|
3. La reproduction
transmet la vie
|
Tout
d'abord, la reproduction est définie ici comme
fonction globale du vivant (voir Qu'est-ce
que la vie
?)
c'est-à-dire au niveau d'un organisme et non d'une
cellule unique d'un pluricellulaire. |
|||||||||||||||||||||||||||
|
3.1 - La reproduction est une multiplication des individus au sein d'une espèce |
||||||||||||||||||||||||||||
|
|
L'évolution est l'histoire de la vie ou de la transformation des espèces au cours des temps géologiques... |
|||||||||||||||||||||||||||
|
* le vocabulaire du mathématicien René Thom est expliqué sur la page des 4 causes. |
La reproduction n'est pas une création d'un être vivant à partir d'un autre être vivant mais la création d'un nouvel être vivant au sein d'une espèce. Dans le cadre de la vision continue de la vie et de l'espèce, la reproduction d'un individu au sein de l'espèce n'est donc plus la capacité de créer un nouvel être vivant à partir d'un individu, ce qui va fortement contre l'idée que le moins ne peut donner le plus; mais si c'est la réalisation des conditions nécessaires à l'apparition d'une nouvelle vie individuelle - saillance* - (au sein du continu de l'espèce - prégnance*), cela devient beaucoup plus simple. Comme la notion de reproduction et celle d'espèce sont indissociables, la notion d'espèce est donc inséparable de la notion d'évolution. Définir l'évolution au niveau d'un seul individu n'est pas possible, même si ce sont des individus que l'on compare, soit au sein d'une même espèce, soit entre espèces que l'on considère comme différentes. |
|||||||||||||||||||||||||||
|
Pourquoi a-t-on besoin de parler d'espèce ? Pourquoi ne parle-t-on pas de reproduction au seul niveau d'un organisme ? Parce que cette fonction globale n'est pas toujours le propre d'un organisme mais est toujours celle de l'espèce. Non pas dans la capacité à se reproduire, qui est bien une fonction de l'individu, mais dans son utilisation (on est pas obligé de se reproduire pour être totalement vivant). C'est le cas de nombreux unicellulaires (les amibes acrasiales par exemple) et même de bactéries (bactéries dites sociales) qui vivent en colonies et qui peuvent réaliser cette fonction non pas chacun séparément mais en la "déléguant" à quelques individus. On considère que c'est un premier pas vers la pluricellularité. C'est donc une propriété incontournable de la société du moins dans sa globalité. Un individu peut ne pas se reproduire et dans ce cas sa vie s'arrête à sa mort. Mais c'est impossible pour une espèce car sinon cela signifie qu'il y a extinction. Une extinction signifie qu'aucun descendant ne perpétue l'espèce. On voit que la notion d'espèce, comme population interféconde dont les individus se transmettent la vie sous une forme particulière (species en latin c'est "ce qui se voit", c'est-à-dire la "forme visuelle" au sens le plus courant de "forme"), est indissolublement liée à la reproduction. On arrive ainsi tout naturellement aux deux autres fonctions globales de relation et de nutrition qui sont aussi liées à l'espèce mais peut-être de façon moins évidente. Dans sa définition de l'espèce, qui date de 1989, Mayr fait mention de la niche écologique. Le mot écologique dérivant du grec oïkos (habitat) on peut non seulement y voir l'espace, le lieu, mais aussi les habitants. Donc les fonctions de relations entre les êtres vivants sont très certainement comprises dans cette définition. De la même manière la nutrition comporte toujours un volet relationnel que cela soit pour la recherche de l'énergie lumineuse ou la capture des proies. On peut, à mon avis, avancer comme argument, que la délégation de ces deux fonctions à des cellules spécialisées au cours de l'évolution, tout comme pour les fonctions de reproduction, est un indice de cette socialisation en cours chez les pluricellulaires. |
||||||||||||||||||||||||||||
|
|
Des lions, des chiens, des
tigrons....des bactéries, des plantes, des
insectes... Les espèces apparaissent et disparaissent au cours de l'évolution. |
En linguistique c'est le nom de "genre" qui est employé pour désigner un concept comme "table" ou "chaise". Les éléments particuliers, "telle table", "cette chaise", correspondent aux individus du vivant. Voir Thom, le genre est continu (comme la vie), l'espèce est discrète. |
||||||||||||||||||||||||||
|
|
