|
Plan |
|
|||
|
2. La nutrition,
une fonction globale
|
L'affirmation du programme: «Les activités fondamentales des cellules telles que le métabolisme et la division sont sous le contrôle d'un programme génétique» relève pour moi de la pétition de principe et je refuse de la colporter (certains manuels scolaires n'en touchent pas mot d'ailleurs (Belin) ou sinon sous forme d'exercice (Bordas, quoique la nouvelle édition en fasse un paragraphe du cours)). |
|||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||
|
Remarque |
||||||||||||||||||||||||||||||
|
Un
polymère est une molécule Des polymères... comme des colliers de perles (chaque perle est un monomère) |
homopolymères: ou -o-o-o-o-o-o-o-o-o-o-o-o-o-o- deux types d'homopolymères sont
possibles avec deux types de monomères
|
|||||||||||||||||||||||||||||
|
copolymères
: de très nombreux types de copolymères sont possibles avec deux types de monomères (cela dépend de la longueur du polymère) |
||||||||||||||||||||||||||||||
|
Une nourriture est ce qui permet à l'organisme de se nourrir. Elle doit comprendre de la matière et de l'énergie. La matière peut provenir soit de l'air soit des autres organismes, sans oublier la boisson qui est nécessaire pour tous. L'énergie peut être le soleil ou l'énergie chimique contenue dans les minéraux ou encore dans les composés organiques (pour des détails voir &2.4 de l'ancien cours d'immunologie de TS). |
||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
On regroupe les organismes par type trophique (du grec "trophein" = "se nourrir"). On désigne par là un type de travail de nutrition. |
"autotrophie" |
(autotrophie (au sens restreint au carbone) et phototrophie) Plantes, nombreux Protistes, certaines
bactéries (comme les bactéries bleues)
|
se nourrir d'air pour le carbone et de minéraux pour l'énergie (autotrophie (au sens restreint au carbone) et chimiolithotrophie) certaines bactéries
certaines bactéries
rares bactéries |
|||||||||||||||||||||||||||
|
Au sens restreint l'autotrophie est la capacité à trouver sa matière carbonée dans le dioxyde de carbone de l'air. Au sens large l'autotrophie englobe aussi la capacité à utiliser l'énergie lumineuse du soleil comme source d'énergie (phototrophie). |
Un être vivant peut se nourrir seul, c'est-à-dire sans l'aide d'autres êtres vivants, il est alors à proprement parler autotrophe (au sens étymologique de "se nourrit seul"). |
|||||||||||||||||||||||||||||
|
Au sens restreint l'hétérotrophie est la capacité à utiliser la matière organique comme source de matière carbonée. Au sens large l'hétérotrophie englobe aussi la capacité à utiliser l'énergie chimique (chimiotrophie) de ses proies vivantes ou mortes ou d'hôtes parasités ou associés en symbiose. |
se nourrir des autres hétérotrophie
au sens large |
ne se nourrir que des autres (hétérotrophie (au sens restreint au carbone) et chimiotrophie) Animaux, Mycètes, nombreux
Protistes, nombreuses bactéries
|
||||||||||||||||||||||||||||
|
Un être vivant peut se nourrir des autres de façon plus ou moins étroite, il est hétérotrophe au sens large ou, mieux, allotrophe (ceci est un néologisme formé à partir du grec "allo" = "les autres"). |
||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
Remarque (voir TP): |
Comment suivre expérimentalement le métabolisme ou déterminer simplement le type trophique d'un organisme ? |
|||||||||||||||||||||||||||||
|
Les notions qui suivent NE SONT PAS AU PROGRAMME, mais elles ont déjà été utilisées au collège; il ne s'agit ici que d'employer les mots sans approfondir les concepts; |
La photosynthèse (synthèse de matière organique à la lumière) fait partie du métabolisme des autotrophes phototrophes comme les plantes et de nombreuses bactéries. La réception de lumière (énergie lumineuse) et sa conversion en énergie chimique se font grâce à des pigments comme la chlorophylle (phase de phototrophie) et s'accompagnent de rejet de dioxygène suite à la coupure des molécules d'eau (photolyse de l'eau). L'énergie chimique sert ensuite à la synthèse de matière organique à partir du CO2 qui est donc consommé, ce qui est le signe de l'autotrophie vis-à-vis du carbone. |
