|
Diviser = séparer en plusieurs parties Multiplier = reproduire, ajouter un nombre à lui-même une certain nombre de fois... Proliférer
= se multiplier (reproduire) en grand nombre
|
Il n'est pas rare que l'on confonde division et multiplication des cellules. Mais utiliser le sens précis de ces mots est préférable. La division cellulaire au sens habituel donne deux cellules à partir d'une seule (et non pas deux demi-cellules); il ne s'agit pas d'un fractionnement en sous-unités mais d'une multiplication cellulaire: chaque cellule-fille est une cellule à part entière (1->2). |
La multiplication cellulaire comprend un accroissement (d'une ou deux dimensions, du volume, de la masse - avec des phénomènes de duplication de structures ou de molécules -) puis une division qui est la séparation des différentes parties. Lorsque la multiplication est très active on parle de prolifération cellulaire. |
La division peut être binaire (1->2 = dichotome selon les racines grecques) ou multiple (1-> n, n>2). La division peut être égale ou inégale. |
|||
|
Cette partie ne traite
pas de la question de façon exhaustive (cette
question sera reprise en 1èreS) mais présente
la division cellulaire dans un contexte élargi de la
reproduction à la croissance - nutrition et à
la croissance - développement. |
||||||
|
Plan |
||||||
|
3 - La
prolifération des cellules eucaryotes peut avoir des
finalités
différentes |
||||||
|
|
||||||
|
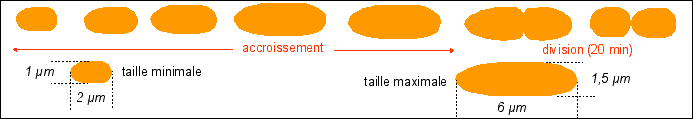
1- La
multiplication des bactéries comprend une phase de
croissance et une phase de division (division par
scissiparité)
|
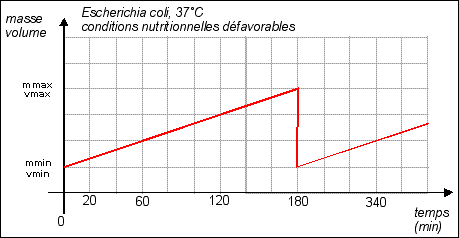
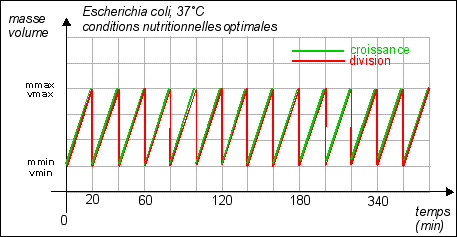
Remarque: dans cette partie les courbes sont théoriques (certaines très hypothétiques...) |
|||||||
|
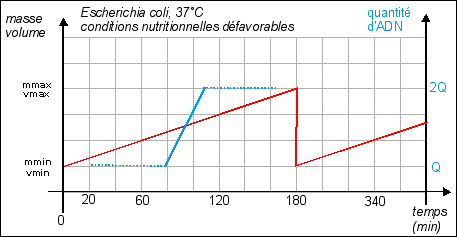
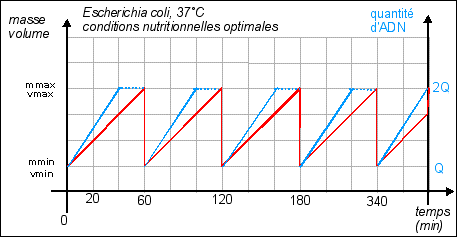
La durée de la
division est quasiment fixe (20 min), c'est la phase de
croissance qui varie selon les conditions de nutrition. Croissance =
variation de la masse (ou du volume) / temps. |
||||||||
|
scissiparité = division d'une cellule en deux par étranglement (formation d'un septum) puis scission (séparation) (ce mode de division s'oppose au bourgeonnement (excroissance d'une sphère de petite dimension) ou aux modalités de la cytodiérèse des cellules eucaryotes qui accompagnent la mitose, qui désigne plus précisément les phénomènes nucléaires) |
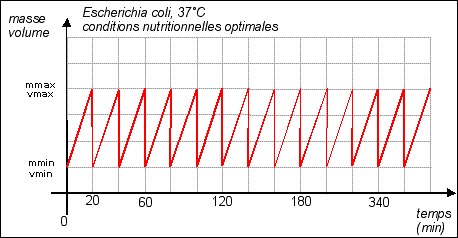 Dans cette représentation la croissance est supposée régulière (croissance constante). |
|||||||
 |
||||||||
|
|
||||||||
|
Le temps qui sépare deux
divisions successives est d'abord appelé le
cycle cellulaire ou encore le
temps de
génération |
Ces deux notions font
référence à des
phénomènes différents : |
|||||||
|
|
||||||||
|
La partie croissante de
la courbe rouge
est concomitante avec les
phénomènes de préparation de la
division |
||||||||
|
|
||||||||
|
Seuls les accroissements de la quantité d'ADN ont été représentés. La quantité d'ADN de chaque cellule est divisée par deux lors de la division cellulaire (barre verticale bleue non représentée) (Pour des détails sur la réplication de l'ADN, voir les compléments du cours de 1èreS) |
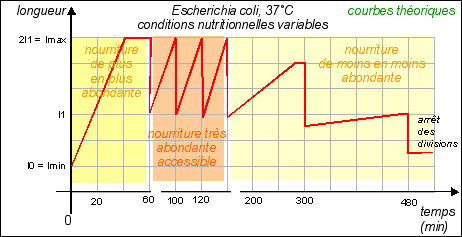
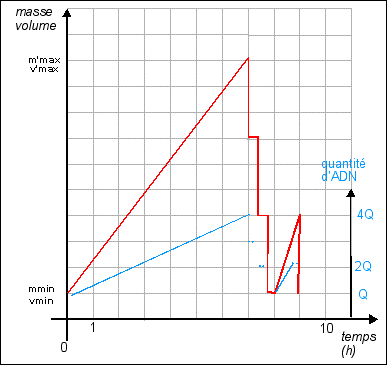
