La cellule
Retour Histoire de science - page
d'accueil, cours lycée
Cette page a été
modifiée en octobre 2007 dans une nouvelle page:
Au-delà
de la cellule
La théorie
cellulaire (Buffon, Oken, Vischov) comprend deux
affirmations :
* tout être vivant est composé en
totalité (et uniquement) d'au moins une cellule
* et toute cellule est issue d'une autre cellule.
Affirmations auxquelles G. Canguilhem
ajoute deux autres axiomes (dont il attribue la paternité aux
personnalités dont les noms sont cités entre
parenthèses) :
* les vivants non composés sont
unicellulaires (Dujardin, Haeckel)
* l'œuf d'où naissent les
organismes vivants sexués est une cellule dont le
développement s'explique uniquement par la division
(Schwann, Kölliker).
La cellule vivante
(en travaux 2005)
Sources:
L'eau dans la cellule vivante; État physique,
fonctions, Pascale Mentré, 2002, in L'eau dans les
aliments, M. Le Mestre, D. Lorient et D. Simatos coord., Tec
& Doc (ISBN: 2-7430-0523-8)
L'eau dans la cellule, Pascale Mentré, 1995, Masson
Encyclopedia Universalis, article "vie",
G. Canguilhem.
La théorie cellulaire, G.
Canguilhem in La connaissance de la vie, Hachette, 1952 (pp
47-98)
Comment les cellules construisent l'animal, Rosine CHANDEBOIS,
1999, Phénix éditions, Paris
Le gène et la forme (ou la démythification de
l'ADN), Rosine CHANDEBOIS, 1989, Ed. Espaces 34
Le cycle cellulaire chez les animaux et les
végétaux, Jean Clos, Marc Coumans et Yves Muller,
Biologie-Géologie, 3-2002, p 497-564
Un site en anglais (de Martin Chaplin) particulièrement riche:
http://www.sbu.ac.uk/water/index.html
Un site à recommander avec des vidéos et un texte clair
:http://www.cerimes.fr/e_doc/cellule/cellule.htm
Avertissement
Contrairement aux apparences, relayées par le titre et les
intertitres, cette page n'est qu'un essai de synthèse
d'éléments modernes et anciens qui, bien souvent,
dépassent le cadre de la théorie
cellulaire mais, pour cette fois encore, sans chercher à
le dépasser.
La cellule vivante n'est pas plus une chambre (cellula en
latin), qu'une solution aqueuse, ni encore qu'une unité d'un
programme du vivant. Une cellule est tout simplement un moyen commode
que scientifiques et philosophes ont trouvé pour
décrire le vivant. C'est une unité structurale et
fonctionnelle. La cellule comme unité de structure
découle du premier axiome de la théorie (tout
être vivant est composé d'au moins une cellule). La
cellule comme unité fonctionnelle est le plus petit
élément VIVANT, qui possède TOUTES les
caractéristiques du vivant. Si on reprend la formulation qui
m'est chère, c'est la plus petite unité AU TRAVAIL:
plus petite unité qui réalise un travail de nutrition,
de relation et de reproduction. Le deuxième axiome de la
théorie cellulaire (toute cellule est issue d'une autre
cellule) permet de repousser la question de la vie hors du champ
biologique et vers le champ philosophique, mais n'est pas une
réponse. La biologie doit se contenter de cette
propriété du vivant qu'elle ne peut relier à
aucun objet sans sortir de sa méthode (qui est
expérimentale et qui ne peut répondre à la
question de la vie). La question de la pluricellularité est
solutionnée en disant que chaque cellule est vivante mais
n'est pas un être vivant, autonome. Seule la totalité de
l'être vivant, composé d'innombrables cellules, peut
être qualifié d'être vivant. C'est aussi une fuite
ou une esquive et non une réponse. Un début de
réponse (philosophique) peut être apporté si l'on
considère l'organisme pluricellulaire comme une
société de cellules.
|
Plan de cette page:
|
|
1 Deux barrières à ouvrir pour
une nouvelle conception de la cellule:
1ère
barrière entrouverte: l'unité
structurale de la cellule ne vient pas de ses
membranes (fluides) ou de structures
macromoléculaires plus ou moins complexes baignant
dans une solution aqueuse mais de son eau qui est,
pour sa quasi-totalité, "coincée" au sein
de la foule de macromolécules qui remplit la
cellule: cette eau est interfaciale (structurée)
et c'est elle qui contrôle le fonctionnement
cellulaire:
Une cellule surpeuplée de
macromolécules (le
terme anglais de crowded est
particulièrement
évocateur).
Une cellule riche en eau mais en eau structurée
et fonctionnelle.
2ème
barrière entrouverte: l'unité
fonctionnelle cellulaire ne vient pas d'une chimie du
hasard (les lois probabilistes des milieux
homogènes) mais d'une biochimie qui pourrait
être fondée sur la
finalité:
la vie cellulaire est un travail
orienté vers une fin biologique.
2. Des chiffres anciens, un regard nouveau
(en cours)
|
Ce schéma personnel ne doit pas
être utilisé sans mon accord; je suis
intéressé
par toute remarque à son sujet
Ce schéma a pour but de fixer les ordres de
grandeurs des dimensions et des objets observables ou imaginés
aux différentes échelles (il a
été réalisé pour une cellule eucaryote
mais peut aussi s'appliquer, en tenant compte de l'absence de noyau
et de structures membranaires internes, à la cellule
procaryote):
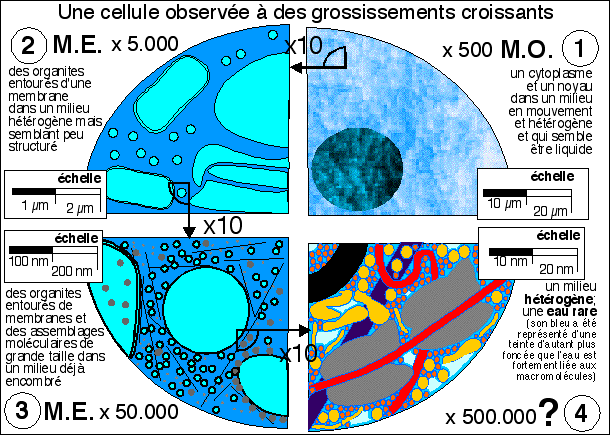
- le microscope optique (M.O. -
1) ne permet guère que de voir le noyau et le
cytoplasme dont le contenu nous paraît clair mais
hétérogène (un peu grumeleux). On peut aussi
entr'apercevoir de gros organites comme des vacuoles, des
chloroplastes ou des mitochondries. On peut colorer la cellule
sans la tuer et voir par exemple les chloroplastes se
déplacer entraînés par les mouvements du
cytoplasme.
- Au microscope électronique
(M.E. 2 et 3) la cellule est morte
et fixée. Les structures ne sont pas vivantes mais mortes.
De plus, on ne voit que celles qui fixent le colorant
utilisé (tétroxyde d'osmium). On observe cependant
une grande variété de structures. En
2 on distingue des mitochondries,
des vésicules et les citernes du réticulum. En
3 les plus gros assemblages
protéiques apparaissent comme les ribosomes et les
microfilaments du cytosquelette.