La respiration fait partie du métabolisme de nombreux organismes aussi bien autotrophes que hétérotrophes, phototrophes ou chimiotrophes. C'est un échange gazeux entre le milieu extérieur et l'organisme. L'organisme consomme du dioxygène et rejette du dioxyde de carbone. Le dioxygène est utilisé, et le dioxyde de carbone est produit, dans les réactions de dégradation (catabolisme) de substances organiques, essentiellement pour produire de l'énergie chimique pour la cellule (chimiotrophie). La plupart des cellules eucaryotes respirent puisqu'elles dégradent de la matière organique, quel que soit leur type trophique. Ainsi, même les cellules chlorophylliennes, que l'on pourrait croire autotrophes réalisent aussi une chimiotrophie. Il faut donc réserver ces types trophiques aux organismes et non à une seule cellule d'un pluricellulaire. |
||||||||||||||||||||||||||||
|
Du point de vue du bilan, et en termes d'échanges gazeux, photosynthèse (rejet de dioxygène, absorption de dioxyde de carbone) et respiration (rejet de dioxyde de carbone, absorption de dioxygène) semblent antagonistes (avoir des échanges opposés), mais les gaz absorbés et produits n'ont ni la même origine, ni le même rôle. Ce bilan sera repris dans le chapitre sur l'écosystème terrestre.
|
||||||||||||||||||||||||||||||
|
Sais-tu quels sont les
aliments essentiels de la plante, en plus de l'eau de
"boisson", qui ont permis au saule d'atteindre 74 kg en ne
consommant que 57 g de "terre".
|
Un petit exercice d'histoire... Jan Baptist Von Helmont (1577-1644), comme les chimistes de son époque, considérait que la terre n'est pas un élément, car elle résulte de la transformation de l'eau. Il démontra son hypothèse en faisant pousser un jeune saule dans une caisse de bois contenant une quantité de terre bien déterminée. Après arrosage, durant cinq ans, avec de l'eau de pluie filtrée sur tamis, il observa que le poids de l'arbre avait augmenté de 74 kg, tandis que celui de la terre n'avait diminué que de 57 g. La terre n'ayant accusé aucune variation sensible de poids, c'est donc l'eau qui s'est changée en bois et en racines, c'est-à-dire en substances solides que l'on qualifiait de « terre ». (in EU) |
|||||||||||||||||||||||||||||
|
Remarque (d'après Esquisse d'une sémiophysique, Thom, 1988, p 69-71): la morphologie des organismes repose sans aucun doute sur le travail de nutrition. |
* Pour les organismes de
petite taille, unicellulaires procaryotes et eucaryotes, la
forme globale est une boule (topologiquement),
plus ou moins allongée, avec plus ou moins de
prolongements et la nutrition demande soit un passage des
nutriments ou déchets à travers les structures
limitantes (paroi par exemple), soit une déformation
de cette limite (phagocytose, exocytose...).
|
* Pour les organismes de plus grande taille (pluricellulaires) les allotrophes doivent mettre en jeu des dispositifs de capture des proies (tentacules, lacets, filets, pièces buccales...) qui vont de pair avec des organes locomoteurs et sensoriels de plus en plus sophistiqués; alors que les autotrophes s'identifient à leur milieu environnant en occupant l'espace, on croissant vers la lumière, ce qui fait que leur forme est ramifiante (leur croissance est dite indéfinie, car elle ne semble pas avoir de limite nette dans l'espace). Les botanistes ont depuis longtemps modélisé la croissance des plantes avec des algorithmes basés sur les fractales. |
||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
*La croissance est un terme non
spécifique aux biologistes qui désigne
l'augmentation d'une grandeur au cours du temps. |
||||||||||||||||||||||||||||||
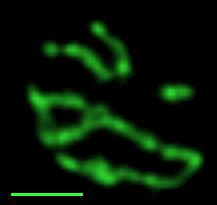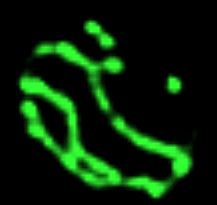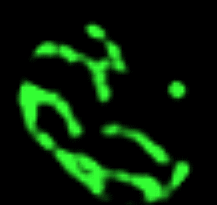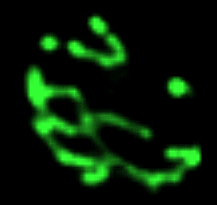 Réseau mitochondrial de la levure de bière Les prises de vue qui défilent
sont espacées de 3 min; |
Nous nous efforcerons de ne pas confondre ici la muliplication cellulaire et la division cellulaire (voir cours suivant de seonde) Diviser = séparer en plusieurs parties Multiplier = reproduire,
ajouter un nombre à lui-même une certain nombre
de fois...