On considère habituellement que la durée de réplication de l'ADN est fixe : 40 min (quelles que soient les conditions de nutrition). Sachant qu'il faut encore qu'il s'écoule 20 min entre la fin de la réplication et la fin de la division, on atteint un temps de génération minimum de 60 min. Si l'on s'efforce de représenter l'évolution de la quantité d'ADN (que l'on superposera au schéma précédant) lorsque le temps de génération est de 180 min, il n'y a pas de problème particulier; si ce n'est que l'on ne sait pas trop où placer le début ou la fin de la réplication : Courbe pour un temps de génération de 180 min Il est encore facile de tracer cette courbe théorique dans le cas où le temps de génération est de 60 min: Courbe pour un temps de génération de 60 min Pour atteindre des durées de génération de 20 min il faut obligatoirement que croissance et division soient simultanées et surtout que l'ADN commence à se répliquer plusieurs fois au cours de ces 20 min, ce qui fait que l'on a une quantité d'ADN variable par cellule mais supérieure à 1 unité. |
|||||||
|
On considère alors que la réplication, la croissance et la division sont simultanées. Mais ce n'est pas très convaincant. |
||||||||
|
On sait maintenant que les bactéries comme Escherichia coli contiennent en permanence de 4 à 6 exemplaires de leur matériel génétique. |
||||||||
|
Hypothèse: Une solution possible serait que les chiffres des temps de génération nous soient donnés pour des populations et ne couvriraient alors que de brefs laps de temps. Pour visualiser l'évolution d'une population bactérienne , voir par exemple la page : http://lyon-sud. univ-lyon1.fr/bacterio/cours/ Croissance.html. Dans ce cas, pendant une phase de croissance lente de la population, les bactéries croissent en taille (s'allongent beaucoup) et répliquent leur ADN PLUSIEURS FOIS sans se diviser. puis pendant une seconde et brève phase exponentielle de croissance de la population, les bactéries "allongées et riches en ADN" se divisent rapidement avec une faible croissance. Dans ce cas le temps de génération minimal peut être de 20 min, mais rapidement il faut de nouveau une phase de croissance cellulaire. (voir figure, la répartition de l'ADN n'étant bien sûr pas égale entre les cellules filles) |
||||||||
|
|
||||||||
|
On remarquera que cette description de la multiplication cellulaire bactérienne ne prend pas en compte le niveau d'organisation supérieur à la cellule (la colonie). De nombreuses bactéries gardent en commun leur revêtement de sucres (glycocalix), une paroi (streptocoques en chaînettes...). On observe une véritable différenciation entre les cellules au centre de la colonie (certaines sont mortes) et celles qui sont sur les bords... Certains chercheurs comme Shapiro n'hésitent pas à affirmer que les bactéries sont des organismes pluricellulaires. L'individu étant la colonie. Les phénomènes de multiplication des cellules sont alors comparables aux phénomènes de croissance tissulaire lors du développement d'un pluricellulaire. (Pour avoir une vue des travaux de James Shapiro, voir par exemple : http://shapiro.bsd.uchicago.edu/ Shapiro98AnnRevMicro.pdf ). |
||||||||
|
Les documents cinématographiques pour les bactéries sont peu nombreux car difficiles à obtenir du fait de la petite taille des cellules (on ne peut pas prendre en film en microscopie électronique puisque l'objet observé est mort, fixé, coloré (OsO4) et observé sous vide !!!!!). Quelques petits films pris en microscopie optique avec ou sans fluorescence sont disponibles sur internet. Les films de Shapiro sont superbes : par exemple : http://shapiro.bsd.uchicago.edu/ singlecell_microcol_08_88.mov ; mais ils manquent d'explications (technique, temps séparant deux images...).