- L'étape
4 n'est pas vraiment
accessible à la microscopie pour des structures complexes
en place mais représente ce que l'on peut imaginer
être la réalité à partir d'observations
de quelques éléments réellement
observés mais isolés de la cellule et d'autres
structures. On reconnaîtra en partant du coin gauche et en
se déplaçant en diagonale vers le bas à
droite: une vésicule dont la membrane (en noir) est une
structure de près de 8 nm d'épaisseur, des
macromolécules (en jaune) avec leur(s) couche(s) d'eau
liée (les plus grosses protéines globulaires
atteignent typiquement 5 nm de diamètre), un microfilament
fin dessiné en arrière plan (en violet, leur
diamètre varie de 5 nm pour les plus fins à 25 nm
pour les plus épais), deux ribosomes (en gris, de
diamètre environ 20 nm) autour d'un ARNm (en rouge, un
filament de diamètre inférieur à 2 nm qui est
le diamètre de la molécule d'ADN); les
molécules organiques plus petites (de diamètre
voisin du nanomètre - pensez qu'un aa comme la
phénylalanine atteint déjà le
nanomètre si l'on tient compte de sa couche d'eau
liée - voir plus bas) avec leur(s) couche(s) d'eau
liée encombrent le hyaloplasme; l'eau "libre" (en bleu
clair) est rare et concentrée dans certains espaces.
1. Deux barrières à ouvrir pour une
nouvelle conception de la cellule
1ère barrière
entrouverte: l'unité structurale de la cellule ne vient
pas de ses membranes (fluides) ou de structures
macromoléculaires plus ou moins complexes baignant dans une
solution aqueuse mais de son eau qui est, pour sa
quasi-totalité, "coincée" au sein de la foule de
macromolécules qui remplit la cellule: cette eau est
interfaciale (structurée) et c'est elle qui contrôle le
fonctionnement cellulaire:
Une cellule surpeuplée de
macromolécules (le terme anglais de
crowded est particulièrement
évocateur).
Une cellule riche en eau mais en eau structurée et
fonctionnelle.
Deux visions de la cellule
s'affrontent encore...
|
une vision classique...
physique
|
vers une vision plus complexe...
biologique
|
|
l'eau, solution du
vivant moléculaire
une cellule est un sac rempli d'eau et de
molécules organiques et minérales en
solution aqueuse, siège d'innombrables
réactions chimiques qui façonnent sa forme et
son activité
|
l'eau structurante,
structurée et exerçant un contrôle
fonctionnel sur les complexes moléculaires du
vivant
une cellule est entièrement
structurée, dynamique (en évolution:
naissance, vie et mort), autonome (au fonctionnement
intégré mais dont l'unité
dépasse la somme des parties), vivante (au travail:
de relation, de nutrition et de reproduction); les
molécules ne sont plus en solution mais
complexées, transportées, associées
à des structures d'ordre supérieur
|
|
les structures intracellulaires sont des assemblages
moléculaires stables : des complexes fonctionnels
comme les ribosomes ou structurants comme les membranes
(auxquelles sont associés des complexes fonctionnels
enzymatiques par exemple). Ces structures sont
dispersées ou regroupées dans le
hyaloplasme, qui est fluide (ou à l'état de
gel).
|
La totalité de la cellule peut être
décrite à l'aide d'assemblages
moléculaires et fonctionnels comme une membrane
ou un ribosome: il n'y a pas d'espace, sauf vésicules
particulières, où règne un milieu
aqueux où puisse s'appliquer la chimie des solutions;
lorsqu'on décrit un système moléculaire
en interaction avec l'ADN, il faut prendre en compte toutes
les relations de voisinage structural et non pas seulement
les molécules que l'on pense pouvoir s'accoler
directement à l'hélice d'ADN: l'action des
molécules peut être mécanique par
resserrement de la double hélice et agir donc
à distance, voir, depuis la membrane cytoplasmique...
changer notre conception du hyaloplasme change
singulièrement notre approche des
régulations... qui ne sont plus toujours
chimiques mais qui peuvent être
mécaniques, thermiques où
ioniques...
|
|
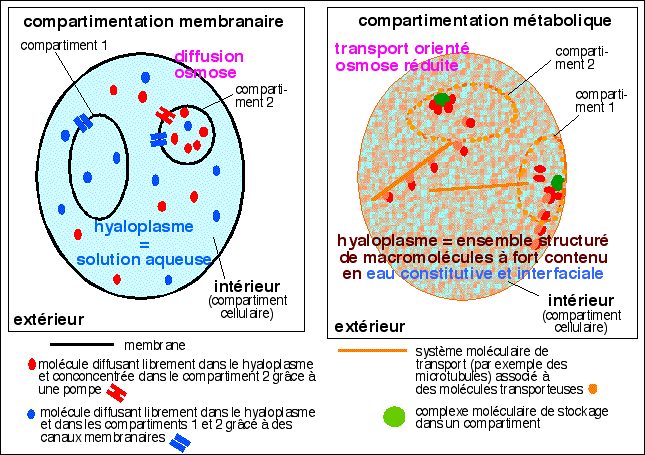
Les membranes délimitent des compartiments
où sont réalisées certaines parties
du métabolisme; les molécules hydrosolubles
diffusent librement dans chaque compartiment et passent d'un
compartiment à l'autre à l'aide de pompes ou
de canaux
|
Les compartiments ne sont plus seulement membranaires
mais doivent être redéfinis en terme
d'accessibilité des molécules aux
différentes réactions chimiques
réalisées dans ce compartiment et en termes
d'échanges entre compartiments: la compartimentation
est aussi bien temporelle que spatiale: le terme de division
du travail est on ne peut plus d'actualité; les
molécules sont toujours transportées dans un
compartiment aussi bien que d'un compartiment à
l'autre. Ces transports peuvent rendre compte de
métabolismes rapides.
|
De l'observation à
l'imagination
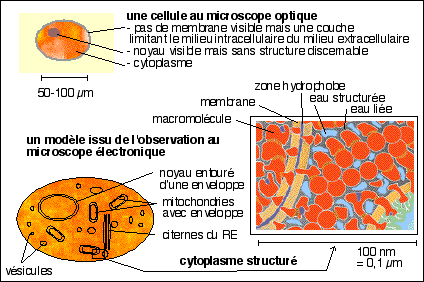
Un essai de représentation d'un modèle
biologique d'une cellule "théorique" comme
"complexe moléculaire structuré"
(le schéma du cytoplasme est
extrait et très modifié de Pascal
Mentré, L'eau dans la cellule, Masson, 1995, p 173
fig 8.7)
|
Quelques points majeurs, essentiellement d'après
Mentré, 2002 (à qui on se reportera
pour les références aux publications scientifiques dont
les auteurs ne seront pas cités dans cette page de
synthèse, étant donné que je ne les ai pas
lus... ):
- l'eau représente près de 80% en masse
de la cellule vivante mais cette eau est à près de
80% sous forme d'un film interfacial d'environ 1,1 nm
d'épaisseur qui se trouve entre les molécules
organiques structurant la cellule. Il reste donc 20% d'eau libre
où s'appliquent les lois de la chimie des solutions
aqueuses, principalement de l'eau présente dans des
vésicules.