|
|||||||||||||||||||||||||||||
|
Les nombreuses variations en nombre
(multiplication, mais aussi
fusion) et en forme des organites des cellules
eucaryotes en fonction de leur état de nutrition est
un domaine encore peu exploré. On ne peut
s'empêcher de citer les magnifiques images des
variations du réseau tubulaire mitochondrial de la
levure de bière par Gustavo Pesce et Peter Walter
- du CDROM de Biologie
Moléculaire de la Cellule (Flammarion-Médecine
Sciences) - (non trouvés sur
le net mais il reste celle de Jodi
Nunnari (http://www.ucsf.edu/sedat/Images/
mitoanim.gif),
nettement moins belles mais tout de même parlantes :
les photos sont prises en fluorescence (mito-GTP), chaque
sous-image gif est séparée de 3 min, le
fichier gif contient 14; pour mieux observer le
phénomène j'ai porté à 5
secondes le temps d'affichage de la première image
avec la barre d'échelle de 2 µm, puis chaque
image suivante est séparée d'une seconde
(pour ceux qui auraient accès
à l'article de référence voir Shaw JM,
Nunnari J, 2002, Mitochondrial dynamics and division
in budding yeast. Trends Cell Biol. 2002
Apr;12(4):178-84.PMID: 11978537)
|
||||||||||||||||||||||||||||||
|
Chez un procaryote qui forme une colonie (la plupart des bactéries) les multiplications cellulaires peuvent être regardées non pas comme des multiplications de reproduction, mais comme des multiplications de croissance ou de développement (de la colonie). C'est la colonie bactérienne qui est le véritable individu (multicellulaire) et chaque bactérie n'est qu'une cellule de cet individu. C'est la thèse développée notamment par James Shapiro (voir par exemple http://shapiro.bsd.uchicago.edu /Shapiro98AnnRevMicro.pdf). |
La multiplication bactérienne et ses caractéristiques est à mon avis bien mieux placée ici qu'en 1èreS où l'on est obligé de la traiter en plus de la mitose. Ce qui permettrait alors en 1ères S de se focaliser sur la seule question de la réplication de l'ADN qui finalement est la seule au programme. Et pour les secondes se serait une ouverture à une compréhension plus large de la multiplication cellulaire. |
|||||||||||||||||||||||||||||
|
Chez les eucaryotes unicellulaires, les multiplications sont considérées comme des multiplications de reproduction à moins que l'on considère le niveau de la colonie. Cependant, c'est clairement l'état nutritif de la cellule qui est le déclencheur de la multiplication. |
La
"mitose" vient aussi tout naturellement ici comme
phénomène de croissance et de renouvellement
cellulaire, ce qui permet d'éviter la
réduction par les élèves de
l'héritage aux mécanismes
génétiques qui ne devraient pas être
traités avant la terminale : que l'on parle de
chromosomes, oui, mais pas de gènes.