Beaucoup mieux, la croissance d'une
microcolonie de Bacillus subtilis (dont les certaines
cellules sporulent (forment des
petites spores sphériques reproductives)
et d'autres meurent...) dans le
dernier article de l'équipe Jan-Willem Veening, Eric
J. Stewart, Thomas W. Berngruber, François Taddei,
Oscar P. Kuipers et Leendert W. Hamoen
(Bet-hedging and epigenetic
inheritance in bacterial cell development, PNAS 18 mars
2008, vol 105, n°11, 4393-4398 : http://www.pnas.org/content/105/11/4393
).
On peut aussi voir le site commercial de
cells alive (http://www.cellsalive.com/
qtmovs/ecoli_mov.htm) mais c'est
à la limite de l'animation informatique.
|
||||||||
|
2 - La multiplication bactérienne peut être considérée comme une multiplication de croissance de la colonie et non une reproduction |
La multiplication bactérienne
n'est pas une reproduction |
|||||||
|
* Les cellules bactériennes vieillissent et gardent en mémoire l'âge de leur cytoplasme |
||||||||
|
* Pour le vocabulaire de René Thom, voir page sur les 4 causes d'Aristote |
1ère hypothèse: |
|||||||
|
|
||||||||
2ème hypothèse: |
||||||||
|
Pour essayer de trancher cette question une équipe française s'est efforcée de suivre les lignées cellulaires au sein d'une microcolonie bactérienne d'E. coli : |
||||||||
|
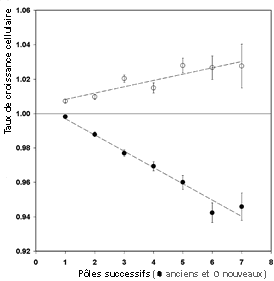
Les expériences de Eric Stewart, Richard Madden, Gregory Paul et François Taddei sont rapportées dans un article de PLOS de février 2005 (article complet en anglais disponible à l'adresse http://biology.plosjournals.org/perlserv/? request=get-toc&issn=1545-7885&volume=3&issue=2). Je vous encourage à lire les explications en français qui sont à l'adresse http://www.forumlabo.com /2006/actus/actus/ INSERM/0305bacterie.htm. Une des 96 vidéos (microscopie en fluorescence; 1 image toutes les 4 min de 0 à 160 min et 1 image toutes les 2 minutes ensuite) de la croissance d'une microcolonie qui atteint 505 cellules en 305 min: http://biology.plosjournals.org/archive/ 1545-7885/3/2/supinfo/10.1371_ journal.pbio.0030045.sv001.wmv  Une microcolonie avec reconnaissance et coloriage automatique des cellules http://biology.plosjournals. org/perlserv/ ?request= slideshow & type=figure &doi=10.1371 / journal.pbio. 0030058&id=21204 |
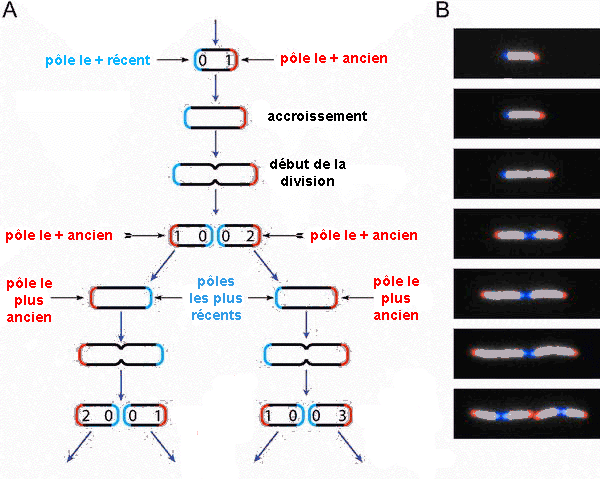
Ils ont suivi au microscope (pendant 6h
avec 94 vidéos (voir ci-contre) et sur plus de 35.000
cellules) et analysé automatiquement les images
numérisées de la croissance d'une culture de
la bactérie Escherichia coli . Ce suivi visuel
de la lignée bactérienne a été
rendu possible par la caractéristique morphologique
de la bactérie qui est en forme de bâtonnet et
qui se divise par allongement, ce qui permet de
considérer un pôle ancien et un pôle
nouveau, au niveau du sillon de division
(voir l'image: http://biology.plosjournals.org/perlserv/
?request=slideshow&type=figure&doi=10.1371/
journal.pbio.0030045&id=22668).
