B1 - La relation par héritage ou la division
cellulaire
Cette page a
été modifiée et scindée en plusieurs
pages : voir plan
du cours
retour plan du cours, accueil
Plan de cette page
B1. 1 - Chez
les unicellulaires la division est une reproduction
B1.1.1 - cycle
cellulaire chez Escherichia coli
B1.1.2 - cycle
cellulaire chez les eucaryotes
B1.2 - Chez
les pluricellulaires la division conduit à une
différenciation
B1.2.1 Chez
les plantes et les champignons la division n'est jamais suivi de
déplacement
B1.2.2 Chez les
animaux certaines cellules se déplacent activement
Annexe: les jonctions
cellulaires
La théorie cellulaire (voir page sur
la cellule), affirme que "toute cellule vient d'une autre
cellule": c'est une relation par
héritage.
Selon que l'on a affaire à un unicellulaire ou à un
pluricellulaire la signification de la division n'est pas la
même, nous traiterons donc deux parties.
B1.1 -
Chez les unicellulaires la division est une reproduction
La division d'un unicellulaire est une reproduction qui
donne deux individus pour partie nouveaux et pour partie anciens (du
point de vue de la matière) et hérités (qui
conservent les dynamiques de l'espèce à laquelle
appartient l'individu mère: c'est cette relation
d'éhritage que l'on qualifie de copir conforme dans la
théorie de l'information génétique; mais il y a
bien plus dans l'héritage que de l'information
génétique).
La reproduction des unicellulaires conduit à des relations
entre organismes issus de la division. Considérer les
organismes dans leur milieu relève de
l'écologie. Le terme vient du grec
oïkos (l'habitat) et logos (parler). Les
procaryotes sont considérés comme des cellules
isolées mais leur écologie est bien plus riche. Les
bactéries forment des colonies. De nombreuses espèces
forment des biofilms à la surface de solides (galets au fond
d'une rivière, feuille d'une plante, muqueuse buccale d'un
mammifère...tuyauterie domestique...).
D'autres espèces vivent en étroite relation avec
d'autres organismes: c'est la symbiose au sens large
(du grec syn = avec et bio = vie). On distingue avec
parfois un peu d'anthropomorphisme et de façon peu
tranchée: le commensalisme (partage d'une source de
nourriture), la symbiose au sens strict (association à
bénéfices réciproques) et le parasitisme (un
organisme parasite vit aux dépens d'un organisme
hôte).
Sources :
Le cycle cellulaire chez les animaux et les végétaux,
Jean Clos, Marc Coumans et Yves Muller, Biologie-Géologie
(revue de l'APBG), n°3-2002, p497-564
Biologie végétale, tome 2 : Organisation
végétative, D. Robert et A.M. Catesson, Doin, 1990
Biologie moléculaire de la cellule, Alberts et al., 1994,
Médecine-Sciences Flammarion
Biologie du Développement, S.F. Gilbert, 1996, De Boeck
Université
Microbiologie, Prescott, Harley et Klein, 1995, De Boeck
Université
B1.1.1 -
cycle cellulaire chez Escherichia coli
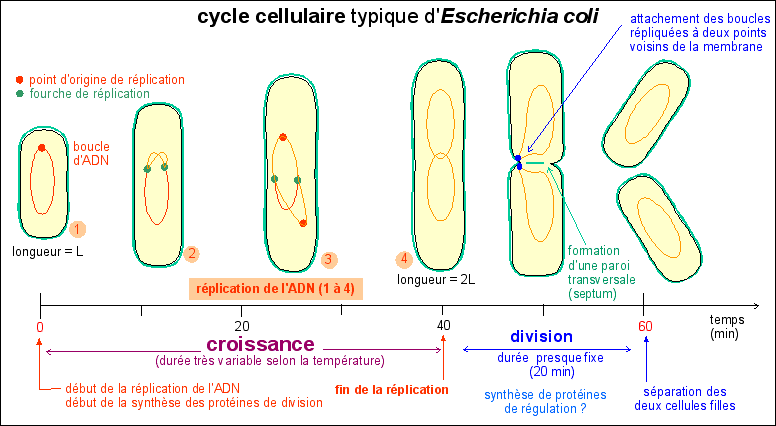
Le cycle cellulaire comprend les
événements situés entre deux divisions.
Il comprend une phase de croissance cellulaire de durée
variable, pendant laquelle l'ADN est répliqué, et une
phase de division de durée fixe (20 min) chez la
bactérie Escherichia coli, qui répartit l'ADN dans les
deux cellules filles qui se séparent.
Il commence donc, pour une cellule, lors de sa séparation
d'avec sa cellule sœur; et finit lors de sa séparation en
deux cellules filles. Le cycle cellulaire commence par un allongement
de la cellule, sans augmentation de diamètre, qui atteint
ainsi 2 fois sa longueur initiale : c'est la croissance cellulaire.
Comme il n'y a pas d'augmentation de diamètre
on peut penser que la masse initiale double pendant cette croissance
étant donné que la bactérie peut être
assimilée à un cylindre de diamètre constant.
Pendant cette croissance l'ADN est dupliqué et de
nombreuses protéines dites de division sont
synthétisées. Si l'on bloque la réplication de
l'ADN on empêche la division et la cellule s'allonge
démesurément en formant un long filament. La division
cellulaire commence presque toujours, quelque soient les conditions
de culture de la bactérie et donc quelque soit la durée
du cycle cellulaire, 20 minutes avant la fin de la
réplication.
Remarque: dans certains cas, la croissance est
très rapide et une nouvelle phase de réplication de
l'ADN commence immédiatement alors que la division de fait que
commencer. Ce qui conduit à un phénomène continu
de réplication (d'après Microbiologie, fig 11.18 et
11.19).
