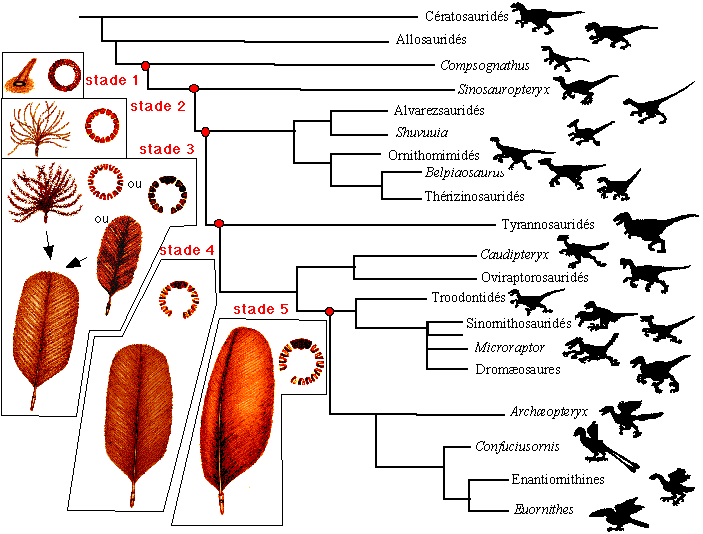
|
stade 1 |
plume cylindrique creuse, collier folliculaire simple |
Sinosauropteryx |
|
stade 2 |
apparition de barbes non ramifiées attachées au calamus, collier folliculaire avec crêtes barbaires différenciées |
Alvarezsauridés, Shuvuuia, Ornithomimidés, Belpiaosaurus , Thérizinosauridés |
|
stade 3 |
deux chemins évolutifs sont proposés: l'apparition de plumes avec barbes non ramifiées fusionnant sur un rachis central ou apparition de barbes ramifiées (avec barbules) rattachées à leur base à un calamus court: les deux hypothèses conduisant à des plumes planes avec des barbes ramifiées. |
Tyrannosauridés |
|
stade 4 |
apparition de barbules qui s'accrochent sur les barbules adjacentes ce qui resserre le vexillum |
Caudipteryx, Oviraptorosauridés, Troodontidés, Sinornithosauridés, Microraptor , Dromæosaures |
|
stade 5 |
les plumes deviennent assymétriques et ressemblent aux plumes modernes |
Archæopteryx, Confuciusornis, Enantiornithines, Euornithes |
En résumé, on peut dire qu'à l'heure actuelle, on considère que la plume n'est pas une structure unique mais qu'il existe de nombreuses plumes pour différentes fonctions. Les homologies anciennes sont devenues des analogies, au sens habituel. Avec le vocabulaire cladiste, plus précis, les plumes ayant existé chez les différents groupes de dinosaures emplumés non directement apparentés sont des homoplasies car c'est ainsi que l'on nomme les ressemblances entre caractères n'ayant pas un ancêtre commun direct (voir cladisme). Dans le cas où l'on estime que ces plumes avaient le même rôle on peut parler de convergence, qui sont des cas particuliers d'homoplasies.
Deux lectures de ces données sont possibles.
* un néodarwinien y voit
des innovations résultant de mutations apparues au
hasard (dans des gènes complexes dont les Shh et
Bmp2 ne sont qu'un tout petit échantillon) et de
sélections naturelles (avantage du à la
présence de plumes, favorisant la survie des dinosaures
emplumés qui transmettent de façon plus efficace leurs
gènes...) mémorisées et transmis
dans l'ADN.
La faiblesse de ce modèle vient du fossé entre le
niveau moléculaire et le niveau histologique et anatomique. En
effet, les quelques gènes qui ont été reconnus
comme impliqués dans le développement des
phanères, aussi bien des poils que des plumes, l'ont
été aussi dans le développement du squelette; ce
qui met plutôt l'accent sur l'unité des
mécanismes moléculaires que sur leurs
différences. Le fonctionnement de ces gènes est plus
difficile à impliquer dans l'émergence de plumes, de la
différenciation des différents types de plumes, ou de
la localisation de ces plumes.