Les limites de cette méthode sont la formation de
plusieurs couches de cellules ou bien sûr la sortie du
champ de la caméra.
Mais on peut aussi
imaginer d'autres phénomènes de
déplacement de matière au sein de la cellule
qui rendraient ces observations, purement morphologiques et
externes à partir d'une forme figée,
fausses. |
|||||||
|
||||||||
|
|
||||||||
|
Les bactéries communiquent grâce à des nanotubes, Anne Debroise, La Recherche, 452, mai 2011, 20 Intercellular nanotubes mediate bacterial communication, Dubey and Ben-Yehuda, Cell, 144, 590, 2011 |
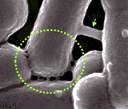 De grands (flèche verte, environ 130 nm de Ø) et petits (cercle en pointillés, environ 30 nm de Ø) nanotubes assurant une communication cytoplasmique entre des cellules de Bacillus subtilis (le diamètre du cercle est d'environ 0,8 µm) Image extraite de l'original accessible ici librement (sans autorisation mais avec une résolution très faible) |
On connaissait déjà les
"pili" ou "poils sexuels" qui étaient des ponts
cytoplasmiques établis entre bactéries
CONJUGUANTES. Ces dernières s'échangeaient
alors du matériel génétique dans le
sens donneuse -> receveuse. Ces nanotubes ont été
observés aussi bien entre les cellules d'une
même espèce bactérienne (B.
subtilis) qu'entre cellules d'espèce
différente (B. subtilis et Staphylococcus
aureus). Ce mode de communication doit encore être exploré (généralité, sens du transfert, mode d'apparition des ponts, mécanisme moléculaire du transport des substances, devenir des molécules transférées...). |
||||||
|
|
||||||||
|
une
référence récente pour comprendre
l'aspect moléculaire non traité ici bien
sûr: Bacterial Cell Division: The Mechanism and Its
Precison, Elizabeth Harry, Leigh Monahan and Lyndal
Thompson, 2006, A Survey of Cell Biology, Volume 253, pp
27-94, |
La multiplication des cellules bactériennes serait la croissance d'un individu pluricellulaire, sous le contrôle de la nutrition. Chaque cellule bactérienne de l'individu colonie-bactérienne serait très jeune (guère plus de 4 divisions d'ancienneté). Puisque la vie de l'individu dure au moins pendant 4 multiplications, le cycle cellulaire n'est pas adapté pour décrire la vie de l'individu. 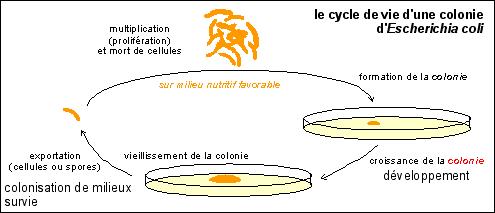 On peut considérer la colonie comme l'individu, la cellule bactérienne n'étant alors qu'une partie de l'organisme pluricellulaire... La reproduction est la phase qui fait passer d'une colonie à une autre colonie à partir d'une cellule ou d'une spore qui se détache de la colonie et va dans un autre milieu. |
|||||||
|
|
||||||||
|
3 - La
prolifération des cellules eucaryotes peut avoir des
finalités différentes
|
Exemple étudié en TP: l'hermelle ou Sabelle, documents à partir du site de bmedia |
|||||||
|
Une riche source de vidéos: http://www.bioclips.com/dvd/ |
3.1 Se multiplier pour se développer La prolifération des cellules embryonnaires chez un pluricellulaire ... est une suite de multiplications de développement orientées et interdépendantes |
|||||||
|
* Ia multiplication des
cellules embryonnaires chez l'animal
n'exige pas forcément de phase de
croissance nette mais met en jeu plusieurs types de
divisions
|
||||||||
|
Un site "officiel" pour les enseignants : http://www.snv.jussieu.fr/bmedia/sommaires/dvpt.htm Un des laboratoires
français qui travaille AUSSI sur les aspects
cytologiques et non pas uniquement moléculaires du
développement , et qui présente un site
très riche : la station
de Villefranche sur mer. Voir
par exemple: |