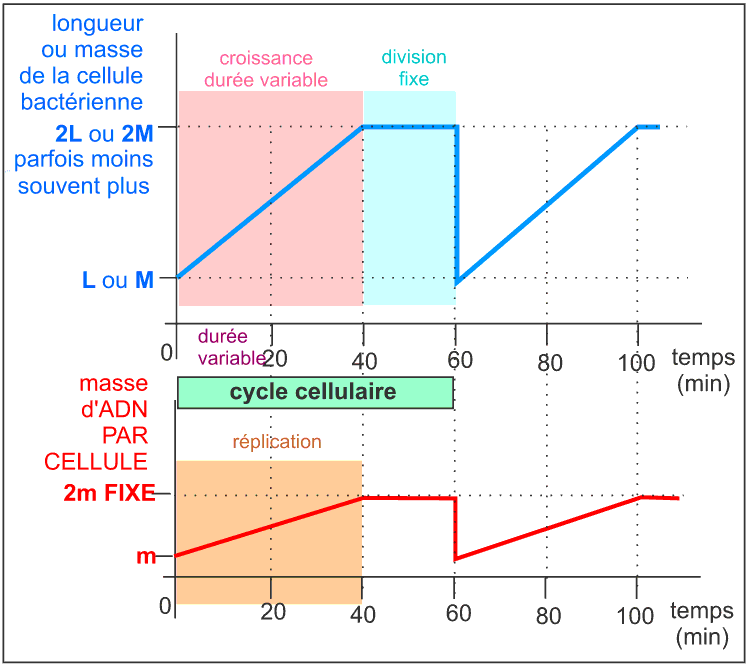
Le cycle cellulaire - d'une durée de 60 min - d'une souche
d'Escherichia coli à croissance lente
Remarque:
Le cycle cellulaire ne se superpose pas au cycle de vie
bactérien car ne nombreuses bactéries peuvent sporuler,
c'est-à-dire former une spore résistante, qui
leur permet notamment de résister dans des milieux
extrêmes et d'être transportées par l'air sur de
très longues distances.
De plus il existe des conjugaisons bactériennes qui
sont des échanges d'ADN entre bactéries dites
recombinantes.
La réplication de l'ADN est de loin le mécanisme le
mieux connu de tout le cycle cellulaire des Procaryotes. Elle a
particulièrement été étudiée chez
Escherichia coli. La division est fort mal connue.
L'expérience historique de Meselsohn et Stahl a permis de
préciser le type de mécanisme de réplication de
l'ADN.
|
Expérience de Meselson et
Stahl (1958) - Bordas p 101;
Nathan p 112
|
|
Escherichia coli est cultivée pendant de
nombreuses générations (= durée d'un
cycle cellulaire puisqu'une génération
sépare une cellule mère de sa cellule fille)
sur un milieu de culture contenant des
désoxyribonucléotides marqués à
l'15N (témoin 1). Cette souche de
départ est transférée pendant un
génération sur un milieu contenant uniquement
des désoxyribonucléotides marqués
à l'14N (expérience 1). Enfin on
transfère des bactéries issues de cette
culture à nouveau sur un milieu contenant uniquement
des désoxyribonucléotides marqués
à l'15N (expérience 2).
La mesure de la quantité de
désoxyribonucléotides incorporés dans
l'ADN lors de la phase de réplication (croissance
cellulaire) se fait à l'aide de
prélèvement de bactéries dont l'ADN est
extrait et centrifugé sur gradient de densité;
l'ADN se plaçant plus ou moins bas dans le tube selon
sa densité. La bande d'ADN, fluorescente sous U.V.
est révélée par éclairement U.V.
du tube. Une densité de 1,8 signifiant un ADN ne
contenant quasiment que de l'15N et une
densité de 1,65 signifiant un ADN ne contenant
quasiment que de l'14N.
Le tube 2 correspond à un témoin d'une souche
cultivée pendant de nombreuses
génération sur sur milieu contenant des
désoxyribonucléotides marqués à
l'14N. Le tube 5 correspond à une
génération supplémentaire sur milieu
contenant des désoxyribonucléotides
marqués à l'14N (expérience
3).
|
|
densités
(selon les
auteurs!!!!)
|
tube 1
(témoin 1)
|
tube 2
(témoin 2)
|
tube 3
(expérience 1)
bactéries de la souche
témoins 1 cultivées 1
génération sur
14N
|
tube 4
(expérience 2)
bactéries de la souche
témoin 1 cultivées 2
générations sur
14N
|
tube 5
(expérience 3)
bactéries de la souche
témoin 1 cultivées 3
génération sur
14N
|
|
1,65 - 1,710
|
-
|
*********
*********
|
-
|
********
|
********
|
|
1,72 -1,717
|
-
|
-
|
*********
*********
|
********
|
****
|
|
1,80 - 1,724
|
*********
*********
|
-
|
-
|
-
|
-
|
Interprétation:
Les témoins correspondent à des
bactéries dont la quasi-totalité de l'ADN est
composé de désoxyribonucléotides
possédant de l'14N (témoin 2) qualifié
d'ADN "léger" ou de l'15N
(témoin 1) qualifié d'ADN "lourd".
Dans l'expérience 1 (tube 3) la position de l'ADN
intermédiaire en densité entre l'ADN
léger et de l'ADN lourd permet de dire que le
mécanisme de réplication de l'ADN, qui s'est
normalement déroulé une seule fois depuis le
transfert sur milieu léger, est
semi-conservatif. En effet, les molécules
d'ADN obtenues lors de la réplication contiennent
TOUTES pour moitié de l'14N et de
l'15N. Ce qui peut s'expliquer aisément si
l'on imagine un système de réplication qui
conserve un des brins anciens dans chaque nouvelle
molécule et synthétise un brin nouveau
complémentaire. Les deux molécules d'ADN
issues d'une telle réplication sont donc identiques
et pour un brin (nouveau) composées d'14N
et pour l'autre brin (ancien) composées
d'15N. On peut parler de molécules
hybrides 14N-15N.
