 http://biology.clc.uc.edu/fankhauser/ Labs/Genetics/ Root_tip_chromosomes/ chromosomes/ meta_P1161783.JPG |
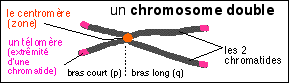
Les chromosomes sont de gros "corps colorables" allongés (de quelques micromètres de long pour 0,7 µm de diamètre de chromatide) qui apparaissent dans le noyau des cellules eucaryotes lors de la division cellulaire. |
chromo (en grec) = couleur |
||||||||
|
|
Remarque: certains chromosomes sont sphériques et non pas allongés. |
|||||||||
|
On devrait vraiment
éviter de parler de chromosomes pour le
matériel génétique de la cellule
procaryote (il vaut mieux parler de nucléoïde
pour désigner la structure qui est partiellement
visible au ME mais pas au MO !!! -
voir cours de
1èreS) ainsi
que pour le matériel génétique des
organites de la cellule eucaryote (mitochondries et
chloroplastes).
|
 |
|||||||||
|
1. Les chromosomes sont colorables
|
||||||||||
|
Par des détails voir ancien cours de spécialité de TS |
Le noyau interphasique (au repos) contient une substance qui peut être colorée très facilement par des colorants basiques : la chromatine. |
|||||||||
|
La chromatine a une forte affinité
pour de très nombreux colorants: |
La chromatine forme habituellement un enchevêtrement de fibres (en "réseau") mais peut aussi prendre un aspect plus condensé (les zones denses sont appelées "chromocentres"). On peut la mettre en évidence par coloration de Feulgen ou par le test de Brachet (voir ci-contre) |
 voir image sur le site Bmedia de jussieu Coloration de Feulgen
On réalise cette coloration sur
des coupes histologiques ou sur des cellules vivantes mais
elle est létale (elle provoque la mort de la
cellule). La chromatine se colore en rouge violacé alors que le cytoplasme, le nucléoplasme et le nucléole restent incolores. C'est la présence du désoxyribose de l'ADN qui est ainsi mise en évidence dans la chromatine. |
Test de Brachet
: Chez une cellule eucaryote témoin
le nucléole et le cytoplasme sont colorés en
rouge et la chromatine en vert. Cette affinité
différentielle pour les deux colorants basiques n'est
pas très clairement expliquée à ma
connaissance. Après action de la ribonucléase,
la coloration rouge a disparue du cytoplasme et du
nucléole alors que la chromatine reste colorée
en vert. On peut mettre en relation directe la coloration
rouge avec la synthèse d'ARN dans différentes
cellules. Plus une cellule réalise des
synthèses protéiques plus elle possède
d'ARN.
|
|||||||
|
Le ou les nucléoles (nombre presque fixe chez une espèce donnée), sont des inclusions très facilement colorables par les colorants basiques, la plupart du temps sphériques et d'aspect homogène. |
|
|||||||||
|
pour l'ADN voir le TD acides nucléiques |
|
|||||||||
|
La chromatine contient environ 50% d'ADN et 50% de protéines. |
Dans les coupes ultrafines
observées au microscope électronique (MET) la
technique est bien différente de la coloration
réalisée pour le MO. Mais on garde le terme de
chromatine pour désigner la substance
osmiophile (qui fixe le tétroxyde d'Osmium, le
colorant opaque aux électrons utilisé) MAIS ce
n'est pas forcément la même structure que la
chromatine vue au MO. |
Les histones (protéines basiques) structurent l'ADN dans la chromatine , c'est là qu'on les étudie. Mais on pense qu'elles formeraient aussi le squelette du chromosome, au moins partiellement. |
Les protéines acides
(non-histones) sont plus ou moins étroitement
liées au complexe ADN-histones. Dans la plupart des
espèces étudiées, on dénombre
plus d'une centaine de variétés
moléculaires comprenant une trentaine de
protéines intervenant dans la morphologie du
chromosome et des protéines impliquées dans la
régulation de la synthèse de l'ADN et de
l'ARN. Leur effectif varie donc au cours du cycle cellulaire
et dans les différents tissus ; leur rôle
exact reste encore bien souvent mal défini.