Un beau film (téléchargez le fichier lié, modifiez son extension en ".mov" seul puis visionnez-le : http://biodev.obs-vlfr.fr/ recherche/ lepage/images/ cleave.mov%20copy) qui va de la première division à la fin de la gastrulation chez le poisson-zèbre sur la page de l'équipe de Thierry Lepage à Villefranche su mer (http://biodev.obs-vlfr.fr/recherche/lepage/). 30s de film correspondant à 11h20s de développement soit une accélération d'environ 1321 fois. Sur une page du même labo, un film (http://biodev.obs-vlfr.fr/ recherche/ lepage/ images/Film%20oursin%20pwp1.mpg) sur le développement de l'oursin méditerranéen Paracentrotus lividus qui vaut largement les films scolaires commerciaux (dommage que l'on ait pas le timing avec le temps séparant les prises de vues; on peut se référer au site de bmedia: http://www.snv.jussieu.fr/ bmedia/ oursinMDC/index.html et particulièrement à la chronologie : http://www.snv.jussieu.fr/ bmedia/ oursinMDC/timingdev.html ). |
 Photos "repiqués" du site de la station de Villefranche-sur-mer avec une plus faible dimension - pas de copyright connu Oursin méditerranéen  |
||||||
|
Un
film
(http://www.bio.unc.edu/faculty/salmon/lab/mafia/wt.mov)
montrant les 4 premières
divisions du zygote chez le ver
nématode Cænorhabditis elegans
en microscopie à contraste de phase;
le film correspond à
1h52min45s de prises de vues pour un film de 37s soit une
accélération d'environ 183 fois. |
 © image réduite http://exploration.nasa.gov/ programs/station/ images/ICE-First1.jpg Ces vers mesurent environ1 mm lorsqu'ils ont atteint la taille adulte. |
|||||||
|
d'autres films : http://www.bio.unc.edu/faculty/ salmon/lab/mafia/mafiamovies.html Une belle mitose (un peu partielle) chez le rat kangourou: http://www.bio.unc.edu/ faculty/salmon/lab/mafia/phmit1.mov Une mitose avec marquage de l'actine par une protéine GFP de cellules épithéliales du foie de porc (il vaut mieux visionner les images une à une... le film est un peu rapide): http://www.bio.unc.edu/faculty/ salmon/lab/mafia/gfpactinsheldon.mov |
||||||||
|
|
||||||||
|
Voir fiche : "les chromosomes" |
La multiplication cellulaire eucaryote binaire typique est décrite principalement par les 2 phases du noyau: l'interphase (noyau au repos), puis la mitose (noyau en division depuis l'apparition des chromosomes jusqu'à leur disparition). Parmi les phénomènes cytoplasmiques seule la cytodiérèse (séparation du cytoplasme en deux masses cytoplasmiques) porte un nom. |
|||||||
|
Film http://www.bio.unc.edu/ faculty /salmon/lab/ mafia/ urchin.mov de la première division chez l'oursin Lytechinus variegatus :  http://www.bio.unc.edu/faculty/salmon/lab/mafia/img18.gif |
Ces vues au microscope à contraste
de phase sont particulièrement intéressantes:
on peut noter
|
|||||||
|
Pour un petit aperçu des mystères de l'embryologie... |
|
|||||||
|
Pour des divisions égales et synchrones on peut voir l'extrait de film sur les premières divisions chez Asterina gibbosa (étoile de mer): http://www.cerimes.education.fr/index.php? page=bi_admin&op1=plan,,,99,,,,,1,1884 Le film en totalité vaut le coup d'œil : http://www.cerimes.education.fr/index.php? page=affiche_video,http://sfrs-streamer.sfrs.fr/ ramgen/media-2/sfrs/real/realvideo/ 515.rm,,269,1,7,386 |
Observez les deux premières divisions de l'oursin Paracentrotus lividus en microscopie optique; il faut extraire les 9 premières secondes du film http://biodev.obs-vlfr.fr/recherche/ lepage/images/Film%20oursin %20pwp1.mpg |
La première division est identique
à celle observée dans le film
précédant : le plan du film contient bien
l'axe de séparation des deux cellules filles (ou le
fuseau de division). L'orientation est environ
15-20°N-105-110°S. |
Si la seconde cytodiérèse
semble intervenir rapidement, c'est probablement qu'il
manque des images... L'orientation du fuseau de division est
plongeante obliquement en arrière du plan du film et
l'on ne voit pas les phénomènes fusoriaux. Le
deuxième fuseau est donc bien perpendiculaire au
premier. Le réaccolement intervient ensuite pendant que se prépare une troisième phase de divisions. |
|||||
|
|
||||||||
|
Je recommande un n° spécial