Lors d'une seconde réplication avec des
désoxyribonucléotides contenant de
l'14N (milieu "léger"), les
molécules hybrides sont aussi
répliquées selon un mécanisme
semi-conservatif qui donnera une molécule d'ADN
léger et une molécule d'ADN hybride à
partir de chaque molécule ancienne (hybride).
Une troisième réplication sur "milieu
léger" donnera aussi ces deux types de
molécules mais dans un rapport différent. Il y
aura une molécule hybride pour 3 molécules
d'ADN léger.
|
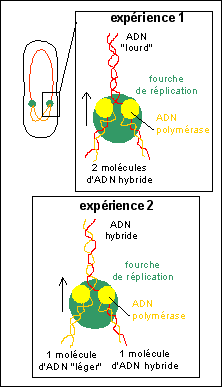
Une illustration de
l'interprétation du mécanisme de
réplication semi-conservative de l'ADN dans
l'expérience de Meselson et
Stahl.
|
La
réplication de l'ADN est une
synthèse de deux molécules d'ADN
identiques à partir d'une molécule d'ADN. Chaque
molécule d'ADN fille est composée d'un brin ancien et
d'un brin nouveau complémentaire du brin ancien; on dit que la
réplication est semi-conservative. La
réplication de l'ADN est catalysée par un gros complexe
enzymatique contenant plus de 20 enzymes mais surtout l'ADN
polymérase. La réplication commence en un point origine
et se poursuit en général dans deux sens opposés
au niveau de fourches de réplication. Chez les eucaryotes, du
fait de la longueur du génome, elle commence en de multiples
points origine simultanément. (Il
faudrait plus de 500 h à raison de 50 nucléotides
à la seconde pour répliquer un seul chromosome humain
à partir d'un seul point origine).
L'ADN polymérase nécessite une molécule d'ADN
à répliquer, des désoxyribonucléotides,
de l'énergie (ATP) et un petit fragment d'ARN
complémentaire d'une petite séquence de l'ADN à
répliquer (amorce). Il existe plusieurs ADN polymérases
chez les bactéries (au moins 3) et plus encore chez les
eucaryotes. La réplication se déroule avec une
fidélité excellente: chez E. coli on estime le nombre
d'erreurs de copie à 1 pour 109 à 1010
nucléotides, ce qui fait, pour un génome
évalué à 4,7.106 paires de bases, une erreur
pour 1.000 à 10.000 réplications. Il existe des
systèmes de réparation de l'ADN qui utilisent aussi une
ADN polymérase. La vitesse de polymérisation (ajout des
désoxyribonucléotides pour former le nouveau brin) est
de l'ordre de 20 à 1.000 nucléotides par seconde chez
E. coli.
Le problème du déterminisme du cycle cellulaire est
aussi celui du lien entre la croissance cellulaire et la
réplication de l'ADN.... il est loin d'être
résolu. En tout il ne sera pas traité dans ces
pages.
B1.1.2
- cycle cellulaire chez les eucaryotes
Le cycle cellulaire des eucaryotes à surtout
été étudié chez la levure de
bière. Les données obtenues chez les cellules en
culture et les cellules embryonnaires sont les plus nombreuses pour
les animaux. Selon l'organisme, l'âge de la cellule, et le type
cellulaire, on a des différences fondamentales. Seuls quelques
points communs seront présentés ici. En tout cas il est
clair que pour un pluricellulaire la notion de cycle de
développement, qui se réfère à
l'individu, est plus adéquat que la notion de cycle
cellulaire, qui ne rend compte que de la vie d'une seule cellule.
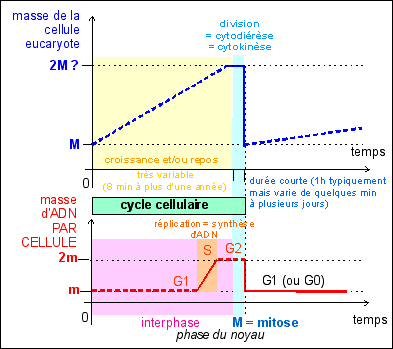
|
Le cycle cellulaire des
cellules eucaryotes est beaucoup plus variable que celui des
cellules procaryotes. Deux phases sont toujours
observées:
- une phase de croissance,
la plus longue, très généralement
continue, qui correspond à l'interphase
nucléaire (et pendant laquelle l'ADN est
répliqué);
- et une phase de division
(cytodiérèse) concomitante avec la
mitose, division du point de vue nucléaire,
très généralement brusque et plus
courte que la phase précédente
|
a - les événements nucléaires
En interphase le noyau est au repos du point de vue de la division
mais pas de la synthèse car c'est en phase S de l'interphase
qu'a lieu la réplication de l'ADN qui précède
toute division (sauf dans le cas de la
deuxième division de la méiose, voir cours de
terminale).
L'expérience de Taylor met en évidence une
caractéristique essentielle de cette phase de
réplication: c'est une réplication semi-conservative.