Pour comprendre comment une augmentation de la surface de contact,
qui est due à la faible taille des macromolécules,
augmente considérablement le volume d'eau associé
à un ensemble de macromolécules, on peut prendre
l'exemple de l'ADN: à chaque nucléotide est
associé environ 20 molécules d'eau (eau "liée",
très structurée, non congelable) ce qui
représente environ 80% du poids de la molécule d'ADN.
Si l'on place de l'ADN cristallisé en solution aqueuse il
gonfle, ce que l'on peut expliquer par la mise en place d'au moins
une seconde couche d'eau entre les brins d'ADN, couche moins
structurée et congelable, dont l'origine pourrait provenir du
caractère fortement électrolytique de la
molécule d'ADN. Dans une cellule vivante, l'eau
associée à l'ADN atteint ainsi une masse nettement
supérieure à la masse d'ADN. On peut donc comprendre
que, tout comme ses composants moléculaires, dont la forme est
à la fois stabilisée par l'eau et due en partie
à cette hydratation, la cellule contient une énorme
quantité d'eau, fortement structurante et
structurée.
L'ADN, une molécule très
hydratée
|
|
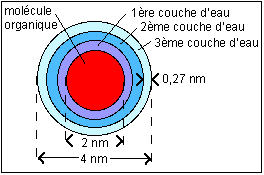
Pour une molécule organique de 2 nm de
diamètre (de l'ordre de celui de la
molécule d'ADN) supposée
sphérique (donc de volume 33,5
nm3), une couche d'hydratation dont
l'épaisseur est estimée à 0,27
nm correspond à un volume de (4/3 º
(2,273-23)nm3=
15,5 nm3 ) soit un volume relatif de
(2,273-23/23 )
soit 46%. Avec une deuxième couche
d'eau on atteint 105 % et 177% avec
une troisième couche d'eau.
Il faut cependant raisonner en masse et non
en volume et avec des molécules de forme
très complexe, mais toute augmentation
de la surface de la molécule va bien de le
sens d'une augmentation de la masse d'eau
associée dans des proportions très
importantes qui ne sont évidentes que pour
un chimiste.
|
|
|
Il est évident que, si la majorité des
scientifiques actuels s'accordent pour la présence
d'une couche d'eau "liée", l'épaisseur de la
"coquille d'hydratation" est fortement
discutée par les spécialistes.
La première couche d'eau est appelée
eau liée
(bound water).
Les couches d'eau d'ordre supérieur sont
appelées eau
structurée (structured) ou
polarisée (polarized) ou contrainte
(strained) ou encore vicinale (vicinal).
L'eau dont les molécules ne sont pas en interaction
avec la macromolécules forme une eau "libre"
qualifiée d'eau
bulk. C'est
par exemple l'eau contenue dans des vésicules (de
pinocytose) d'un diamètre de l'ordre de 100 nm ou
supérieur (vacuole des plantes ?).
Enfin il existe aussi une eau constitutive des
macromolécules, inextractible sans modifier
profondément la structure et la fonction de la
molécule.
|
|
En sachant que l'empilement de sphères le
plus dense atteint près de 75% du volume
total (c'est la fameuse conjecture
de Kepler qui a reçu une
démonstration faisant intensément
appel à l'informatique par Hales en
1999), on peut faire entrer
approximativement, dans un volume Vo donné
d' environ 70 µm3, 100
sphères de 1
micromètre (1 µm) de diamètre,
possédant une couche
d'eau liée environ 1,1 nm
d'épaisseur ou 73 millions de sphères
d'un diamètre 100 fois plus petit (10 nm)
avec une couche d'eau liée de la même
épaisseur.
Le volume d'eau liée dans le premier cas est
de l'ordre de 0,2 µm3 (en fait
0,25% du volume total) alors que dans le second
cas, il atteint 14 µm3 (soit 20% du volume
total).
Pour des ions hydratés les
calculs sur le volume sont inefficaces mais on peut
raisonner sur le nombre d'ions et de
molécules. L'eau représente bien plus
de 50% du volume. Un ion K+ est
solvaté en moyenne par 4 molécules
d'eau. Le rayon ionique (voir plus bas) est de
l'ordre de 0,121 nm ce qui ferait plus de 7.000
milliards d'ions dans un volume de 70
µm3. Ce qui correspondrait à
un nombre 4 fois plus élevé de
molécules d'eau associées. En masse
on a 35% de K+ pour 65%
d'eau.
Cette analogie géométrique peut
aider à comprendre comment une cellule
contiendrait 80% de son eau sous forme d'eau
interfaciale (liée) constituant un microfilm
de 1,1 nm d'épaisseur autour des
macromolécules cellulaires.
|
|
|
DIVISER pour CROÎTRE
en surface
|
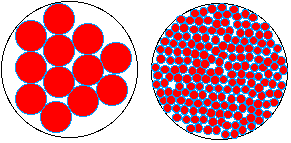
|
volume Vo (environ 70
µm3)
|
|
100
sphères
|
73 millions
de sphères
|
|
1 µm de
diamètre
|
10 nm de
diamètre (1/100
µm)
|
|
une couche
d'eau liée de 1,1 nm
d'épaisseur
|
|
volume d'eau
liée d'environ 0,2
µm3 soit
0,25% du volume
total
|
volume d'eau
liée d'environ 14
µm3 soit
20% du volume
total
|
|
|
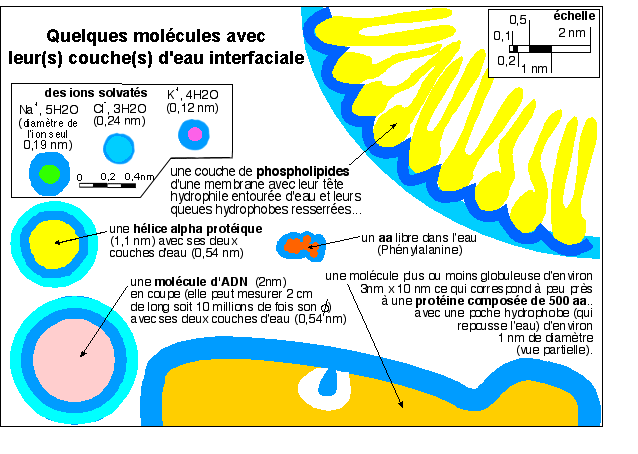
Même si les spécialistes discutent pour savoir quel est
le nombre habituel de couches d'eau liée autour des
macromolécules, elles sont omniprésentes. Une
molécule est toujours entourée d'eau plus ou moins
fortement attachée à elle. Cette eau est
ordonnée selon les interactions qu'elle développe avec
la molécule à laquelle elle se lie: on parle ainsi
d'eau liée ou d'eau structurée. Par rapport à la
figure 1, cette figure correspondrait à un grossissement
environ dix fois plus fort le grossissement le plus fort de la figure
1 soit environ 5.000.000 de fois; ce qui n'est bien sûr pas
accessible à une quelconque technique microscopique sur des
molécules en place dans un cytoplasme, et encore moins sur de
la matière vivante. Nous devons donc nous contenter
d'interprétations, réalisées par des
méthodes indirectes, et non d'observations.
|
Quelques formes de l'eau structurée
interfaciale:
- les molécules d'eau établissent des liaisons
hydrogène entre elles (structure ice-like) qui
sont fortement favorisées dans les poches hydrophobes
et au voisinage des domaines apolaires des
molécules;
- les molécules d'eau (polarisées) sont
fortement attirées par les domaines
ionisés des molécules et forment une
eau électrostrictée dont la
densité peut atteindre 1,2. Les molécules
d'eau électrostrictée s'échangent sans
cesse avec l'eau bulk;
- les molécules d'eau sont fortement
immobilisées par des liaisons hydrogène (de
structure variées) au niveau des domaines
polaires.