de The International Journal of Developmental
Biology, Vol 50, Nos 2/3, 2006 a pour titre
Morphodynamics. |
Si l'on observe les divisions suivantes,
plusieurs phénomènes peuvent être
notés: Les divisions successives ne sont donc pas indépendantes, leurs axes sont orientés et elles sont synchronisées ou non, et peuvent être inégales. On est donc loin du modèle d'une
cellule (boule) qui donne deux cellules identiques (boules).
Les divisions embryonnaires marquent au contraire une
construction ordonnée: celle de l'embryon. |
|||||||
|
voir cours de 1ère S Sur le site du CERIMES on peut trouver un petit extrait de film présentant la germination de la graine de moutarde des champs et la croissance des racines (en vue externe): http://www.cerimes. education.fr/ index.php? page=bi_admin& op1=plan,,,99,,,,,1,3624 |
* La multiplication cellulaire au sein d'une jeune racine de Plante (multiplication de développement) met en jeu une croissance importante sans déplacement La grande différence du développement chez les plantes par rapport à celui des animaux réside dans l'absence de déplacement des cellules des plantes qui sont réunies par une paroi sécrétée en permanence et dont la composition change au cours de la vie de la cellule (phénomène qui accompagne la différenciation). La croissance d'un tissu végétal est réalisée par un accroissement cellulaire (souvent un allongement mais pas toujours; ce qui permet une grande diversité des types de tissus) suivi de la formation d'une cloison (paroi) entre les deux nouvelles cellules. Les phénomènes nucléaires sont ceux d'une mitose avec des caractéristiques particulières, notamment l'absence de centrioles aux deux extrémités du fuseau de division. |
|||||||
|
Suivre le développement complet d'un champignon : Stropharia semiglobata sur http://www.cerimes.education.fr /index.php?page=fiches,view, 569,10,7,385,,croissance,, |
* Le développement d'un mycélium de champignon par allongement d'une cellule puis (le plus souvent) cloisonnement La croissance d'un hyphe (filament de champignon formé d'une file de cellules) se fait de façon similaire aux cellules végétales. Mais il n'y a jamais de cloisonnement longitudinal et l'hyphe mycélien est toujours constitué d'une seule file de cellules. Les formes (et notamment l'appareil reproducteur ou carpophore) des "organes" du champignon sont toutes réalisées à partir de l'agglomération de ces hyphes. Certains champignons (dits inférieurs) ne cloisonnent pas leur hyphe qui est donc composé d'un immense cytoplasme pourvu de nombreux noyaux. La paroi des champignons, parfois chitineuse (un "sucre", ou plutôt un composé du groupe des glucosamines...que l'on retrouve dans la cuticule des insectes), reste souple. Les champignons ne peuvent se développer sans eau. Les phénomènes nucléaires sont assez variés et les mitoses parfois atypiques. |
|||||||
|
|
||||||||
|
3.2 - Se multiplier pour accroître la population ou assurer sa survie La multiplication des cellules des unicellulaires
eucaryotes (royaume des Protistes) se fait selon des
modalités variées ; |
||||||||
|
|
+ Sporulation chez
un champignon qui ne présente qu'une forme
unicellulaire : la levure de
bière
(multiplication de
croissance)
|
Le mot "levure" désigne tout champignon unicellulaire ayant une multiplication par bourgeonnement ou scission binaire. |
||||||
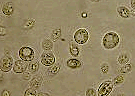 Saccharomyces cerevisiae MO X 600 - image partielle - www.inrp.fr/bdd_image/ gloss_levure.jpg |
La division se fait sans disparition de l'enveloppe nucléaire (on parle de mitose atypique...). La formation de la spore asexuée se fait par étranglement (un anneau de protéines (actine) se met en place: sa constriction (rétrécissement) accompagne la séparation de la spore de la cellule mère par une cloison (paroi)). Dans le film ci-dessous la division est