Elle fait appel aux mêmes mécanismes que la
réplication chez les procaryotes. Les molécules d'ADN
polymérases et les nombreuses enzymes associées sont
bien moins connues que chez les procaryotes.
|
Expérience de Taylor
(1957) - Bordas p
100
|
|
Les cellules eucaryotes cultivées in vitro sont
des fragments racinaires de Bellevalia romana, une
plante de la famille du Lys. La réplication de l'ADN
est suivie à l'aide de thymidine tritiée (la
thymidine est le nucléoside (thymine + sucre))
marquée par le tritium (3H ou 3T) isotope lourd et
radioactif de l'hydrogène. L'incorporation du
désoxyribonucléotide radioactif est suivi par
des autoradiographies qui sont des expositions de
préparations cellulaires à une plaque ou un
film photographique. Les composés argentés de
l'émulsion photographique précipitent
lorsqu'ils rencontrent un électron émis par
radioactivité ß- par les atomes de 3H
incorporés dans le thymidine radioactive. Les grains
d'argent forment des points noirs que le
développement révèle. L'emplacement des
points noirs indique donc de façon approximative la
position des molécules de thymidine tritiée de
la préparation cellulaire. Les temps d'exposition des
émulsions photographiques sont bien sûr
beaucoup plus longs que pour une impression par les
photons.
|
|
Les racines de Bellevalia romana, sont
cultivées pendant la durée d'un cycle
cellulaire sur un milieu "chaud", c'est-à-dire
contenant de la thymidine tritiée. Les cellules en
métaphase de mitose sont repérées et
certaines sont autoradiographiées. L'aspect d'un
chromosome est représenté ci-dessous à
gauche. Certaines de ces cellules sont replacées
immédiatement sur un milieu "froid",
c'est-à-dire dépourvu de thymidine radioactive
, pendant la durée d'un deuxième cycle
cellulaire. L'aspect d'un chromosome d'une autoradiographie
d'une cellule en prophase de mitose est
présentée ci-dessous à droite.
|
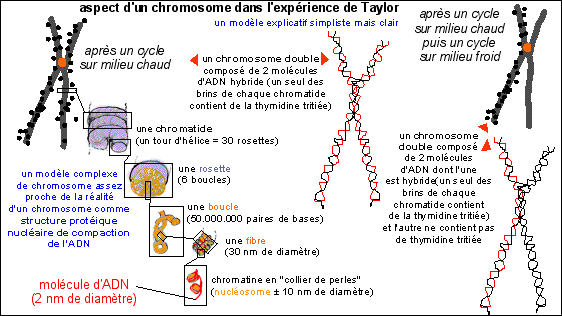
Illustration de l'interprétation de
l'expérience de Taylor
(en orange: le centromère (qui n'est
pas visible); les points noirs correspondent aux grains
d'argent de l'autoradiographie)
(schémas d'après Principe
de Biochimie, Lehninger et al, 1994,
Flammarion-Médecine-Sciences)
|
Explication de
l'expérience:
Tout en n'oubliant pas que le chromosome est une structure
complexe de compaction de l'ADN nucléaire (comme le
représente le schéma ci-contre -
l'ADN d'une cellule humaine est
estimé à environ 2 m soit quelques
centimètres par chromosome pour un diamètre de
2 nm soit un rapport longueur sur diamètre de
quelques dizaines de millions !!!!) on
s'intéresse à la composition de chaque
chromatide.
Les chromatides radioactives sont composées d'une
molécule d'ADN hybride. C'est-à-dire que la
molécule d'ADN hybride à incorporé dans
UN de ses brins, le brin nouvellement formé, de la
thymidine tritiée. Cette incorporation n'a lieu que
pendant la phase S du cycle cellulaire (synthèse de
l'ADN) et ne touche donc que les cellules qui étaient
dans cette phase de l'interphase peu après le
début de la mise en culture. Lorsque les cellules
sont retirées du milieu chaud et observées,
seules celles qui se trouvent maintenant en métaphase
peuvent présenter de tels chromosomes.
Le fait que les deux chromatides, issues de la
réplication de l'ADN en phase S de l'interphase, sont
TOUTES LES DEUX hybrides (c'est-à-dire
composées d'un brin ancien, non radioactif et d'un
brin nouveau ayant incorporé de la thymidine
tritiée radioactive) peut être expliqué
par un mécanisme semi-conservatif de la
réplication de l'ADN, comme chez les Procaryotes.
|
Remarque:
les molécules d'ADN dans le noyau en interphase ne sont pas
libres et flottant dans le nucléoplasme mais sont à
l'état de chromatine. La
chromatine a un aspect en "collier de perles" (voir
l'illustration ci-dessus), est colorable en
ME (par le
tétroxyde d'Osmium) et comporte
un filament d'ADN et des protéines (des
histones) regroupées en petits cylindres (les
nucléosomes). Cette structure ne semble
pas empêcher la réplication et les nucléosomes
semblent être aussi présents dans le chromosome qui
résulterait alors de la compaction de la
chromatine (voir de nouveau
l'illustration ci-dessus qui n'est cependant qu'un modèle -
voir cours de
seconde pour quelques remarques à
ce sujet).
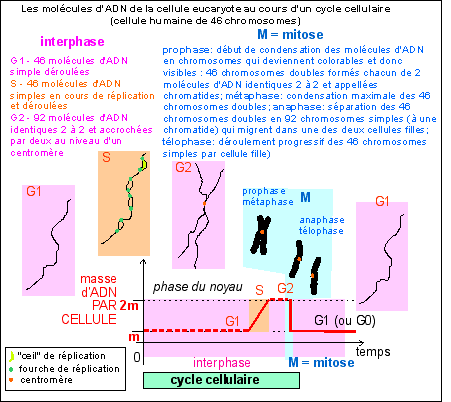
|
Place de la réplication dans
le cycle cellule d'une cellule eucaryote et aspect
très schématique des chromosomes au cours des
différentes phases nucléaires.