(EU "chromosomes")
|
|||||||
|
N.B. Le nucléoïde bactérien est pratiquement composé d'ADN pur (la teneur en protéines est très faible) chez un grand nombre de bactéries. C'est aussi le cas du noyau chez certains eucaryotes comme les Dinoflagellés. le noyau de certains spermatozoïdes comme chez certains Crustacés ou encore l'ADN mitochondrial des protozoaires agent de la maladie du sommeil (Trypanosome). Dans ces cas l'ADN se présente sous la forme de filaments formant des agrégats plus ou moins ordonnés (E.U. article "Cristaux liquides : phases mésomorphes biologiques" Yves Bouligand). |
||||||||||
|
Lors de la mitose la chromatine disparaît et les chromosomes apparaissent: ils sont probablement issus de la condensation de la chromatine |
||||||||||
|
Lors d'une mitose, le nucléoplasme ne contient plus de chromatine (colorable) mais tout l'ADN ne se condense pas en chromosomes. Le ou les nucléoles restent souvent séparés et sont plus ou moins répartis entre les cellules filles. Il existe aussi une certaine quantité d'ADN extrachromosomique chez certaines espèces. Il semble que cet ADN contienne surtout des gènes codant pour des ARN ribosomiaux. |
Les chromosomes sont aussi facilement colorables que la chromatine avec des colorants basiques. Les chromosomes contiennent plus de protéines que d'ADN (1/3 d'ADN et 2/3 de protéines). Les protéines se répartiraient pour moitié en protéines histones et pour moitié en non-histones. La technique qui consiste à colorer les chromosomes puis à prendre un photo lorsqu'ils sont étalés (en vue polaire en métaphase de mitose, la cellule étant éclatée) pour les dénombrer, est appelée technique de caryotypage. On dit que l'on réalise un caryotype. |
Le protocole du caryotypage comprend
: |
||||||||
|
Un extrait de film du CERIMES présentant la formation des chromosomes dans un noyau dans un tissu de l'ovule de la plante (endosperme) : http://www.cerimes.education.fr/index.php?page=bi_admin&op1=plan,,,99,,,,,1,4050
Références de
l'article: Three-Dimensional
Maps of All Chromosomes in Human Male Fibroblast Nuclei and
Prometaphase Rosettes |
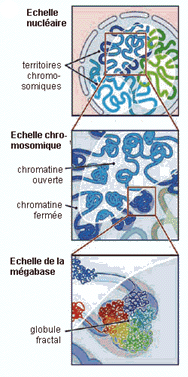
Dans le film ci-contre (<-) il est particulièrement clair que ce sont des territoires nucléaires qui donnent naissance aux chromosomes. Pour des données plus récentes on peut voir par exemple un article en anglais accessible sur le net librement (licence Creative Commons Attribution) est à l'adresse http://biology.plosjournals.org/archive/1545-7885/3/5/pdf/10.1371_journal.pbio.0030157-S.pdf dont voici une figure (Fig. 1-D - réf: 10.1371/journal.pbio.0030157.g001) particulièrement accrocheuse: 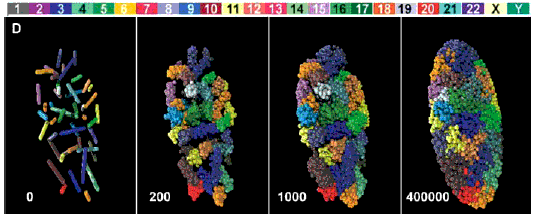 Le positionnement des 46 chromosomes (dans un état de décondensation croissant de la gauche vers la droite) d'un fibroblaste humain en reconstitution 3D simulant les résultats obtenus par microscopie (voir l'article pour des précisions) J'aime beaucoup la légende ajoutée par La Recherche (brève du n°388, juillet-août 2005, 15): Paysage nucléaire, qui rappelle le paysage épigénétique de Waddington (voir page annexe). |
|||||||||
|
Une référence (accès restreint): Cytogénétique humaine. De 1956 à 2006, R. Berger, Pathologie Biologie 55 (2007) 1-12 |
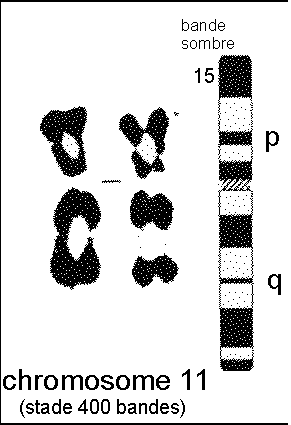