|
|||||||||||||||||||||||||||||
|
Chez un eucaryote pluricellulaire, les
multiplications cellulaires (comprenant
une mitose
et donc souvent
qualifiées de "mitoses";
le mot sera défini plus loin
dans le cours
sur la multiplication cellulaire)
peuvent avoir plusieurs finalités: lors du
développement ce sont des "mitoses" de
croissance; à l'âge adulte et lors de la
sénescence ce sont plutôt des "mitoses" de
renouvellement. En tout cas, les "mitoses" n'ont pas de
pas de rôle reproductif, sauf dans le cas d'un
unicellulaire. Pour éviter d'employer le mot de
reproduction et pour signifier le grand nombre de divisions,
on parle de
prolifération
cellulaire. |
||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||
|
la multiplication bactérienne
|
Un peu
de philosophie: Si l'on considère maintenant l'individu comme multicellulaire, le problème ne se pose plus, chaque cellule n'est qu'un partie de l'individu. Certaines, dans des conditions particulières, peuvent transmettre la vie et former un nouvel individu (la reproduction est clairement séparée de la croissance). |
|||||||||||||||||||||||||||||
|
Les documents pour les bactéries sont peu nombreux du fait de la petite taille des cellules, mais il existe de très bons films pris en microscopie optique avec ou sans fluorescence. Les films de Shapiro sont superbes: par exemple : http://shapiro.bsd.uchicago.edu/ singlecell_microcol_08_88.mov ; mais ils manquent d'explications. On peut aussi voir le site commercial de
cells alive (http://www.cellsalive.com/
qtmovs/ecoli_mov.htm)
|
||||||||||||||||||||||||||||||
|
Des explications en français sont à l'adresse http://www.forumlabo.com/ 2006/actus/actus/INSERM/ 0305bacterie.htm. |
|
|||||||||||||||||||||||||||||
|
la multiplication des cellules eucaryotes |
http://www.bio.unc.edu/ faculty/salmon/lab/mafia/ mafiamovies.html |
|||||||||||||||||||||||||||||
|
Pour les unicellulaires
chez Dictyostelum discoïdum : une amibe acrasiale http://www.bio.unc.edu/ faculty/salmon/lab/ mafia/bigrc.mov la phase terminale de la division chez Sacharomyces cerevisiae (double vue en fluorescence avec marquage des protéines de type actine (anneau de division), et en contraste de phase): http://www.bio.unc. edu/faculty/salmon/lab/ mafia/contr.mov |
Pour les pluricellulaires
chez l'oursin: http://www.bio.unc.edu/faculty/ salmon/lab/mafia/ urchin.mov une belle mitose (un peu partielle) chez le rat kangourou: http://www.bio.unc.edu/ faculty/salmon/lab /mafia/phmit1.mov avec marquage de l'actine par une protéine GFP de cellules épithéliales du foie de porc (il vaut mieux visionner les images une à une... le film est un peu rapide): http://www.bio.unc.edu/ faculty/salmon/lab/ mafia/gfpactinsheldon.mov les 4 premières divisions du zygote chez Cœnorhabditis elegans: http://www.bio.unc. edu/faculty/salmon/ lab/mafia/wt.mov les idées à retenir: cellules reproductrices spécialisées. On ne reproduit pas un individu pluricellulaire à partir de cellules quelconques... |
|||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
2.4.1 La "mitose" est un événement de type catastrophique (au sens mathématique) et peut être modélisée |
Une présentation a été faite lors de la conférence de 10/2005 à Poitiers
|
|||||||||||||||||||||||||||||
|
La division de la cellule peut être modélisée (de façon extrêmement approximative, mais particulièrement intéressante) par l'apparition d'une cassure (singularité) dans la forme de la cellule (assimilée ici à une boule) due à un décalage entre les flux d'énergie entrant et sortant. Le modéle devrait bien sûr être amélioré par d'autres flux (informatifs par exemple)... « Dans un milieu extérieur
E où règne un flux continu d'énergie en
voie de dégradation thermique, on suppose
donnée une boule B de rayon r, où règne
un régime dynamique local g consommateur
d'énergie. Admettons que la consommation
d'énergie soit proportionnelle au volume ; la capture
d'énergie, provenant des échanges avec le
milieu extérieur, n'est alors proportionnelle
qu'à la surface. Si le rayon r est au départ
assez petit, il y aura excès de l'énergie
capturée sur l'énergie consommée ; on
admettra que cet excès d'énergie est
consacré à la synthèse de substances
support du régime g, de sorte que le volume V (donc
le rayon r) ira croissant. Si ce processus de croissance
était rigoureusement proportionnel à
l'accroissement d'énergie, le rayon r irait en
croissant asymptotiquement vers un rayon r0, valeur
d'équilibre pour laquelle l'énergie
capturée est égale à l'énergie
consommée. Supposons alors que ce mécanisme de
synthèse présente une certaine inertie, un
certain retard. Il pourra alors y avoir accroissement du
volume au-delà du volume optimal, au détriment
de la consommation d'énergie interne ; la boule, de
régime g et de rayon r > r0 entre alors dans un
état d'instabilité dont elle ne pourra sortir
que par cassure ; après fragmentation en deux boules
de petit rayon r1, plus petit que r0, la croissance pourra
reprendre, suivie de multiplication ».