Le "piège" pour un élève inattentif
vient de ce que certains auteurs qualifient, improprement,
de chromosome, tout filament d'ADN de la cellule. Ce
mot doit être réservé aux structures
colorables et donc composées d'ADN et de
PROTÉINES de la cellule en phase M du noyau
(voir ancien
cours de spécialité de
terminale).
|
Les étapes de la mitose sont
classiquement regroupées en 4 phases qui présentent
bien sûr des limites parfois floues ou chevauchantes:
* la prophase où la chromatine se condense et forme les
chromosomes qui deviennent COLORABLES et VISIBLES en
microscopie optique; ce sont des chromosomes doubles (à deux
chromatides issues de la réplication en phase S de
l'interphase); l'enveloppe nucléaire se
désagrège par fragmentation des citernes du
réticulum qui entourent le nucléoplasme ;
* la métaphase où les chromosomes MIGRENT car
ils sont TIRÉS par les microtubules kinétochoriens et
se disposent de part et d'autre du plan équatorial de la
cellule en division; ce plan décrit de façon
précise l'orientation des deux cellules filles
(il est
matérialisé par un anneau ou un disque de filaments
d'actine, voir partie b suivante); si la
cellule est étirée, ce plan peut diviser la cellule
longitudinalement ou transversalement ce qui donnera une division en
épaisseur ou en longueur, déterminante pour la
croissance des cellules végétales à paroi par
exemple; entre les deux pôles de division, situés
symétriquement par rapport au plan équatorial, se
mettent en place des microtubules qualifiés de fusoriaux car
ils dessinent un fuseau de division;
* l'anaphase où les chromosomes doubles SE SCINDENT au
niveau des centromères, chaque chromosome fils (composé
d'une chromatide) est TIRÉ vers l'un des pôles de la
cellule par ses microtubules kinétochoriens qui glissent le
long des microtubules du fuseau de division;
* la télophase où les chromosomes se
déroulent et les filaments d'ADN redeviennent NON COLORABLES
sous forme de chromatine; l'enveloppe nucléaire se met
à nouveau en place autour du nucléoplasme.
Remarques:
* toutes les "figures de mitose" ou "cellules en
mitose" magnifiques de vos livres scolaires photographiées
au microscope optique sont des COLORATIONS SPÉCIFIQUES
des seuls CHROMOSOMES et, si la cellule vous semble VIDE, c'est
qu'elle est TRANSLUCIDE et NON COLORÉE artificiellement comme
le sont les chromosomes (rappelez-vous l'étymologie de
chromosome: chromo = couleur et soma = corps: des corps
colorables). Vous pouvez comparer ces photos avec les coupes
ultrafines fixées et colorées (au tétroxyde
d'osmium, colorant opaque aux électrons) observées au
microscope électronique: vous verrez alors le
cytoplasme nettement plus encombré; tout en sachant que,
là encore, on ne colore, avec le tétroxyde d'osmium,
que certaines structures et non pas toutes... (par exemple: Bordas p
98, 103; Nathan p 105).
* si vous cherchez à
approfondir les mécanismes de la division nucléaire
vous serez rapidement plongés dans un inextricable
réseau de molécules intervenant à telle ou telle
étape du cycle cellulaire, chacune étant censée
être indispensable à tel ou tel mécanisme
précis: assemblage des microtubules, appariement des
chromosomes, déplacement, clivage... Et pourtant cette vision
est loin d'être claire et exempte d'a priori contestable. C'est
au moyen de "mutants" sélectionnés pour telle ou telle
déficience cytologique que l'on s'est efforcé de
construire ce réseau explicatif. Le leitmotiv étant le
pouvoir explicatif de la protéine active
(voir partie
précédente).
Dans le cadre de ce cours je crois beaucoup plus fécond de
remplacer à chaque fois que possible la fonction locale par
l'insertion dans une fonction globale.
Voici un exemple: dans le texte suivant issu de Cloning Mammals:
What Does It Mean ? (An
opinion from Scott Gilbert) Breaking News: Primate cloning might not
be possible with existing methods
(http://8e.devbio.com/article.php?id=205);
il suffit de remplacer "étaient du" par
"conduisaient à" pour replacer correctement les
relations de causalité: d'abord les dynamiques, ensuite les
molécules ...«
When nuclear transplantations were done with the rhesus oocytes, the
chromosomes did not assort correctly to the mitotic spindle fibers,
and the microtubules, themselves, were not arrayed properly. The
cells became aneuploid. Simerly and colleagues further showed that
these disruptions were due to the absence of two proteins, NuMA and
HSET on the rhesus mitotic spindles. NuMA is a protein responsible
for spindle pole assembly, and HSET part of the kinesin motor system
along the microtubules. In the rhesus oocytes, these proteins
aggregate at the centrosomes of the meiotic
spindles»...Lors des
transplantations nucléaires avec les oocytes rhésus,
les chromosomes ne s'apparient pas correctement le long des fibres
fusoriales, de même que les microtubules eux-mêmes ne se
disposent pas correctement. Les cellules deviennent aneuploïdes.
Simerly et ses collaborateurs ont ensuite montré que ces
désordres étaient dus à l'absence de deux
protéines, NuMA et HSET au niveau des fibres fusoriales.
NuMA est une protéine responsable de l'assemblage du fuseau au
niveau des pôles et HSET fait partie du moteur à
kinésine situé le long des microtubules. Dans les
oocytes rhésus ces protéines restent
agglomérées au niveau des centrosomes des fuseaux
mitotiques.