* La coloration la plus ancienne pour les chromosomes , réalisée depuis les années 1970-1972, est celle qui permet de mettre en évidence des bandes (méthode du banding en anglais). Des bandes claires (non colorées) alternent avec des bandes sombres (colorées). On obtient jusqu'à 300 bandes pour l'ensemble de tous les chromosomes. On distingue 4 techniques : |
|||||||||
|
|
(G-banding) |
(R = reverse) |
(C-banding) |
|||||||
|
Caspersson, utilise la quinacrine qui permet, en lumière fluorescente, de distinguer sur chaque chromosome des régions qui se subdivisent en bandes, caractéristiques d'un chromosome donné |
Les chromosomes sont soumis à
différents traitements (action de la trypsine ou de
tampon SSC à 65°C -
Seabright,
Summer, voir références
dans l'article de l'APBG cité plus
haut) puis colorés avec le
colorant de Giemsa. On observe des bandes , assez
voisines des bandes Q : certaines prenant fortement le
colorant (G), d'autre le prenant très peu. |
Les bandes R sont obtenues par dénaturation thermique à 85°C (Dutrillaux et Lejeune, voir références dans l'APBG, La fluorescence in situ par hybridation (FISH), M. Lessard, I. Luquet, M.-J. Lebris et A. Herry, Biologie-Géologie (Bulletin de l'APBG), 3-1996, 531-542) et marquent les télomères en sombre à l'inverse des bandes Q et G, avec lesquelles les télomères sont pâles |
Le traitement utilisé permet de rendre fortement colorable l'hétérochromatine du centromère de tous les chromosomes. |
|||||||
|
|
On s'accorde maintenant à penser
que non seulement les bandes chromosomiques ne sont pas des
artefacts, mais qu'elles correspondent à des
caractéristiques précises
résumées ci-dessous : Riches en AT Riches en GC Remarque : |
|||||||||
|
* Le caryotype de l'homme comprend 23 paires de chromosomes |
||||||||||
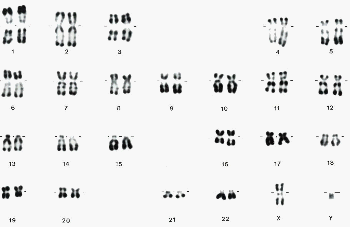 caryotype (faible nombre de bandes) classé d'un homme (http://www.ac-reims.fr/datice/svt/docsacad/caryotype/caryo2/caryonormal.htm) |
22 paires d'autosomes (chromosomes qui ont deux paires ayant LES MÊMES BANDES): A (paires 1, 2 et 3), chaque chromosome pouvant être reconnu par les bandes colorées 1 paire de gonosomes (paire 23): ou chromosomes sexuels ; ils ont les mêmes bandes chez la femme (XX) mais ni la même taille ni les mêmes bandes chez l'homme (XY) |
|||||||||
|
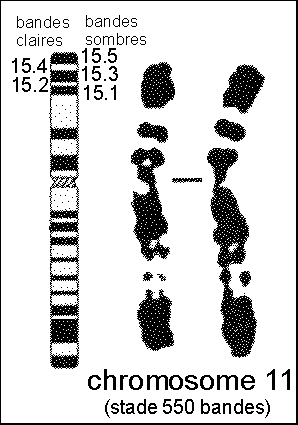
* Depuis 1976 on utilise une technique de coloration en bandes à haute résolution. |
||||||||||
|
On passe de 300 à 600-800 bandes par caryotype haploïde. Du point de vue physique c'est un étirement du chromosome qui permet de séparer les bandes. Mais là encore, on ne connaît rien de plus sur la signification de ces bandes. Ces techniques permettent des découvertes de micro-délétions (par exemple la microdélétion del (13) q13-14 associée au rétinoblastome). Une bande moyenne correspond à 5 à 10 millions de paires de bases. |
||||||||||
|
|
||||||||||
|
|
|
|||||||||
|
|
||||||||||
|
Un chromosome marqué par 2 sondes fluorescentes : rouge et verte (image en négatif à droite) par FISH |
* Depuis les années 1985-1990 on sait marquer chaque chromosome avec des sondes spécifiques fluorescentes de couleurs différentes (technique de la FISH: Fluorescence In Situ par Hybridation), ce qui permet de faire un dénombrement rapide automatisé. |
|||||||||
|
Pour ce rendre compte du développement de la reconnaissance automatique des caryotypes (normaux et anomalies de caryotypes) on peut visiter par exemple la page de la société Alphelys : http://www.alphelys.com/site/fr/ pBICY_Cytogenetique.htm |