Les liaisons hydrogène sont responsables de nombre de
propriétés originales de l'eau liquide et
solide (voir page sur
l'eau).
La petite taille du proton est à l'origine de
l'intensité de la liaison hydrogène car un
champ électrostatique engendré par une
particule de charge q varie en q/d2, si d est la
distance au centre de la particule. Le proton forme un pont
entre un atome donneur auquel il est lié de
façon covalente (O) et un atome accepteur qu'il
attire électriquement (O ou N, mais aussi C ou encore
un nuage d'électrons º d'un cycle aromatique...) .
L'habituelle linéarité de la liaison
hydrogène observée entre molécules
d'eau dans la glace, n'est pas conservée dans toutes
les liaisons hydrogène qui peuvent aussi
présenter des énergies variables en fonction
de leur longueur (8 kJ.mol-1 pour les plus
courtes liaisons à 42 kJ.mol-1 pour les
liaisons les plus longues). L'établissement d'une
liaison hydrogène modifiant les
caractéristiques du donneur et de l'accepteur, on
observe un phénomène de
coopérativité qui facilite
l'établissement de liaisons en chaîne. Ces
chaînes de molécules d'eau liée
expliqueraient nombre de fonctions de l'eau
interfaciale.
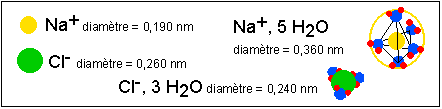
La solvatation résulte de
l'établissement de liaisons électrostatiques
entre les molécules d'eau et les ions et de liaisons
hydrogène entre les molécules d'eau de cette
"couche" dite de solvatation. Le nombre moyen de
molécules d'eau de la couche de solvatation
dépend de la charge de l'ion et de son rayon
atomique: plus l'ion est chargé plus il attire les
molécules d'eau; et à charge égale,
plus un ion a un rayon ionique important, moins il attire de
molécules d'eau. Pour un ion de grande taille de
charge négative comme le chlore, cette charge est
répartie sur toute la surface de l'ion (1/S=1/º
R2) est est donc plus faible que pour un ion de
petite taille comme de sodium. Ainsi l'ion chlorure est
solvaté par environ 3 molécules d'eau pour un
rayon de solvatation inférieur au rayon de l'ion dans
un solide ionique (NaCl) alors que l'ion sodium est
solvaté par environ 5 molécules d'eau et son
rayon ionique est presque deux fois plus faible que
lorsqu'il est solvaté. Un autre exemple est le
lithium (ion Li+) qui est solvaté par environ 6
molécules d'eau ce qui fait passer son rayon ionique
de 0,060 nm à 0,236 nm (soit près de 4 fois
plus) lorsqu'il est solvaté. L'ion potassium (K+),
volumineux, et ne portant qu'une seule charge positive,
n'est solvaté que par 4 molécules d'eau en
moyenne, et passe d'un rayon ionique de 0,133 nm à
0,121 nm lorsqu'il est hydraté.
Dans un champ électrique, EN SOLUTION, les ions les
plus petits et les plus solvatés migrent le plus
lentement.

Les clathrates sont des structures
polyédriques à faces pentagonales qui
contiennent des molécules d'eau réunies par
des liaisons hydrogène et des molécules
hydrophobes (on connaissait les clathrates
en géologie où c'est une forme d'association
de l'eau avec les hydrocarbures, et en astrophysique
où ces structures ont été
utilisées pour expliquer certaines
caractéristiques spectroscopiques des rayonnements de
certaines corps célestes) . Les clathrates
sont très stables, cristallisables, et peuvent
contenir jusqu'à 90% d'eau.
On pense que des structures de type clathrates, imparfaites,
seraient extrêmement fréquentes pour les
molécules biologiques hydrophobes et même au
voisinage des domaines hydrophobes de
macromolécules.
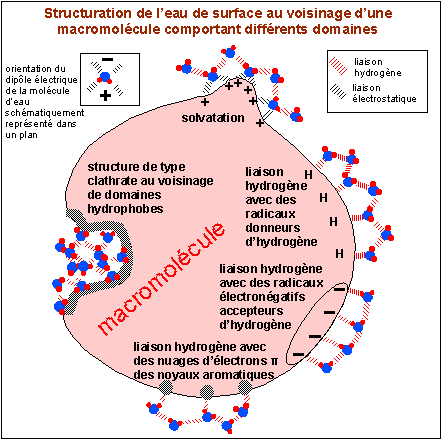
Les 3 types principaux de structuration de l'eau au
voisinage d'une macromolécule: eau de solvatation
(remarquez la seconde couche d'eau liée à
la première par des liaisons
hydrogène: la première couche
est appelée eau liée (sous-entendu à la
macromolécule), la seconde, eau
structurée), eau liée par des
liaisons hydrogène à des domaines polaires
ou à des nuages d'électrons º (là aussi
une seconde couche d'eau stabilise la première
couche), eau liée par des liaisons
hydrogène en structures de type clathrate au
voisinage de domaines hydrophobes
(d'après Mentré, 1995, Fig
12.2 principalement).
|
- L'eau cellulaire est très
hétérogène:
* elle est polarisée (anisotrope),
* elle présente des variations importantes dans son
architecture, son degré de coopérativité et sa
densité,
* elle possède un pouvoir solvant réduit,
* elle possède une activité osmotique
réduite.
- L'eau cellulaire n'est pas immobile mais
percole la cellule.
Toutes les liaisons évoquées ci-dessus sont des
liaisons dynamiques dont le turn-over est rapide. Une molécule
d'eau engagée dans une liaison électrostatique avec un
domaine ionisé d'une molécule reste moins longtemps
dans cet état que dans l'eau bulk qu'elle rejoint au bout de
quelque temps. L'eau interfaciale peut aussi se déplacer avec
les structures cellulaires qu'elle enrobe, et ce, avec des vitesses
bien supérieures à celle de la diffusion dans une eau
bulk. On peut considérer que les molécules de toute
petite taille sont capables de percoler avec l'eau interfaciale. La
plupart des macromolécules, qui sont d'ailleurs souvent
associées en complexes moléculaires, sont
transportées.
Il n'en reste pas moins que ce que l'on nomme l'autodiffusion de
l'eau dans la cellule reste un phénomène à
explorer ( ce qui a déjà été
commencé mais sort du cadre de cette synthèse partielle
- voir Mentré,2002, p 103-104 et
références) à la lumière de cette
nouvelle vision de la cellule vivante.
Le transport de l'eau par les vésicules de pinocytose reste
aussi un mode de transport rapide et efficace.