(René Thom, 1968
(Stabilité structurelle et morphogenèse; essai
d'une théorie générale des
modèles, stabilité.pdf, p 326))
|
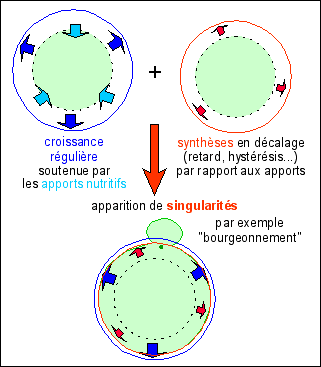 |
|||||||||||||||||||||||||||||
|
Il est frappant, lorsque l'on observe attentivement une mitose au microscope de voir comment lors de la division les deux cellules filles sont SPHÉRIQUES et ne se réaccolent qu'ensuite. |
Des images sont aussi très
parlantes sur le site BIOCLIP,
notamment celles de Manuel THERY qui a
travaillé sur la physique de la division en orientant
celle-ci au moyen d'adhésifs
géométriques (chercher Manuel Thery pour voir
les vidéos et choisir Randomly oriented cell
division). |
|||||||||||||||||||||||||||||
|
2.4.2 Les chromosomes sont les organites de la mitose |
||||||||||||||||||||||||||||||
|
voir page Les chromosomes |
2.4.2.1 - Les chromosomes sont de petits bâtonnets colorables |
|||||||||||||||||||||||||||||
|
Il n'y a pas de chromosomes au sens cytologique chez les Procaryotes. Le nucléoïde bactérien présente un ADN compacté avec des protéines, mais l'ensemble est instable. Les protéines s'y associent et se dissocient en quelques minutes. D'une façon plus profonde, la question est de savoir si le nucléoïde bactérien est l'équivalent d'un unique chromosome lors de la division bactérienne. René Thom y répond par l'affirmative (de même pour l'ADN des mitochondries ou des chloroplastes). |
Un chromosome est
étymologiquement une particule colorable (du grec
chromos = la couleur et soma = le corps). Ces
particules ont été observés et
décrites en 1879 par Flemming lors de la division de
cellules animales. Depuis des chromosomes ont
été observés lors de la division
cellulaire des toutes le cellules eucaryotes.
Les chromosomes se colorent à
l'aide de colorants basiques (parce qu'ils sont
composés de molécules acides). Mais si l'ADN
peut se colorer, même sans être à
l'état de chromosomes, pour que l'on puisse le voir
au MO il est nécessaire qu'il soit condensé
dans un chromosome.
Maintenant on sait colorer spécifiquement chaque chromosome par des colorants fluorescents et le suivre de loin lors de la division cellulaire. C'est aussi un moyen très rapide pour compter les chromosomes dans la cellule. (taille, nombre, forme, "comportement:
apparition-disparition")
|
|||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
L'ADN est associé à des
protéines dans les chromosomes. |
Les histones (protéines
basiques) structurent l'ADN dans la chromatine , c'est
là qu'on les étudie. Mais on pense qu'elles
formeraient aussi le squelette du chromosome, au moins
partiellement. |
|||||||||||||||||||||||||||||
|
2.4.2.3 - L'ADN est une molécule filamenteuse |
||||||||||||||||||||||||||||||
|
Les acides nucléiques sont des polymères (du grec poly = plusieurs et merein = partage) (voir ci-dessus) car il sont composés d'unités identiques (les monomères). Comme il peut y avoir plusieurs types de nucléotides, on parle de copolymères. |
Les
désoxyribonucléotides au sens strict
(ou les nucléotides au sens large) sont
des molécules composées d'une base
azotée (A= l'adénine, T= la thymine, C=la
cytosine, G=la guanine), d'un sucre (le désoxyribose)
et d'un, de deux ou de trois groupement(s) phosphate(s). Les
désoxyribonucléotides ont de nombreux autres
rôles dans le métabolisme
(voir cours de 1ère et
TS).