On sait marquer chaque chromosome par des sondes spécifiques de certaines régions du chromosome (centromère, télomères...). On a ensuite trouvé des substances fluorescentes de différentes couleurs. Il a suffit d'associer ces sondes à ces marqueurs colorés pour obtenir un système de marquage qui se prêtait à l'automatisation. |
Couplés à des techniques classiques de coloration de bandes (banding en anglais) on a maintenant des outils commerciaux de caryotypage. |
||||||||
|
Les techniques de FISH ont grandement
évolué en 20 ans et elles sont
utilisées couplées aux analyses de bandes,
à la consultation des bases de données... et
la résolution de la cartographie chromosomique est en
passe de rejoindre celle obtenue par génie
génétique sur des molécules d'ADN
extraites.
|
||||||||||
|
Une référence (accès restreint): Cytogénétique humaine. De 1956 à 2006, R. Berger, Pathologie Biologie 55 (2007) 1-12 |
* De nombreux sites fragiles ont été recensés dans les chromosomes « Les sites fragiles correspondent à points des lacunes (manque de coloration) et/ou des cassures chromosomiques non aléatoires visibles sous certaines conditions. Le site fragile situé sur le chromosome X en Xq27.3 (FRAXA, du groupe sensible aux folates) a été l'un des premiers étudiés dans la mesure où il est associé à un syndrome particulier comportant un retard mental, le syndrome de l'X fragile, dans le sexe masculin. ... On peut considérer actuellement qu'il n'a pas été possible de prouver une relation générale de cause à effet entre points de cassure des maladies acquises et localisation des sites fragiles. Cependant, les sites fragiles sont des régions préférentielles d'insertion virale, et peuvent servir d'ancrage pour un processus d'amplification génomique, par exemple dans les tumeurs malignes. Les conséquences des cassures au niveau des sites fragiles restent donc un sujet de recherche pour les cytogénéticiens. » (in Berger 2007). |
|||||||||
|
|
||||||||||
|
2. La structure des chromosomes n'est pas connue avec certitude |
||||||||||
|
|
||||||||||
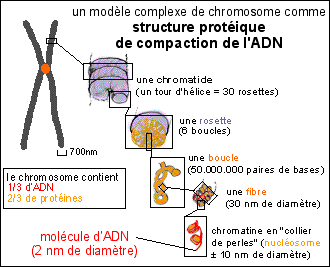 |
Le modèle le plus habituel présente le chromosome comme une structure de compaction d'une UNIQUE gigantesque molécule d'ADN enroulée autour de petits cylindres (les nucléosomes) eux-mêmes compactés en une fibre, puis les fibres torsadées en boucles, les boucles associées en rosettes pour finir par donner une fibre chromatidique d'environ 700 nm de diamètre, fortement colorable. Des électronographies de structures de chromosomes "éclatées" sont données pour "preuve" de la présence d'une unique molécule. |
Les modèles "alternatifs" de la structure chromosomique proposent une structure répétitive de petits ADN en boucles. |
||||||||
|
Les
éléments qui suivent sont placés ici
dans le but de souligner qu'ils dépendent fortement
de l'interprétation du chromosome comme structure de
compaction d'une molécule d'ADN
par chromatide. Dans le cadre d'un autre modèle les
données du cycle cellulaire seraient à
reprendre différemment. Quelques arguments sont
donnés dans l'article chromosomes de l'Encyclopedia
Universalis (v 12) mais ils ne seront pas repris ici ; il
suffit à un élève de savoir que la
présence d'une seule molécule d'ADN n'est pas
une certitude.