- les transports de molécules,
d'ions, de signaux se font d'autant plus rapidement qu'ils sont
associés à des structures de transport de grande
taille;
par exemple au sein d'un axone on estime la vitesse de diffusion de
protéines isolées (de diamètre de l'ordre de
quelques nm) inférieure à 0,01 µm.s-1,
alors que avec des protéines transporteuses (de
diamètre de l'ordre d'une dizaine de nm) on atteint des
vitesses de l'ordre de 0,05 µm.s-1 et enfin, avec des
vésicules (de l'ordre de 100 µm de diamètre) on
atteint des vitesses de l'ordre de 0,5 µm.s-1. Pour
chaque ordre de grandeur de taille on gagne un ordre de grandeur en
vitesse (in Mentré, 2002, p 91).
- les réactions chimiques du
métabolisme se font dans des structures complexes, sans
eau libre, ce qui explique des rendements proches de
100%;
la faiblesse des rendements en solution aqueuse peut être due
au peu d'accessibilité des substrats; l'excellent rendement
pour des enzymes associées en complexe est dû à
la diminution du temps de transit d'une enzyme à l'autre; on
désigne ce mécanisme d'association par le terme de
canalisation (chanelling) ou d'une façon plus
générale de compartimentation métabolique
qui exprime l'idée d'association des enzymes impliquées
dans une même chaîne métabolique .
La compartimentation métabolique est donc une notion qu'il
faut étendre à la totalité de la cellule,
en relativisant la notion de membrane et des canaux ou transporteurs
qui lui étaient associés. Un compartiment
métabolique peut très bien se répartir à
la fois dans un organite, le long d'une membrane et à
l'extérieur de celle-ci.
L'eau structurée pourrait servir de réseau de
coordination des signaux intracellulaires au sein de laquelle les
transports seraient nettement plus rapide que dans une eau libre; des
résultats assez convaincants ont déjà
été obtenus avec des mouvements de protons qui
transmettent des signaux extérieurs en signaux
intracellulaires.
- les concentrations in vivo sont trop
faibles pour obéir aux lois de la chimie des solutions:
la notion de gradient de concentration perd tout sens dans une
cellule:
on estime qu'une bactérie comme Echerichia coli
ne contient qu'une centaine de protons qui sont
éloignés les uns des autres de plusieurs centaines de
nm et donc ne peuvent être responsables d'un gradient de
concentration. De même pour les ions calcium que l'on estime
à une concentration cellulaire de 10-3M dont
10-8M à 10-6 libres (non complexé
par des macromolécules comme la calmoduline...) mais
solvaté (entouré par des molécules d'eau).
|
Dans une bactérie comme Escherichia coli
que l'on peut estimer parallélipipédique
(de volume V=2.1.1 µm3 = 2.10-15
L), une concentration de 10-6M correspond
à un nombre de molécules de calcium n = N
(nombre d'Avogadro). 10-6. V soit n =
6.1023.10-6.2.10-15
molécules = 1200 molécules. De la même
façon, une concentration de 10-8M
correspond à un nombre de molécules de calcium
n = N (nombre d'Avogadro)x 10-8. V
molécules soit n =
6.1023.10-6.2.10-15
molécules = 12 molécules. En supposant chaque
molécule au centre d'un cube de côté
"a", la distance entre deux molécules est très
approximativement égale à "a" soit RACINE
CUBIQUE [(V/n)]. On trouve
0,1 µm pour la concentration de
10-6M et
0,6 µm pour la
concentration de
10-8M. Pour
une cellule supposée sphérique, on a les
mêmes ordres de grandeur.
|
Comment imaginer qu'une dizaine d'ions ou même quelques
milliers d'ions, séparés par de très nombreuses
macromolécules puissent déterminer un gradient de
concentration à de telles distances ! Quand aux compartiments
membranaires habituellement invoqués pour présenter une
concentration homogène, leur taille étant de l'ordre de
la centaine de nanomètre (épaisseur d'une citerne
golgienne ou du réticulum endoplasmique) au micromètre
(mitochondrie), ils ne peuvent non plus contenir plus de quelques
unités à dizaines d'unités de ces ions, bien
incapables donc de déterminer un gradient de concentration.
Cette remarque d'une part remet en cause probablement la mesure
des concentrations aussi faibles que celles
présentées et d'autre part oblige à
considérer que les molécules sont avant tout
liées à d'autres molécules dans la cellule et
que l'eau ne joue que très rarement, le rôle de
solvant.

Les concentrations extrêmement faibles estimées pour le
vivant conduisent à des interprétations trompeuses si
l'on pense que le compartiment cellulaire bactérien, par
exemple, est un milieu homogène où la chimie des
solutions en milieu aqueux pourrait s'appliquer. Ainsi une
concentration en calcium d'une nanomole correspond à environ
12 ions calcium dans le cytoplasme. Avec des chiffres aussi petits il
est indispensable d'envisager une chimie des milieux
hétérogènes pour le cytoplasme dont une
représentation très imagée est proposée.
On reconnaîtra la membrane plasmique (en orange) avec des
protéines membranaires (en jaune) , des ribosomes (en gris) et
des éléments du cytosquelette (en violet). Le calcium
libre est très probablement séquestré dans des
microdomaines très localisés (en bleu foncé)
dans lesquels on peut peut-être utiliser une chimie statistique
avec par exemple la loi d'action de masses, mais il est
incohérent de l'appliquer à l'ensemble du compartiment
cellulaire.
- les propriétés physiques du
cytoplasme, sont hétérogènes et
très variables selon les types cellulaires, ce qui peut
notamment être expliqué par les différentes
formes de l'eau au sein de ce cytoplasme:
le hyaloplasme ne formerait pas une entité homogène au
comportement physique identique sauf dans quelques cellules
très spécialisées comme les hématies dont
la compartimentation est réduite à un quasi-unique
compartiment, monofonctionnel. L'immense vacuole des cellules
végétales peut aussi constituer un bon exemple de
compartiment d'une telle homogénéité. La
pression osmotique par exemple est un facteur que l'on ne peut
probablement étendre à l'ensemble du compartiment
cellulaire que dans ces cas exceptionnels. Il est bien plus probable
que la quasi-totalité des cellules présentent
plutôt une organisation en microdomaines où la
pression osmotique est un facteur qui dépend des conditions
locales principalement.
Une des théories les plus séduisantes sur
l'organisation de l'eau cellulaire est la théorie de
Watterson (1982,1991); la tension existant dans un volume d'eau
est reliée par l'auteur aux mouvements vibratoires des
molécules qui seraient dus aux phénomènes de
coopérativité (résonance) observés entre
les molécules d'eau reliées par des liaisons
hydrogène. Il a ainsi mesuré que le cytoplasme serait
organisé en domaines (clusters) d'environ 40
nm3 dont le diamètre correspond à la
longueur d'onde vibrationnelle, voisine de 3,4 nm, les nœuds
étant situés aux points où la tension est la
plus faible. Chaque cluster contiendrait environ 1300 à 1400
molécules d'eau (pour un poids moléculaire de 24kDa).
Un point essentiel d'application de cette théorie est que la
plupart des protéines intracellulaires ont des
dimensions qui correspondent précisément à un
cluster: 40 nm3 pour un nombre de résidus compris
entre 120 et 300 et un diamètre moyen de 3,5 nm. Ces clusters
ne seraient pas des structures rigides mais dynamiques, très
sensibles aux éléments introduits; les protéines
polymériques par exemple, d'une taille supérieure
à un cluster, modifiant la résonance de l'ensemble.