Pourquoi ne pas tout simplement dire que l'absence de ces deux
protéines est la marque des dysfonctionnements
observés. Les protéines ne sont pas une cause de
l'appariement mais bien la trace d'une dynamique. Ce qui est
caché est la relation entre la dynamique (qui est un fait) et
l'absence de telle ou telle protéine (qui n'est pas un fait
d'observation mais la conséquence d'une
sélection).
b. Les événements cellulaires
Plus difficiles à observer que les chromosomes, les
composants cytoplasmiques sont de mieux en mieux connus, du fait des
progrès des techniques d'observation et de marquage
spécifique. Les films pris au microscope à contraste
interférentiel, ont laissé désormais la place
aux films présentant un marquage immunologique par
plusieurs substances fluorescentes de couleurs différentes
qui permettent in suivi IN VIVO des protéines comme la
tubuline (en VERT le plus souvent), composant
des microtubules, de l'actine (en ROUGE le plus
souvent), composant de certains filaments fins de la cellule,
ou encore des histones (en BLEU le plus
souvent), composant de la chromatine (Nathan p
115; voir par exemple le CDRom qui accompagne la nouvelle
édition (2004) de Biologie Moléculaire de la Cellule,
Alberts et al., Médecine-Sciences-Flammarion... de très
nombreux films au format ".mov" sont disponibles... et copiables pour
être visionnées avec Quick-Time en
vidéoprojection.... une merveille de souplesse
pédagogique). Cette vision est cependant plus chimique
que structurale et il faut se garder d'abandonner les anciennes
méthodes. Les nouvelles méthodes doivent
compléter les anciennes.
Une coloration spécifique (rouge) existe pour les microtubules
en microscopie optique (voir par exemple Bordas p 102-103).
Les microtubules (cylindres creux et
rectilignes de 25 nm de diamètre) et les filaments d'actine
(microfilaments flexibles d'environ 5 à 9 nm de
diamètre) constituent un cytosquelette
dynamique. Il existe aussi des filaments
intermédiaires (d'environ 10 nm de diamètre), par
exemple ceux qui constituent un réseau dense situé
contre la membrane interne de l'enveloppe nucléaire et que
l'on qualifie de lamina nucléaire.
Les mitochondries, les chloroplastes,
l'appareil de Golgi, le réticulum endoplasmique sont des
structures incapables de se régénérer à
partir de leurs composants, ils doivent être transmis par
hérédité cytoplasmique directe. Les plus gros se
fragmentent (RE, Golgi, chondriome
(voir remarque
ci-dessous)), le plus petits se
répartissent après une phase de
synthèse-multiplication (mitochondries ? chloroplastes ?). Une
phase de synthèse membranaire intense fait suite à la
cytodiérèse.
Remarques:
* Des données cytologiques récentes tendent à
montrer que le chondriome (ensemble des mitochondries d'une cellule)
pourrait notamment être formé d'un réseau de
tubules dynamiques anastomosées comme l'est le REL. C'est par
exemple le cas chez la levure.
* Les microtubules forment un réseau qui rayonnent à
partir d'une structure particulière qualifiée de
"centre organisateur des microtubules" qui, chez les cellules
animales, comporte deux petits cylindres protéiques creux
disposés perpendiculairement l'un à l'autre que l'on
appelle centrioles. Le centre organisateur des cellules
animales comportant deux centrioles est appelé
centrosome.
* Les microtubules sont des structures dynamiques qui s'allongent ou
raccourcissent très rapidement et continuellement. Les
microtubules accrochés au centromère des chromosomes
(fibres kinétochoriennes accrochés par le
kinétochore à des parties spécifiques du
centromère) se raccourcissent ainsi en tirant les chromosomes
fils vers les pôles où sont situés le centre
organisateur ou le centriole dans la cellule animale.
* De protéines motrices semblent pouvoir se déplacer le
long des microtubules en transportant de grosses molécules,
des particules protéiques ou des organites.
* Les cils et flagelles sont des composés complexes de
microtubules associés à des protéiques
particulières.
* Le début de la division cellulaire peut se repérer
dans sa phase la plus précoce par le changement d'organisation
du réseau des filaments d'actine du cytosquelette, ce qui se
fait en fin de phase G2 pour la cellule des plantes, avant que les
chromosomes se condensent et marquent le début de la
prophase.
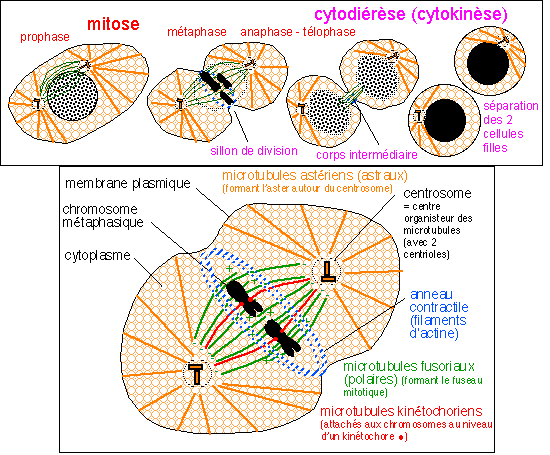
|
La division d'une
cellule animale (phase M du cycle cellulaire) comprenant
typiquement :
- la mitose
(division nucléaire)
- et la
cytodiérèse ou cytokinèse
(division cytoplasmique).
Les événements
chromosomiques, voir ci-dessus, n'ont pas été
détaillés.