|
||||||||||
|
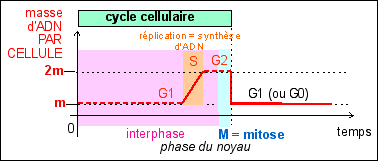
Avant chaque division les molécules d'ADN de la cellule eucaryote doivent être dupliquées (c'est la réplication). Cette augmentation de la quantité d'ADN par cellule a lieu lors de l'interphase (phase de repos du noyau). |
 |
|||||||||
|
|
||||||||||
|
Lors de la mitose (phase de division du noyau) les chromosomes apparaissent. |
coloration bleu de
méthylène, |
|||||||||
|
On peut donc relier les
différentes phases du noyau au cours du cycle
cellulaire (temps séparant deux divisions
successives) aux mécanismes moléculaires qui
seront vus en
1èreS. Les noms des phases
de l'interphase (G1, S, G2) ne sont pas à
mémoriser en seconde.
|
||||||||||
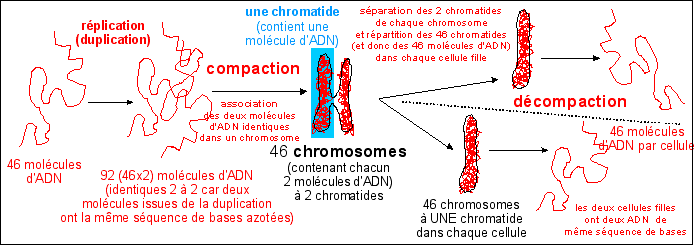 Dans le cas d'une cellule humaine, les 46 chromosomes sont donc associés à 46 immenses molécules d'ADN, dont la longueur totale atteint environ 2m (ce qui fait environ 2 cm d'ADN par chromosome). |
||||||||||
|
|
||||||||||
|
3. Les chromosomes sont déplacés dans la cellule grâce à des microtubules |
non traité à ce niveau d'enseignement |
|||||||||
|
|
||||||||||
|
4. Les chromosomes se scindent en deux à l'anaphase |
non traité à ce niveau d'enseignement |
|||||||||
|
|
||||||||||
|
5. Les chromosomes s'accolent à la prophase de la première division de méiose et forment des complexes synaptonémaux |
non traité à ce niveau d'enseignement |
|||||||||
|
|
||||||||||
|
6. Le rôle des chromosomes est encore mystérieux |
||||||||||
|
|
||||||||||
|
Deux rôles classiques : |
* les chromosomes sont le support des particules héréditaires (gènes héréditaires) dans la théorie mendelienne-morganienne de l'hérédité |
|||||||||
|
* les chromosomes ne sont que le
système d'emballage de l'ADN qui est la
molécule-clé de la compréhension du
fonctionnement chimique de la cellule (le gène
moléculaire comme unité fonctionnelle de la
machinerie chimique du vivant)
|
||||||||||
|
Des pistes cytologiques moins conformistes : |
* les chromosomes sont de vraies structures spécifiques des cellules eucaryotes qui portent en elles la compréhension de la division cellulaire (explications structurales de la mitose...; compréhension des chromosomes comme des éléments spectraux, figures stables de la dynamique de division...) |
|||||||||
|
|
||||||||||
|
extrait de René Thom,
Stabilité structurelle et morphogenèse;
essai d'une théorie générale des
modèles,
stabilité.pdf,
1968: |
||||||||||
|
Je suis bien conscient que les arguments fournis ici sont bien insuffisants si l'on voulait étayer une théorie, ce qui n'est pas mon rôle. Je crois que mon rôle d'enseignant recouvre aussi la charge d'ouvrir les jeunes esprits à une compréhension moins réductrice que ce qui m'a été enseigné. |
|
|||||||||
|
* les chromosomes gardent les traces
des métabolismes de la cellule; ils sont donc
modifiables par la cellule elle-même; ils pourraient
consituer une mémoire dynamique de la
cellule. |
||||||||||
|
|
||||||||||
|
voir cours de seconde §2.4.2.6 et §2.4.7 |
- le paysage nucléaire : les chromosomes se forment au centre d'un territoire nucléaire spécifique. De là à penser qu'ils puissent conserver (et transmettre ?) une organisation spatiale, ce qui est mathématiquement possible, il y a un fossé que peut combler l'imagination nécessaire à tout chercheur (voir ci-dessus ou cours de seconde §2.4.7). |
|||||||||
|
|
||||||||||
|
Données complémentaires sur la page sur les mutations |
- les anomalies
chromosomiques : on connaît des anomalies
chromosomiques dans certains groupes de cellules
cancéreuses que l'on bien du mal à expliquer
comme résultant d'une contamination par des cellules
étrangères: le dogme de l'identité
caryotypique de toutes les cellules d'un organisme est
fissuré (cours
de seconde
§2.4.6). |
|||||||||
|