Watterson propose de relier la stabilité de la matrice
hyaloplasmique à l'harmonie vibratoire des domaines d'eau et
de macromolécules entourées d'eau. La cohérence
résultant alors non pas de forces de liaison mais de
mouvements vibratoires résonnants, sensibles aux perturbations
(et permettant ainsi de transférer extrêmement
rapidement un signal (modification de la longueur d'onde...) d'un
point à un autre) mais résistants grâce à
leur souplesse.
- l'eau n'est donc plus seulement
constitutive du vivant (matière vivante, milieu de vie)
mais ses rôles se découvrent aussi
énergétiques (mécaniques, chimiques...)
et relationnels.
On n'a encore que quelques rares éclairages sur ces nouveaux
rôles de l'eau qui n'est plus essentiellement un solvant mais
une interface (détails in Mentré, 2002, p
94-101). C'est un renversement complet de la compréhension de
l'eau qui tient désormais un rôle actif:
- la première couche d'eau interfaciale (eau liée)
semble joue le rôle d'élément de
contrôle principal, du fait notamment de sa forte
coopérativité (elle est plus difficilement
perturbable); la seconde couche étant surtout un
réservoir. Mais les échanges entre ces deux
couches (mécanisme appelé micro-osmose) sont
eux-mêmes un élément essentiel du pouvoir
régulateur de l'eau structurée en
général.
- l'eau interfaciale, par ses liaisons multiples avec des
domaines variés des complexes moléculaires
contrôle leur assemblage-désassemblage et
leur fonctionnement.
L'eau impliquée dans les assemblages des complexes
macromoléculaires, par exemple avec les liaisons
hydrogène établies avec les domaines polaires,
interviendrait aussi pour orienter ces assemblages et
participerait à la dynamique de leur
assemblage-désassemblage. De la même manière,
l'eau interfaciale semble intervenir directement dans le
contrôle de l'activité de molécules
fonctionnelles comme par exemple les enzymes. Récemment on
a attribué la transition allostérique (avec
changement de conformation et coopération entre les 4
sous-unités protéiques) entre la molécule
d'hémoglobine désoxygénées (Hb) et la
molécule oxygénée (HbO8) à la liaison
de 60 molécules d'eau à la molécule d'Hb,
cette eau provenant essentiellement de domaines ice-like de
l'eau interfaciale associés à des domaines
hydrophobes de la molécule d'Hb. L'eau est directement
impliqué dans de nombreux autres processus que l'on
découvre petit à petit: réactions
allostériques, dénaturation des protéines,
assemblage des microtubules, dénaturation de l'ADN... ,
notamment à travers le processus de compensation
enthalpie-entropie qui manifeste thermodynamiquement la
structuration-déstructuration de l'eau
(voir références in
Mentré 2002, p 97).
On peut aussi citer les nombreuses enzymes activées
spécifiquement par le potassium et non par le sodium. Le
contrôle de l'activité de ces enzymes serait
directement due à l'eau interfaciale. En effet, le site
catalytique des enzymes est généralement hydrophile
mais situé la plupart du temps au fond d'une poche
hydrophobe. Quand le substrat se lie au site de l'enzyme le
caractère hydrophobe de la poche s'accentue ce qui tend
à structurer davantage l'eau (ice-like). Cette
structure à la propriété d'exclure fortement
le Na+ mais non le K+ (voir deux paragraphes plus loin,
l'utilisation de la série de Hofmeister). Les ions K+
entrent alors dans la poche et déstructurent l'eau
(état vapor-like) qui augmente de volume. Cette
effet mécanique entraînerait l'ouverture de la poche
et la libération du produit de la réaction
enzymatique (in Mentré, 2002, p 96,
§3.2.2).
- l'eau interfaciale, comme son nom l'indique,
contrôle, par sa position, les
mécanismes informationnels supposant une
reconnaissance moléculaire
(topologique).
Un exemple démonstratif est ce l'on nomme l'effet
star: le pourcentage de coupures non canoniques
(erronées) réalisées par une enzyme de
restriction (EcoR1, dont la séquence canonique est AATT)
augmente linéairement avec la pression osmotique. Les
auteurs de cette expérience l'interprètent comme la
nécessité de l'expulsion de l'eau interfaciale du
domaine de liaison enzyme-ADN. Aux fortes pressions hydrostatiques
l'eau interfaciale est stabilisée, la reconnaissance ne
peut se faire et toute activité de coupure enzymatique est
bloquée. On peut extraire une partie de l'eau d'hydratation
en augmentant la pression osmotique, dans ce cas on favorise la
reconnaissance et la liaison enzyme-ADN et donc la coupure de
celui-ci, même pour des sites non canoniques.
- les chaînes de molécules d'eau, à
la surface de macromolécules où reliant des
molécules entre elles, présentent, comme dans la
glace (mais avec une mobilité réduite cependant),
une conductivité protonique efficace qui pourrait
emprunter les circuits de conduction de l'eau interfaciale
formant, selon l'expression de R. Peters un réseau
subtil coordinateur.
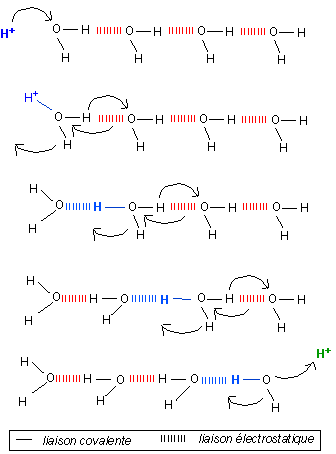
Conduction protonique
ou
"transfert" d'un proton le long d'une chaîne de
molécules d'eau liées par des liaisons
hydrogènes (d'après P.
Mentré, 1995 et 2000)
Si le phénomène n'est guère contesté,
ce modèle n'est cependant pas encore admis de façon
unanime.
Dans ce modèle, il n'y a pas vraiment de
déplacement du proton mais liaison covalente d'un
proton en début de chaîne, transfert de liaisons
covalente et électrostatique tout le long de la
chaîne et libération d'un proton en fin de
chaîne par rupture de liaison covalente. En terme de
bilan il y a changement d'orientation de la chaîne de
molécules d'eau à la suite du "transfert" d'un
proton.
Cette conductivité est actuellement une piste majeure de
recherche en biochimie, notamment dans l'activité
enzymatique (les molécules d'eau de la surface des
enzymes, réunies en chaînes conductrices de protons
à partir d'un certain seuil d'hydratation, seraient
responsables, même sans aucune déformation de la
molécule enzymatique, des changements d'activité) ou
dans le couplage entre pompes et canaux (le transfert de
protons associé au fonctionnement d'enzymes hydrolysant
l'ATP pourraient être à l'origine non pas d'un
gradient d'H+ mais de courants protoniques
microlocalisés qui donneraient un autre sens à
la théorie dite chimioosmotique...) ou encore dans le
transfert de signaux ioniques (la transduction du signal
calcium souvent présentée comme le résultat
d'une cascade de réactions de
phosphorylations-déphosphorylations le long d'une
chaîne de kinases et de phosphatases, pourrait aussi
résulter de l'enchaînement de proche en proche
d'oscillations de molécules chélatrices entre un
état où le calcium est masqué et un
état où le calcium est libéré: la
propagation du signal se faisant alors sans déplacement de
calcium; des courants protoniques entre chaînes de
molécules d'eau interfaciales le long d'une chaîne de
macromolécules, pourraient aussi intervenir; on
connaît ainsi une "onde" protonique qui semble suivre l'onde
calcium en décrivant des cercles concentriques autour du
point de contact du spermatozoïde avec l'ovocyte d'amphibien
lors de la fécondation).