En fait on peut même parler de forme au sens philosophique d'Aristote du principe de vie de la matière (il divise l'être de toute corps matériel en matière et forme). On pourrait donc dire que l'organisme qui se reproduit donne vie à un nouvel être vivant composé de matière et de forme. La matière est en partie héritée (chromosomes, cytoplasme...) et en partie nouvelle (synthétisée selon le type trophique à partir de matière minérale ou organique venant du milieu extérieur...). La forme est elle aussi en partie héritée (forme génétique, ou information génétique, forme cytoplasmique ou information cytoplasmique et forme environnementale, ou information environnementale) et en partie nouvelle (déformation, transformation....tous ces mots désignant des processus pouvant modifier la forme à transmettre par hérédité). |
|||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
Remarque: |
Comme on distingue 2 grands types trophiques: l'autotrophie et l'allotrophie, on peut distinguer deux grands types de reproduction. * je précise que ces mots sont des néologismes, plus ou moins heureux, inventés pour illustrer une comparaison qui me semble pertinente. |
La reproduction asexuée ou autogenèse*, est solitaire. Dans le cas le plus complet, elle conduit à la mort de l'être vivant géniteur. Le plus souvent l'individu père survit à la reproduction et peut engager plusieurs reproductions successives. |
La reproduction sexuée ou hétérogenèse*, demande l'intervention des autres êtres vivants de la même espèce (et de sexe opposé). "hétéro", comme différent est préférable à "allo", l'autre. |
|||||||||||||||||||||||||
|
3.2 La reproduction asexuée est solitaire |
||||||||||||||||||||||||||||
|
On peut éventuellement associer à la reproduction asexuée les phénomènes de régénération qui sont une capacité de régénération complète ou partielle d'un individu à partir d'un fragment. Cette propriété, quasiment générale chez les Plantes reste assez rare chez les animaux. Une régénération totale est possible chez l'hydre mais elle n'est que partielle chez une Annélide ou une étoile de mer (Echinodermes Stellérides: un bras par exemple), ou encore très localisée comme chez les Crustacés (une pince par exemple). |
Chez les eucaryotes unicellulaires les
modalités cytologiques de la reproduction
asexuée sont extrêmement variées et il
ne faudrait pas croire que la division cellulaire vue dans
le chapitre sur la nutrition rend compte de cette
variété: la mitose binaire
(Paramécie, Trypanosome...) n'est peut-être que
l'exception parmi les mitoses multiples ou
schizogonies (Plasmodium...) et autres
bourgeonnements (Levure...). De nombreuses
mitoses "atypiques" ont été rapportées
chez les Protistes. |
Il existe de très nombreux
organismes, mêmes pluricellulaires, pour lesquelles on
ne connaît pas de reproduction sexuée. Cela ne
veut pas dire qu'elle n'existe pas. Remarque: |
||||||||||||||||||||||||||
|
3.3 La reproduction sexuée se fait à deux individus de sexe différent ou porteurs des deux sexes |
||||||||||||||||||||||||||||
|
C'est bien par comparaison avec la
reproduction humaine que l'on désigne les
différentes modalités de la reproduction chez
tous les organismes vivants. Lors de la reproduction
sexuée deux organismes (qui peuvent être
unicellulaires) de sexe opposés (ou qui
possèdent les deux sexes) produisent lors de la
gamétogénèse des cellules sexuelles (ou
gamètes). Lors de cette
gamétogénèse a lieu une série de
divisions particulières dont la
méïose constitue le
phénomène le plus remarqué. La
rencontre des sexes permet celle des gamètes qui
fusionnent lors de la fécondation. La
fécondation est suivie du
développement qui conduit à la
maturité sexuelle, capacité à produire
de nouveau des gamètes. Pour de nombreux organismes
le développement peut se faire au sein de l'organisme
maternel et les géniteurs peuvent encore longtemps
assurer la nutrition et la protection de leurs
descendants. |
Remarques: |
|||||||||||||||||||||||||||
|
La méïose est une suite de deux divisions successives qui, à partir d'une cellule mère donne typiquement 4 cellules sexuelles ou gamètes. |
On qualifie de femelle la cellule
sexuelle la plus grosse et habituellement
immobile (ovocyte de ovo =
œuf et cyto = cellule) alors que la cellule
sexuelle la plus petite et mobile est appelée
mâle (spermatozoïde). |
La fécondation est l'union de deux gamètes de sexe opposé, typiquement un ovocyte, femelle, et un spermatozoïde, mâle, en une seule cellule: le zygote. Le développement est la transformation du zygote en organisme capable de produire des gamètes. |
Le cycle
biologique (ou cycle de vie ou encore cycle
de reproduction) d'une espèce