« ... les cytogénéticiens ont montré que bon nombre d'anomalies chromosomiques étaient non aléatoires dans la mesure où certaines d'entre elles, surtout des anomalies de structure (translocation, inversion), étaient associées à des types plus ou moins spécifiques de proliférations, et que d'autres, surtout des anomalies de nombre étaient plus fréquentes que ne le voudrait le hasard. Les premières ont été souvent qualifiées de « primaires » pour indiquer le fait qu'elles sont associées à un événement causal et les secondes « secondaires » car plus inconstamment associées aux premières ou pouvant survenir après elles. Ces anomalies secondaires peuvent être présentes dès le premier examen cytogénétique, mais elles peuvent aussi survenir au cours du suivi du patient et définir alors une évolution clonale du caryotype. Plusieurs centaines d'anomalies récurrentes des cancers et hémopathies malignes sont maintenant connues (Mitelman database of chromosome aberrations in cancer : http://cgap.nci.nih.gov/Chromosomes/Mitelman ; Atlas of genetics and cytogenetics in oncology and haematology : http://atlasgeneticsoncology.org// ) et bon nombre d'entre elles ont pu être caractérisées au plan moléculaire». (Berger, 2007) |
||||||||||
|
|
||||||||||
|
Une référence (accès restreint): Cytogénétique humaine. De 1956 à 2006, R. Berger, Pathologie Biologie 55 (2007) 1-12 |
« Plus récemment la mise à disposition de sondes télomériques spécifiques des régions subtélomériques de chacun des chromosomes a permis de rechercher systématiquement des remaniements cryptiques ou subtils chez des enfants retardés mentaux. Le pourcentage de détection de telles anomalies est variable selon les études faites et peut atteindre environ 7 % dans certaines d'entre elles. Plusieurs méthodes ou combinaisons de méthodes peuvent être employées pour la recherche de ces anomalies comme la FISH en multicouleur sur chromosomes métaphasiques qui peut détecter des translocations, ou le couplage avec une hybridation génomique comparative sur chromosomes ou même puces d'ADN mettant en évidence des remaniements déséquilibrés. L'existence de ces remaniements incite à rechercher leurs conséquences sur l'expression des gènes normalement situés dans les régions subtélomériques des chromosomes impliqués, dans la mesure où la proximité de l'hétérochromatine télomérique peut altérer leur expression. Il s'agit là d'une nouvelle approche possible de l'analyse des conséquences des remaniements chromosomiques sur l'expression génique qui peut être mise en parallèle avec les altérations attendues de l'expression de gènes rapprochés de régions d'hétérochromatine constitutive dans les remaniements péricentromériques». (Berger, 2007) |
|||||||||
|
|
||||||||||
|
|
||||
|
Une comparaison (limitée): |
 x1/2
x1/2 |
 x10.000 x10.000 |
||
|
|
|
|||
|
image La recherche et image MEB traité |
|
|||
|
structure |
connecteur |
système d'accrochage au squelette cellulaire (centromère) |
||
|
contrôleur, système d'écriture-lecture |
protéines du squelette du chromosome contrôlant la forme du chromosome, l'enroulement de l'ADN... mais il n'existe pas de système CONNU de lecture de l'ADN, et encore moins d'écriture au sein du chromosome, voir plus bas |
|||
|
|
||||
|
nature physique de la mémoire |
transistors à effet de champ (stockant des bits sous forme d'électrons piégés dans des grilles de silicium). Des défauts sont possibles. |
séquence de nucléotides de l'ADN (4 nucléotides différant par une base azotée (A, T, C, G) avec des modifications possibles: méthylations, bases anormales ou mal appariées....) |
||
|
|
||||
|
contenu de la mémoire |
+ des fichiers (avec une en-tête qui donne la nature du fichier et son mode de lecture) + les zones où ne sont pas stockées les fichiers sont mises à 0 ou à 1 |