- l'eau interfaciale, de par ses propriétés
physiques, peut contrôler la composition chimique
de la cellule: c'est par exemple l'exclusion
sélective des ions.
L'eau interfaciale, polarisée, exclue d'autant plus
fortement les ions qu'ils génèrent un champ
électrique élevé (selon la série de
Hofmeister). Donc à taille égale ce sont les plus
chargés qui sont les plus exclus et à charge
égale, ce sont les plus petits qui sont les plus exclus.
poids atomique croissant
>>>>>>>
et / ou charge électrique
décroissante
<<<<<<<<<<<<
diamètre décroissant à
l'état hydraté
<<<<<<<<<<<<<<<<<<<<<<
|
|
Mg2+ <
Ca2+ < Sr2+ < Ba
2+ < < Li+ <
Na+
<
K+
< Rb+ <
Cs+
|
|
citrate3- <
tartrate2- < SO42-
< < Cl- < NO3-
< ClO3- < I-
< SCN-
|
|
ions cosmotropes
(du grec cosmos = ordre - structurent l'eau;
en anglais kosmotropes)
de gros diamètre - solvatés - rendent l'eau
interfaciale encore plus structurée
plus exclus de la
cellule
ions chaotropes
(du grec chaos = désordre -
déstructurent l'eau; en anglais
chaotropes)
de petit diamètre - peu solvatés - solubles
dans l'eau interfaciale qu'ils déstructurent
moins exclus de la
cellule
|
Série de Hofmeister : cations et anions
Les ions chaotropes, à la différence des
ions cosmotropes circulent facilement dans les couches
d'eau qu'ils déstructurent. C'est une raison avancée
pour expliquer que le potassium, contrairement au sodium n'est pas
exclu de la cellule. Certains auteurs estiment que la plus forte
concentration en potassium du hyaloplasme par rapport au sodium
est une conséquence directe du pouvoir solvant
sélectif de l'eau interfaciale et non le résultat
d'une activité membranaire. On a même montré
expérimentalement que des cellules, dont les membranes ont
été perméabilisées par des
détergents, et placées dans des milieux riches en
sodium, calcium et magnésium, ne s'enrichissaient pas
notablement en ces ions (alors que leur concentration ionique est
faible pour ces ions). Par contre, ces cellules à membrane
perméabilisée, placées dans un milieu riche
en potassium (mais moins riche que la cellule), perdent rapidement
leur potassium.
- les déplacements d'eau depuis la seconde couche
d'hydratation vers la première couche d'eau liée
(mécanisme de micro-osmose)
s'accompagnent de changements de
densité, donc de volume, et induisent alors des
effets mécaniques.
Le rôle de ces changements de densité dans les
moteurs moléculaires a fait l'objet de nombreuses
recherches. On a notamment proposé (voir
références d'Oplatka, in Mentré, 2002 p
100-101) l'existence de minijets d'eau à l'interface
des molécules d'actine et de myosine associées dans
les myofilaments des cellules musculaires. Le glissement relatif
entre les myofilaments serait du à la force
mécanique de l'eau déstructurée
projetée sous forme de minijets, ce qui a fait comparer le
mécanisme de la contraction musculaire ainsi
revisité à un moteur alternatif à vapeur.
Le flagelle bactérien en rotation rapide (40 à 60
tours par seconde) et l'ATPsynthase, fonctionneraient de
même comme des turbines à eau. Des minijets seraient
dirigés sur des minirotors et les feraient tourner.
Les effets mécaniques des changements de densité de
l'eau structurée pourraient aussi rendre compte de la
transduction de signaux notamment depuis la membrane
jusqu'aux structures internes de la cellule par le biais du
cytosquelette. Une déformation de la membrane cellulaire
peut modifier par contrainte mécanique la structure de
l'eau structurée au voisinage de cette membrane. Cette
modification pourrait ensuite se transmettre,
mécaniquement, de proche en proche, au sein de l'eau
d'hydratation des éléments du cytosquelette que l'on
sait se polymériser/dépolymériser à
grande vitesse. On a ainsi mis récemment en évidence
que 10 à 12 molécules d'eau s'associent à la
molécule d'actine au cours de son étape de
polymérisation.
Remarque personnelle 1:
Cette sensibilité de l'eau structurée
située dans le cytoplasme cortical de la cellule à
des stimuli mécaniques, et surtout sa capacité
à transmettre ce signal mécanique le long du
cytosquelette puis finalement à réaliser la
transduction du signal mécanique en un signal
chimique par le biais de la micro-osmose (un changement de
pression et donc de densité de l'eau interfaciale peut
induire la stabilisation d'éléments chimiques
cosmotropes ou l'exclusion d'éléments chaotropes...
voir paragraphe ci-dessus), pourrait rendre compte de la
sensibilité de nombreuses cellules nerveuses, plus ou moins
modifiées, à la pression (que l'on nomme
barorécepteurs).
Remarque personnelle 2:
Cette approche des systèmes protéiques motiles
à base d'actine dont le mécanisme de contraction
serait intimement lié aux propriétés
mécaniques de l'eau structurée semble être une
piste prometteuse de recherche. L'actine est
l'élément motile essentiel du cytoplasme cortical
qui est le siège de la cytocinèse (division du
cytoplasme) pour toute cellule eucaryote (voir ci-dessous). Le
rôle du cytosquelette cortical n'est pas non plus à
négliger dans la contraction musculaire d'une fibre
striée (les microphotographies avec coloration des
filaments d'actine montrent un réseau extrêmement
riche dans ces cellules, en plus de myofilaments classiquement
présentés. C'est encore l'actine qui peut être
à l'origine de la sensibilité à la pression
d'une villosité, si on invoque ce mécanisme de
couplage avec l'eau structurée.
- paradoxalement, l'eau de la cellule
vivante surpeuplée de macromolécules devient
rare, utilisée de façon compétitive
par les macromolécules, son accessibilité
contrôlant les activités des
biomolécules.
Un excès d'eau impliquerait un désordre par manque de
structuration. Un (léger) déficit d'eau entretient la
possibilité d'un contrôle par facteur limitant. Pour
étudier cette hypothèse P. Mentré et G. Hui Bon
Hoa ont étudié des biomolécules sous haute
pression hydrostatique (HPH), facteur connu pour favoriser une forte
hydratation de ces molécules et donc une configuration qui
maximise l'eau interfaciale, plus dense (d'après la loi de Le
Chatelier). Les résultats obtenus sur des cellules eucaryotes
- qui semblent pouvoir supporter sans problème des pressions
atteignant 1000 à 1200 bars (soit 100 à 120 MPa)
pendant une dizaine de minutes - sont époustouflants :
cytosquelette, complexes antigène-anticorps, ADN-facteurs,
etc., se dissocient sous des HP de quelques dizaines de MPa et cet
effet est réversible. Le contrôle des assemblages
biomoléculaires par l'eau est en voie d'exploration.