* un finaliste, partisan d'un
modèle téléologique, voit dans ces
innovations des expressions de l'émergence d'une nouvelle
structure de type phanère (type plume) dont l'apparition
est rendue possible par la variation existant au sein des
interactions entre populations embryonnaires (tout comme les
interactions entre épiderme et derme peuvent conduire aux
écailles ou aux poils), mémorisée dans,
et transmis par, le cytoplasme de l'ovocyte. Une fois
l'évolution engagée dans la réalisation de cette
structure, les organismes qui se la transmettent, ne peuvent qu'en
explorer les variations (par des décalages entre progressions
autonomes de populations mais aussi par des mutations... voir la page
sur une autre théorie de
l'évolution). Le mécanisme embryonnaire principal,
qui différencie les plumes des autres phanères, se
situant probablement dans le stade précoce où la
placode épidermique se soulève. Une recherche
menée dans ce cadre reste à réaliser mais on ne
peut s'empêcher de penser que cette vision du
développement est plus proche d'une interprétation
évolutive, comme celle présentée ci-dessus par
Prum et Brush, que ne l'est le modèle darwinien.
Pour résumer cette vision de l'évolution en quelques
lignes:
Les dinosaures portaient habituellement des écailles, mais
certains ont pu porter en plus ce que l'on a appelé des
plumes. Les populations cellulaires donnant les écailles et
les plumes (et aussi les poils) sont les mêmes. Ce sont les
interactions entre ces populations et les produits
synthétisés par ces cellules qui déterminent
l'émergence d'une structure tubulaire plutôt que la
croissance d'une écaille cornée plane à la
surface d'une papille . Une fois que chez UN organisme on a vu
apparaître cette nouveauté (stade 1 du modèle de
Prum et Brush), elle s'est transmise à tous ces descendants
(qui ne sont pas tous des oiseaux mais dont les oiseaux font partie).
L'innovation est variation. Elle est rendue possible par l'absence de
rigidité des mécanismes dynamiques d'interactions entre
populations. Elle est limitée mais non figée car elle
se répète toujours, embryon après embryon. Cette
variation serait toujours ouverte, mais pas au même niveau. En
effet, si son répertoire s'amenuise de plus en plus au cours
des temps géologiques, cela explique pourquoi, actuellement,
cette variation est limitée à la spéciation. La
mise en mémoire de cette nouvelle phanère s'est
résumée à l'enregistrement non pas d'un
mécanisme à reproduire (programme plume) mais d'une
structure (tube) issu d'un arrangement spatial nouveau d'une
population cellulaire qui peut croître dans plusieurs
directions de l'espace avec plus ou moins d'intensité (qui
possède donc plusieurs alternatives de développement:
structure plane ou tubulaire par exemple; je renvoie
aux structures
proposées par Yves Bouligand
à partir de calculs réalisées sur les formes
membranaires: je pense que ces calculs sont tout à fait
utilisables ici en travaillant sur les différentes couches
épidermiques et dermiques en présence). Les
écailles, les plumes et les poils étant des "solutions"
différentes d'un même "problème" de dynamique des
structures.
Cette interprétation de la variation cadre avec plusieurs études réalisées dans le cadre d'un nouveau paradigme (qualifié de réalisme écologique: voir page d'accueil et page sur les modèles). Pour ceux qui souhaiteraient s'intéresser à cette question, je propose une analogie avec la croissance du Pin. Une étude réalisée par Stéphane Jourdan est disponible à l'adresse: http://www.ifrance.com/tahitinui//pinus/pinli13.htm. En quelques mots: il propose d'interpréter des axes peu ramifiés à croissance rapide (dits "queues-de-renard" ou foxtails) à l'aide d'un découplage croissance/développement dont la cause serait "chaotique" dans le sens où elle résulterait d'une alternative de développement déclenchée par des facteurs externes.