+ des séquences (répétitives pour la plupart) dont on ignore le sens (90% de la mémoire). Pour les partisans d'une théorie de l'information génétique "dure", ces séquences seraient d'anciens fragments inutilisés ("ADN poubelle"). pour les tenants d'une vision plus physique, c'est la masse d'ADN qui important davantage que sa séquence... + des gènes (10% de la mémoire) codant pour des ARN (dont certains peuvent être traduits en protéines - environ 1%, les gènes du protéome) - un gène pourrait être comparé à un fichier (avec séquences de début et de fin; mais les gènes peuvent être morcelés ou chevauchants sur plusieurs "fichiers")...des gènes identiques en terme de fonction pourraient correspondre à deux fichiers image représentant le même objet mais pas forcément avec le même éclairage, ni le même angle de vue... (les différences seraient les différences de séquence des gènes moléculaires). |
||
|
|
||||
|
lecture-écriture |
un système électronique d'écriture/lecture applique ou mesure des tensions aux bornes de chaque microélectrode de chaque grille... un contrôleur d'entrée-sortie arrange les données selon des standards... |
TOTALEMENT
INCONNU + dans la théorie de l'information génétique fusionnée avec le néodarwinisme dans une théorie synthétique moderne de l'évolution , les gènes sont constitués de fragments de gènes ancestraux qui s'assemblent, se dupliquent et surtout "mutent" au hasard + une vraie théorie de la fabrication des gènes pourrait émerger des études physiques de la dynamique métabolique dans la cellule: l'ADN serait la trace stable des dynamiques de synthèse... |
||
|
|
||||
|
différences irréductibles |
composé artificiel La mémoire flash peut stocker des programmes exécutables mais qui doivent être transférés dans une mémoire active pour pouvoir être exécutés; ce sont donc en fait des fichiers, mais d'un type particulier. |
organite d'une cellule eucaryote vivante Pour l'instant aucun programme (ni fichier de type programme) n'a jamais été mis en évidence dans l'ADN malgré les espoirs qui avaient été ceux de la théorie de l'information génétique dans les années 1960. Tout le système de traduction des gènes du protéome (qui peuvent être transcrits en ARN traduits eux-même en protides) est extérieur au chromosome et intervient dans une phase où le chromosome a disparu. Lorsque nous aurons une idée plus précise de la façon dont la cellule construit ses chromosomes, peut-être comprendrons-nous mieux le sens de ses composants (2/3 de protéines, 1/3 d'ADN). Si l'ADN a bien un rôle de mémoire, le chromosome a peut-être un rôle spécifique (comme organite de la division). |
||
|
|
||||
|
Données sur les mémoires flash d'après L'avènement des mémoires flash, La Recherche n°375, 05-2004) |
Les mémoires flash contiennent des transistors à effet de champ (C-MOS), qui fonctionnent en prenant en compte la présence de charges électriques (et donc sans consommation électrique sauf en lecture/écriture). Sur une fine bande de silicium, appelée «canal», chargée négativement, on dépose une pastille également en silicium, baptisée grille, mais chargée positivement. On apporte des électrons à la grille en lui appliquant une tension (et le canal se ferme). Si l'on prélève des électrons à la grille le canal s'ouvre. Des électrodes à chaque extrémité du canal "lisent" l'état du canal : un courant électrique circulera, ou non, en fonction du nombre d'électrons que comporte la grille, sans pour autant modifier son contenu. Chaque transistor à effet de champ est une case mémoire et stocke 1 bit (byte). Pour une clé de 1Go (1 octet = 8 bits) il y a 8 milliards de transistors. Les premiers transistors à effet de champ remontent à 1930, mais il fallut attendre les progrès du traitement du silicium pour atteindre des coefficients d'isolement suffisants pour que les électrons ne fuient pas les grilles (actuellement la mémoire est évaluée à environ 10 ans). La clé contient aussi une électronique chargée de son écriture et de sa lecture et un contrôleur USB assure la communication entre la mémoire et l'ordinateur. |
|||
|
|
||||