2ème barrière
entrouverte: l'unité fonctionnelle cellulaire ne vient pas
d'une chimie du hasard (les lois probabilistes des milieux
homogènes) mais d'une biochimie qui pourrait être
fondée sur la finalité:
la vie cellulaire est un travail orienté
vers une fin biologique.
Ces éléments sont inspirés du
chapitre 12 (L'eau, une interface hétérogène
et dynamique des macromolécules) du livre de P.
Mentré (L'eau dans la cellule, Pascale Mentré,
1995, Masson) mais la formulation et le développement des
idées déforment peut-être la pensée de cet
auteur auquel je renvoie.
La vision de la cellule vivante, abordée à partir de
la question de l'eau, telle qu'elle est présentée
ci-dessus, peut déboucher sur une autre compréhension
du métabolisme cellulaire que l'on pourrait qualifier de
finaliste - au sens de téléologique (voir au
sujet de ces termes la discussion dans le cours
de terminale) - par opposition à la vision
statistique de la biochimie classique.
|
Vivre = une chimie
ou
un travail ?
Vu de très loin, avec une résolution
inférieure à la distance entre les
éléments d'un système
macromoléculaire (que l'on pourrait
chiffrer à quelques dizaines ou centaines de
nanomètres) le cytosol est homogène
chimiquement.
Mais si l'on s'approche des éléments
moléculaires (à une distance
inférieure à la dizaine de
nanomètres)
l'hétérogénéité
saute aux yeux et il semble nécessaire de changer de
modèle.
Prenons l'exemple d'un système
métabolique (ou chaîne
métabolique) théorique où un substrat
est transformé en un produit. Approchons-nous du
système au-delà de la résolution du
microscope et intéressons-nous aux molécules
qui le composent: en voici deux représentations:
|
|
modèle "soupe*"
chaîne métabolique
statistique
|
modèle "machinerie haute
technologie*"
chaîne métabolique
finaliste
|
|
* termes empruntés à P.
Mentré: Organization and properties of water in cell
system, in Integrated Plant Systems, H. Greppin et al.,
eds., 2000, University of Geneva, pp 3-22
|
|
substrat ---->
|
système métabolique
statistiquement homogène
compris comme
- un ensemble statistiquement déterminé de
molécules en solution aqueuse (non
dénombrables) dont le comportement dépend
des:
- concentrations (et donc d'une
répartition supposée statistiquement
homogène) des espèces en solution et
de
- réactions à l'équilibre
(et donc une vitesse moyenne (statistique)
de réaction dépendant des paramètres
physiques et des concentrations...).
|
---> produit
|
substrat ----->
|
système métabolique
hétérogène
composé de:
- molécules dénombrables (tant de
molécules de telle enzyme, tant de molécules
de telle autre enzyme) et donc considérées
comme des molécules individuelles, à un
emplacement donné (environnement particulier), avec
une fonction donnée et des paramètres
individuels (qui peuvent varier selon les
molécules)
- ayant un mobile commun déterminé par
UN SEUL CHEMIN possible (ou
quelques-uns) pour aller du point d'entrée au
point de sortie ("mobile" au sens
étymologique (mécanique) de rouage pris
ici selon son sens analogique dérivé de
fin).
|
---> produit
|
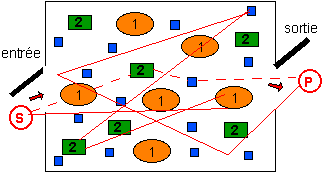
Un système contenant un très grand nombre
de molécules (le nombre
d'exemplaires de chacune des trois espèces
moléculaires est bien inférieur sur ce
schéma simpliste à ce qu'il devrait être
pour que le modèle statistique soit valide
étant donné qu'il nécessite la
diffusion aléatoire des molécules entre les
éléments du système). Les
molécules appartiennent à 3 espèces
chimiques (1,2 et 3 (en bleu)) en solution
(homogène). Un substrat S donne par réaction
métabolique un produit P. Deux chemins sont
représentés qui passe successivement par les 3
espèces moléculaires: le chemin le plus court
(en pointillés) et un des chemins les plus
longs.
|
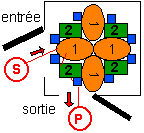
Trois espèces chimiques (1, 2 et 3 (en bleu))
groupées en un système
hétérogène à 4+4+8
éléments. Le substrat S est
métabolisé en un produit P.
Remarque:
il faut bien noter ce que cette présentation a de
nouveau, ce qui n'est peut-être pas si clair en
regardant ce schéma simpliste. En effet, les
complexes métaboliques sont connus depuis fort
longtemps mais on s'était contenté, à
ma connaissance, de signaler qu'ils augmentaient les
vitesses et les rendements des réactions
métaboliques. Dans une vision individualiste
et finaliste le chemin métabolique est une
conséquence structurale et fonctionnelle de ces
assemblages que l'on peut décrire non plus en terme
d'ordre mais de fin (but): tel système pour telle
fonction. Cette démarche était
déjà employée pour des assemblages
moléculaires comme un ribosome ou une molécule
d'hémoglobine (HbO8), notamment lorsque l'on parlait
d'autonomie ou d'autoassemblage en renvoyant implicitement
à un moteur chimique; il s'agit ici de
l'étendre à TOUTES les structures du
vivant.
|
La finalité peut rebuter quelques-uns des lecteurs
qui peuvent par exemple préférer parler tout simplement
de fonction. Le terme de fonction a probablement une
origine biologique et sociologique (cette idée
n'a été soutenue que tardivement par René Thom;
voir Analyse sémantique d'un mot polysémique: la
fonction, 1993, Séminaires de la Société de
Biologie Théorique, année 1992-1993, Acta
Biotheoretica, 42 ), puis qui a été
formalisée en mathématique (dans le
plan, la fonction (numérique) d'une variable (x) est
représentée par un graphe y=f (x); en
mathématique la représentation d'une relation de
quantité (avec des variables numériques) ou de
qualité entre deux ensembles d'objets est défini par le
terme moderne d'application (Fréchet, 1909))
, et enfin est retournée dans le domaine social et dans
celui de la biologie. Voir aussi l'extrait
de Paraboles et catastrophes de
René Thom dans le cours de 1èreS. Mais pour un
système vivant on ne peut pas parler de fonction, au sens
strictement biologique, au niveau local (fonction alcool ou
déshydrogénase...). Il est préférable de
garder le terme de fonction pour des phénomènes
physiologiques globaux comme l'homéostasie par exemple
(voir cours
de 1èresS).
En août 2005, à l'aide du travail de René
Thom, il m'apparaît clairement que la différence
fondamentale entre ces deux modèles repose sur le contraste
entre un ensemble discret de
molécules (dont on cherche à comprendre les
interactions par le biais d'une analyse statistique) et une fonction
biologique continue (qui ne peut donc
être issue des éléments discrets et dont
l'origine doit se trouver ailleurs). Pour des détails voir une
page en travaux sur le continu en
SVT.
à suivre...