Le cytoplasme, moins connu, très encombré
et structuré n'a pas non plus été
détaillé:
seuls les microtubules, les filaments d'actine
et les centrioles ont été
représentés.
|
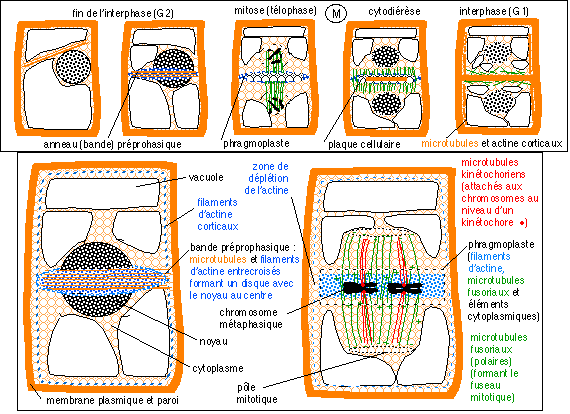
|
Division cellulaire d'une cellule
de plante (phase M comprenant la mitose et la
cytodiérèse).
La fin de l'interphase précédant cette
division et le début de l'interphase qui la suit ont
été représentées.
Deux étapes (fin d'interphase (G2) et début de
phase M (anaphase de mitose)) ont été
détaillées car la complexité est plus
grande que pour une cellule animale.
On retiendra le rôle essentiel des
microtubules et des filaments d'actine dans les
mouvements déterminant le plan de division, la mitose
et la cytodiérèse.
|
Il semblerait cependant que dans le cas de la cellule
d'une plante la formation de la plaque cellulaire soit au moins tout
aussi importante que la bande préprophasique pour la
détermination du plan équatorial de
division.
Remarques:
- un petit aperçu sur la division dans le monde
vivant.
Afin de survoler cette question, on peut traiter les facteurs
d'unité et les facteurs de diversité. On peut aussi
la regarder du point de vue de l'ADN et du point de vue
cytoplasmique, du point de vue de l'origine (cellules
impliquées) ou du devenir (des cellules filles).
|
Procaryotes
|
unité
|
diversité
|
|
une cellule mère se divise et
donne naissance à deux cellules filles
|
La cellule procaryote se
divise, se transforme en spore ou meurt. La division des
Procaryotes est une division binaire ou division
par scissiparité. La boucle d'ADN se
duplique en deux boucles qui se sépareraient
par allongement de la cellule du fait que chaque boucle
(l'ancienne et la nouvelle) serait accrochées
à la membrane en un point différent. Les
ribosomes et autres éléments cytoplasmiques
seraient plus ou moins également répartis
dans les cellules filles.
|
|
Protistes
|
La cellule eucaryote
présente des périodes de division courtes
(environ 1h) et des périodes de repos, plus
longues. La division cellulaire comprend deux
étapes qui se chevauchent plus ou moins : la
division nucléaire ou mitose et la division
cytoplasmique ou
cytodiérèse.
|
Les modalités sont
extrêmement variées: on parle de mitose
atypique pour désigner les divisions
nucléaires originales (certaines cellules
séparent leur chromosomes en étirant
l'enveloppe nucléaire, qui reste intacte, les
centromères étant accrochées
à sa membrane interne; des fuseaux de microtubules
traversent souvent le noyau sans que l'enveloppe
nucléaire ne se désagrège; chez les
levures et les Diatomées, l'enveloppe
nucléaire reste aussi intacte, le fuseau de
division se forme à l'intérieur du noyau
qui s'allonge et les chromosomes sont tirés par un
seul microtubule kinétochorien...).
|
|
Mycètes
|
Lors de la mitose, l'enveloppe
nucléaire se désagrège et un fuseau
mitotique se forme; les chromosomes sont tirés par
les microtubules kinétochoriens. Le centre
organisateur des microtubules n'est bien
individualisé (sous forme d'un centrosome
contenant deux centrioles) que chez les
animaux.
|
|
Animaux
|
|
Végétaux
|
|
matériel
génétique nucléaire
|
- dupliqué avant la division
(sauf exception)
- réparti équitablement dans les deux
cellules filles (sauf exception)
|
|
matériel
génétique mitochondrial ou
chloroplastique
|
transmis avec l'organite qui
habituellement se dupliquent et se répartissent de
façon à peu près égale entre
les deux cellules filles
|
|
cytoplasme et organites
cytoplasmique
|
le cytoplasme, avec ses organites et
après duplication d'un bon nombre de ceux-ci, est
réparti de façon équitable entre les
deux cellules filles
|
|
origine de la cellule
mère
|
une cellule mère peut
être une jeune cellule en fin de division (par
exemple lors de la multiplication cellulaire du
développement embryonnaire ou une cellule
bactérienne...) mais la cellule mère peut
aussi est une cellule quiescente qui est stimulée
brusquement u encore une cellule
différenciée qui peut se diviser en se
dédifférenciant ou non.
|
|
devenir des cellules filles
|
Certaines cellules filles meurent,
d'autres se divisent presque à la suite, d'autres
enfin entrent en quiescence...
|
- la fusion cellulaire:
on ne peut pas terminer un chapitre sur la division cellulaire
sans parler du mécanisme inverse: la fusion cellulaire.
Deux cellules peuvent fusionner en mettant en commun leur
cytoplasme et parfois leur matériel
génétique. On sait pratiquer des fusions in
vitro notamment avec la technique des hybridomes qui
consiste à faire fusionner une cellule cancéreuse
avec une cellule immunitaire de type lymphocyte B. les clones
ainsi obtenus peuvent produire de façon dirigée de
grandes quantités d'anticorps.