|
|||||||||||||||||||||||||||||
|
L'ADN (acide
désoxyribonucléique) est formé de
deux chaînes (brins) de nucléotides
dessinant une double hélice dont le
diamètre est de 2 nm. |
||||||||||||||||||||||||||||||
|
TD - ADN; forme, structure des molécules; simualisation* 3D (attention applet Jmol de 540Ko à télécharger) (* les superbes applications qui visualisent les molécules en 3D sont des simulations; pour souligner ce fait, je parlerai de simualisation) |
||||||||||||||||||||||||||||||
|
2.4.2.4 - Les protéines sont des molécules filamenteuses |
||||||||||||||||||||||||||||||
|
Il existe 20 acides aminés
dans les protéines du vivant. Ce sont des
molécules organiques donc composées de C, H,
O, N et parfois S. Elles ont toutes deux groupes d'atomes
que l'on appelle des radicaux et qui leur donnent leur nom:
un radical acide (-COOH) et un radical
amine (-NH2).
|
Les protéines font partie du groupe plus vaste des peptides. Le terme de peptide désigne toute chaîne d'aa (aa réunis par des liaisons covalentes). Les oligopeptides sont formés quelques aa (10...); les polypeptides sont formés de nombreux aa (10-100) mais lorsque leur poids moléculaire est supérieur à 10.000 on préfère parler de protéines. Les protéines sont souvent formées de l'assemblage de plusieurs molécules et contiennent des éléments non peptidiques. Cette distinction faite on utilisera en seconde le mot protéine comme équivalent de peptide. |
|||||||||||||||||||||||||||||
|
2.4.2.5 - On pense que chaque chromatide ne contient qu'un seul filament d'ADN qui serait répliqué avant la division |
||||||||||||||||||||||||||||||
|
L'ADN est très
compacté dans un chromosome. Dans une cellule
humaine il y a 46 chromosomes (regroupées en 23
paires de même forme et taille) soit environ 2 m d'ADN
déroulé. Chaque chromosome humain ne
dépasse pas quelques µm de longueur.
|
||||||||||||||||||||||||||||||
|
Avant chaque division cellulaire l'ADN est dupliqué (c'est la réplication de l'ADN), ce qui permet de répartir un même matériel génétique dans les deux cellules filles issues de la division cellulaire. Pour les procaryotes la boucle d'ADN est répliquée en deux boucles réparties dans les deux cellules filles. Pour les eucaryotes la réplication de l'ADN donne des chromosomes doubles (les deux parties identiques de chaque chromosome ou chromatides sont attachées au niveau d'un centromère). La cellule les scinde en deux puis répartit chaque chromatide dans une cellule fille (c'est la mitose). Elle divise ensuite son cytoplasme (c'est la cytodiérèse).
|
||||||||||||||||||||||||||||||
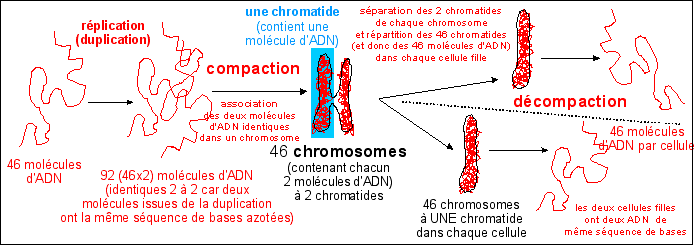 Duplication, compaction, association, séparation, décompaction de l'ADN dans une cellule humaine qui se divise en deux cellules filles. Le chromosome comme structure TRANSITOIRE lors de la division cellulaire d'un eucaryote (formation des chromosomes par compaction de l'ADN, séparation des chromatides, disparition des chromosomes par décompaction de l'ADN) |
||||||||||||||||||||||||||||||
|
Remarque 1 : |
|
|||||||||||||||||||||||||||||
|
Remarque 2 : |
* l'explication la plus courante est
celle fournie ci-dessus: les
chromosomes sont des structures (protéiques) de
compaction et de séparation de plusieurs
molécules immenses d'ADN. Sans
chromosomes les 92 molécules d'ADN d'une cellule
humaine en fin de réplication formeraient un fouillis
inextricable.... |
|||||||||||||||||||||||||||||
|
2.4.2.6 - Il existe des anomalies chromosomiques Les anomalies touchent le nombre, la taille, la forme... Dans une cellule diploide les chromosomes
vont par paire. Certains chromosomes peuvent être en
trois exemplaires (triploïdie)....D'autres peuvent
avoir perdu ou gagné un bras...