pratiquement égale, mais ce n'est pas le cas le plus
habituel, sauf pour des cellules jeunes de petite
taille. |
|||||||
|
film http://www.bio.unc.edu/faculty/ salmon/lab/mafia/contr.mov présentant la phase terminale de la division chez Sacharomyces cerevisiae (double vue en fluorescence avec marquage des protéines de type actine (anneau de division) - à gauche -, et en contraste de phase - à droite -): |
||||||||
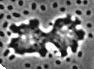
Une mixamibe (Dictyostelium
discoideum) en mitose; chaque
cellule fait près de 10µm diamètre si
l'on compte les prolongements; |
Lorsque la nourriture vient à manquer les myxamibes s'agglomèrent sous l'action d'un facteur stimulant qu'elles sécrètent (l'AMPcyclique) rn un pseudoplasmode (un plasmode est une cellule plurinucléée, souvent issue de la fusion de plusieurs cellules). Après un certain temps, les cellules du pseudoplasmode se différentient et forment un pied surmonté d'une tige puis d'une fructification ou sporocarpe. (voir le superbe montage photos (http://cosmos.bot.kyoto-u.ac.jp/ csm/photo_html/Dd_cul_1.html) et les films (http://cosmos.bot.kyoto-u.ac.jp/ csm/movies.html). |
Les amibes acrasiales sont encore appelées myxamibes ou moisissures visqueuses cellulaires; on les regroupe dans un embranchement des mycètes : les Acrasiomycota. |
||||||
|
film
http://www.bio.unc.edu/faculty/
salmon/lab/mafia/bigrc.mov film http://cosmos.bot.kyoto-u.ac.jp/ csm/movies2/M2_division_3.mov présentant le déplacement et la nutrition de Dictostelium discoideum (elle phagocyte des bactéries) ; la barre fait 10 µm. Aucun phénomène nucléaire n'est visible. La totalité du film correspond à 13 min et 40 s pour 10 s de film. La mitose dure environ 4 min. |
||||||||
 Euglena sp. (chaque individu mesure plus de 100 µm lorsqu'il s'allonge au maximum) photo extraite de http://bio.rutgers.edu/euglena/ samples.htg/colorLM.jpg |
|
|
||||||
|
Le site internet (en anglais) est entièrement dédié à ces organismes ; http://euglena.msu.edu/ |
De belles images de Cyclops (crustacé) parasité par un euglène du genre Colacium : http://aelmh.free.fr/ Colacium_vesiculosum.htm |
|||||||
|
Série de photos http:// bio.rutgers.edu/ euglena/eug_ mitosis_web/mito_ sequence_lab.jpg montrant la mitose chez Peranema trichophorum en microscopie à contraste de phase On observe surtout l'étirement du nucléole et secondairement celle du noyau. |
||||||||
|
|
Les diatomées sont aussi appellées Bacillariophyceae (ce qui signifie algues en forme de "bacille", c'est-à-dire de "bâtonnet", comme les bactéries) |
|||||||
 Pseudo-nitzschia http://planktonnet. sb-roscoff.fr //rawdata/ viewable/ mmoniz_pseudo-nitzshia_australis_ n_xxw_ 20061114111555_w.jpg
Pour voir la frustule au MEB voir par exemple http://www.whoi.edu /redtide/ species/ pseudo-nitzschia_multi.jpg |
Les diatomées du genre Pseudo-nitzschia sont des Protistes chlorophylliens dont le test rigide formé de deux valves emboîtées (appelé frustule) est siliceux. Elles sont capables de mouvements de glissement rapides de leurs frustules le long de sillons (raphé). Elles sont autotrophes mais de nombreuses espèces sont capables de devenir allotrophes dans des milieux riches en matière organique et peu éclairés. Les cellules sont très allongées et peuvent atteindre 140 µm de long pour 1,5 à 3,4 µm de large. Les cellules réunies au sein d'une même gangue mucilagineuse (glycoprotéique) forment des chaînes que l'on peut qualifier de colonies. La multiplication de croissance de la
colonie est une mitose banale pour ce qui est du noyau. La
cytodiérèse s'accompagne de la synthèse
de deux nouvelles valves qui s'emboîtent dans chacune
des valves de la cellule mère. On a donc une
diminution de taille à chaque division. Une photo au
MO est disponible sur le site de l'Ifremer
(http://www.ifremer.fr/
envlit/ photos/Archive/
20041101/photo06_2.htm).