|
Tableau comparatif des divisions
cellulaires eucaryotes et procaryotes
|
|
caractères
|
procaryotes
|
eucaryotes
|
|
cellule
"animale"
|
cellule de
plante
|
|
matériel génétique
|
duplication
|
pendant la phase de croissance précédant la
phase de division
|
en interphase (phase S) avant la division (phase M)
|
|
séparation
|
par accrochage de chaque boucle d'ADN à la paroi
en croissance (phase de division)
|
par accrochage des chromosomes par les
kinétochores de microtubules spécifiques
(kinétochoriens) glissant le long des microtubules du
fuseau (anaphase de la mitose)
|
|
séparation des deux cellules filles
|
élaboration d'une paroi intermédiaire
|
étranglement cytoplasmique
|
élaboration d'une paroi intermédiaire
|
|
matériel responsable des mouvements
|
pas de mouvements autres que la croissance : pas de
microtubules ni de vésicules
|
microtubules et filaments d'actine, vésicules
golgiennes
centrioles dans le centre organisateur des
microtubules
|
microtubules et filaments d'actine, vésicules
golgiennes
pas de centrioles dans le centre organisateur des
microtubules mais une bande préprophasique puis un
phragmoplaste au niveau de la zone de partage cellulaire
|
|
moteur de la séparation (division
bactérienne ou cytodiérèse des
eucaryotes)
|
allongement de la paroi, rupture de la cellule
(synthèse de paroi et/ou déformation ????)
|
microtubules du cytosquelette puis rupture de la cellule
par étranglement
|
microtubules du cytosquelette pour tous les organites y
compris les chromosomes; la paroi ne venant séparer
que tardivement deux compartiments; pas de rupture de la
cellule mère mais un partage de l'espace
|
B1.1.2
- Chez les pluricellulaires la division conduit à une
différenciation
Les pluricellulaires, donc eucaryotes divisent leurs cellules
pour se développer (et croître), pour se
renouveller (renouvellement cellulaire) et secondairement pour
se reproduire. Le cycle cellulaire de leurs cellules n'est
donc qu'un des éléments du cycle de
développement (ou cycle de vie pour l'organisme
entier) qui contient aussi des phases de croissance, mais encore des
phases de différenciation et de sénescence.
du "cycle" cellulaire à la "toile" où à
l'arbre des filiations cellulaire:
Toute cellule étant issue d'une autre cellule (c'est un des
éléments de la théorie cellulaire, voir
cours de seconde), elle donne naissance
à une autre cellule ou meurt après un temps de vie plus
ou moins long. On a pris l'habitude de présenter la vie d'une
cellule comme un cycle, ayant comme point de départ sa
naissance et comme point d'arrivée une division; les cellules
qui n'atteignent pas ce point d'arrivée, qui est un nouveau
départ, meurent au cours du cycle. On peut y substituer une
représentation plus complexes en "toile" ou réseau.
Aucune voie n'est complètement fermée tant que la
cellule est vivante. Il n'en reste pas moins que certaines voies sont
nettement plus empruntées que d'autres. Dans la phase de
développement embryonnaire la plupart des cellules
nouvellement formées se divisent à nouveau, même
si parfois la division est inégale dans le sens où
seule l'une des deux cellules filles garde son pouvoir
mitogène, l'autre se différenciant (on parle de
division formative par opposition à une division
proliférative, où les deux cellules filles gardent
leur pouvoir mitogène). Les division chez un organismes adulte
sont nettement plus localisées (c'est le renouvellement
cellulaire), souvent dédié à des cellules
spécialisées (ou cellules souches) qui
présentent la plupart du temps des divisions formatives. En
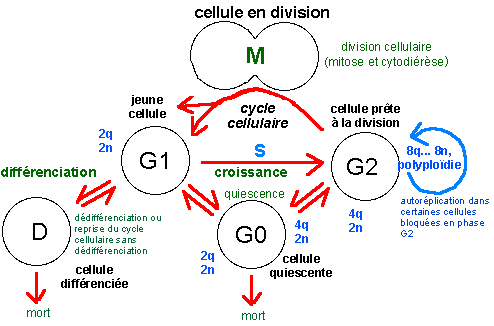
bleu, les quantités d'ADN par cellule (2q étant la
quantité unité dans une cellule diploïde) et
nombre de chromosomes (2n étant le nombre de chromosomes dans
une cellule diploïde); la phase S (synthèse) du cycle
cellulaire est la phase de réplication ou duplication de
l'ADN. La lettre G fait référence au mot anglais "gap"
désignant un fossé ou état stable de la cellule
(d'après la source citée, fig 1, p 498). Au cours du
cycle cellulaire on différencie l'interphase (qui
comprend les phases G1, S et G2) et la phase de division
cellulaire (M, qui comprend deux phénomènes plus ou
moins imbriqués: la mitose, ou division nucléaire, et
la cytodiérèse, ou division cytoplasmique) qui
correspond habituellement à moins de 10% de la durée du
cycle cellulaire.
Pour un organisme pluricellulaire le lignage de chaque
cellule de l'individu adulte a pu être reconstitué
notamment chez le vers Nématode Cœnorhabditis
elegans qui présente entre une dizaine et une vingtaine
d'étapes de divisions successives pour un total de 959
cellules somatiques (voir par exemple fig 14.15, p 510 et s. dans
Biologie du développement, Gilbert, 1996, De Boeck
Université - ce résultat spectaculaire a
été rendu possible par le petit nombre de cellules de
cet animal mais surtout par la transparence de sa cuticule et par la
disposition spatiale des territoires cytoplasmiques du zygote qui
semble fixe....!); on notera qu'au cours du développement
NORMAL de C. elegans 131 cellules subissent une mort naturelle ou
apoptose (voir page sur le
développement pour une explication de cette notion).
B1.2.1 - Chez
les plantes et les champignons la division n'est jamais suivi de
déplacement
a. La paroi n'est pas une boîte mais une
sécrétion sans cesse renouvellée: elle n'est
rigidifiée que dans certaines cellules
particulières
a1. la paroi des champignons
a2. la paroi des plantes
b. Quelques différenciations cellulaires
c. Des organes chez les plantes
B1.2.2 - Chez
les animaux certaines cellules se déplacent activement
Annexe: les jonctions
cellulaires