|
Il est essentiel de comprendre que l'on ne fait pas habituellement de caryotypage chez des individus sains, et que l'on ne compare pas le caryotype de toutes les cellules d'un même individu. L'idée théorique actuellement encore la plus répandue est que "toutes les cellules d'un organisme ont le même caryotype". Ce n'est plus une évidence. Il est inexact de dire que les anomalies chromosomiques sont la cause de ces maladies. On ne peut que dire que ces maladies sont associées à des anomalies chromosomiques. Il pourrait s'agir plutôt de conséquences des fonctions cellulaires sur la formation des chromosomes lors d'une division. |
|||||||||||||||||||||||||||||
|
La première anomalie chromosomique à avoir été mise en relation avec une maladie a été la trisomie 21 associée au syndrome de Down par Lejeune, Gautier et Turpin en 1959 (voir ancienne page). |
D'autres maladies associées à des anomalies chromosomiques ont rapidement été détectées et il n'a pas fallu longtemps pour que, étant donné les développements de la théorie chromosomique de l'hérédité (voir cours de TS), l'on qualifie ces maladies de maladies chromosomiques identifiant ainsi de façon abusive un caractère avec la cause. |
|||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
|
On peut dire que le noyau contient les chromosomes déroulés ou que l'ADN et les protéines nucléaires se condensent en chromosomes lors de la division cellulaire |
||||||||||||||||||||||||||||||
|
On a utilisé des
colorants fluorescents spécifiques de chacun des
chromosomes chez une petite "biche" femelle d'Asie (le
Muntjac) qui possède 6 chromosomes (les
cellules sont des fibroblastes).
|
Le noyau de gauche est au repos et on distingue les territoires de chaque chromosome, bien séparés; il faut donc comprendre le matériel génétique comme une forme dans un espace propre et non seulement comme une molécule chimique. Le passage de la forme
déroulée à la forme filamenteuse
(chromosome) peut s'expliquer par un modèle
mathématique où les chromosomes sont les
singularités associées à la catastrophe
mitotique (voir René
Thom). |
|||||||||||||||||||||||||||||
|
|
Un article en anglais accessible sur le net librement (licence Creative Commons Attribution) est à l'adresse http://biology.plosjournals.org/archive/1545-7885/3/5/pdf/10.1371_journal.pbio.0030157-S.pdf. en voici un extrait d'image (Fig. 1-D - réf: 10.1371/journal.pbio.0030157.g001) particulièrement accrocheur: 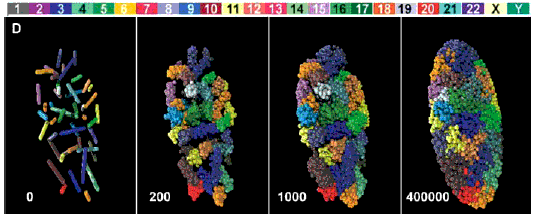 Le positionnement des 46 chromosomes (dans un état de décondensation croissant de la gauche vers la droite) d'un fibroblaste humain en reconstitution 3D simulant les résultats obtenus par microscopie (voir l'article pour des précisions) J'aime beaucoup la légende ajoutée par La Recherche (brève du n°388, juillet-août 2005, 15): Paysage nucléaire, qui rappelle le paysage épigénétique de Waddington (voir page annexe). |
|||||||||||||||||||||||||||||
|
Références
de l'article:
Three-Dimensional Maps of All
Chromosomes in Human Male Fibroblast Nuclei and Prometaphase
Rosettes |
||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||
{kind=link}