On distingue très bien les deux plastes,
déjà dupliqués, et la formation des
nouvelles valves au centre de la cellule. En dessous d'une taille critique a lieu la reproduction sexuée ou auxosporulation qui nécessite l'accouplement de deux cellules et deux méioses. La colonie peut aussi former des spores de survie entourées d'une épaisse paroi silicifiée qui mènent une vie ralentie. On parle beaucoup de Pseudo-nitzschia en Bretagne car ce genre possède plusieurs représentants toxiques (la toxine incriminée est l'acide domoïque) mais la détermination des espèces toxiques est impossible au seul microscope optique (voir page de l'Ifremer). On a donc des données de comptage qui contiennent à la fois des espèces toxiques et non toxiques. |
Sur les mouvements de P. on peut consulter (en anglais): http://www.mbari.org/ staff/ conn/botany/diatoms/ jennifer/mot.htm D'une façon générale les pages (en anglais) http://www.mbari.org/staff/ conn/ botany/diatoms/ jennifer/ indexa.htm sont assez bien renseignées (mais l'article "Diatomées" de l'Encyclopedia Universalis l'est aussi). On peut aussi visiter http://www.indiana.edu/ ~diatom/ diatom.html. http://www.ifremer.fr/ envlit/ documentation/ dossiers/toxines10ans / rephy-c4.htm Pour une petite vidéo présentant des généralités sur quelques diatomées voir http://www.cerimes.education.fr /index.php? page=bi_admin& op1=plan,,,99,,,,,1,3850 |
||||||
|
|
||||||||
|
4 - Après s'être divisées, les cellules des pluricellulaires se différencient sauf les cellules souches qui restent indifférenciées |
La différenciation
est l'état d'une cellule eucaryote qui,
après une division, change profondément sa
structure et perd sa capacité à se diviser.
|
|||||||
|
voir cours de 1ère S pour la différenciation des cellules des plantes |
On ne peut pas traiter de la seule multiplication des cellules eucaryotes sans aborder leur devenir. Chez les unicellulaires, on peut considérer que la formation d'un kyste ou d'une spore de protection est une forme de différenciation qui permet la survie dans des conditions défavorables et donc la reproduction. Chez les pluricellulaires la différenciation est la règle. En effet les cellules, une fois multipliées, changent profondément de forme, de structures et surtout perdent la capacité de se diviser. Lors de cette différenciation l'information génétique est profondément modifiée: de l'ADN est souvent perdu mais parfois une grande quantité d'ADN est multipliée de nombreuses fois. On peut considérer que la mort est la suite normale de la différenciation. Les cellules du xylème, par exemple, dans les tissus conducteurs des plantes, meurent, leur paroi se lignifie (s'imprègne de polyphénols qui la rigidifie) et leur cytoplasme est digéré. Certaines cellules ont un devenir assez original: elles fusionnent. On connaît des fusions cellulaires dans les cellules musculaires des muscles striés par exemple : les myocytes se forment à partir de la fusion de plusieurs dizaines de myoblastes. C'est pour cela que ce sont des cellules géantes pourvues de dizaines de noyaux. |
|||||||
|
www.genethique.org, le site de référence sur l'actualité des recherches sur les cellules souches (et de bien d'autres questions touchant à la génétique et à ses applications) avec un regard humaniste chrétien. |
|
|||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}