regroupe toutes les étapes depuis le zygote
jusqu'à la formation d'une nouveau zygote à
partir d'un ou de plusieurs individus
(ou parle du cycle de vie de la
grenouille ou du cycle de vie de l'homme mais pas de son
cycle de vie en tant
qu'individu). |
|||||||||||||||||||||||||
|
Même chez les unicellulaires on parle de développement car la cellule issue de la fécondation (le zygote) se transforme avant de pouvoir donner des gamètes, même si cette étape est parfois plus réduite que la vie des gamètes. On s'est rendu compte notamment que du point de vue génétique le développement s'accompagnait de "l'activation de gènes particuliers" aussi bien dans une cellule unique que dans les différents tissus d'un pluricellulaire. |
Seules les cellules sexuelles issues d'une méïose sont capables se fusionner naturellement entre elles lors de la fécondation. Ce sont des cellules particulières, notamment au niveau du nombre de leurs chromosomes. On est capable artificiellement de fusionner des cellules non sexuelles (réalisation d'un hybride) mais jamais on obtient d'embryon capable de se développer en individu complet. Par contre on peut obtenir, dans certains cas, des individus complets à partir de manipulation d'une seule cellule sexuelle. On connaît des cas naturels de développement à partir d'une seule cellule sexuelle: on parle de gynogénèse pour une cellule initiale femelle et d'androgénèse pour une cellule initiale mâle, ce qui est beaucoup plus rare. Dans le cas du clonage de la brebis Dolly on a fusionné (artificiellement) un ovocyte énucléé avec une cellule non sexuelle. |
Les modalités de la reproduction sexuée sont tellement différentes chez les organismes des 4 royaumes eucaryotes qu'il est difficile de faire un schéma global. Il existe par exemple des organismes à la fois mâles et femelles (comme chez de nombreuses plantes à fleurs) mais la plupart du temps les individus s'échangent des gamètes mâles; ils ne pratiquent pas ainsi l'autofécondation (se féconder soi-même). La fécondation peut être externe (la grenouille mâle arrose de son sperme les ovocytes pondus par la femelle) ou interne (ce qui nécessite souvent un organe de copulation). La plupart du temps les embryons se développent dans le milieu extérieur mais avec des protections particulières. |
||||||||||||||||||||||||||
|
Chez les pluricellulaires - et dans une certaine mesure chez certains unicellulaires coloniaux - le développement s'accompagne de la différenciation : apparition de différents types cellulaires. |
On peut comprendre ce mot en deux sens : - d'une part la différenciation morphologique: les cellules changent de forme. - d'autre part la différenciation fonctionnelle : certaines cellules ne réalisent plus la totalité du travail cellulaire, il y a division du travail. Une différenciation cellulaire fonctionnelle (et secondairement morphologique) est observée chez certains unicellulaires coloniaux comme les Volvox qui différencient des cellules nutritives et relationnelles et des cellules reproductives. Une différenciation cellulaire morphologique (et secondairement fonctionnelle) est observée chez d'autres unicellulaires coloniaux comme les amibes acrasiales qui forment un appareil fructifère. La limite entre changement de forme et différenciation n'est pas simple: de nombreuses levures par exemple sont connues pour présenter des formes sporulées ou filamenteuses selon les conditions d'environnement. Tant que le changement de forme est
réversible il est difficile de parler de
différenciation.
|
|||||||||||||||||||||||||||
|
À partir du moment où l'organisme survit à l'émission des gamètes, il se pose le problème de l'origine des cellules sexuelles capables de former les gamètes. |
L'idée de base est simple et résulte de la même vision que celle présentée plus haut pour les unicellulaires: les cellules qui donnent les gamètes sont éternelles puisqu'elles transmettent la vie de parents à enfants sans discontinuité autre que la méiose et la fécondation. Ces cellules sexuelles forment la lignée germinale par opposition aux cellules mortelles, les cellules somatiques (du grec soma: le corps). |
En fait, en culture, on sait actuellement maintenir en vie des cellules somatiques plus facilement que des cellules sexuelles. Le maintien en culture passe cependant par des possibilités de division et on observe toujours un vieillissement de la culture qui se solde par un arrêt des divisions, même pour les cellules cancéreuses qui, pourtant, se divisent très facilement spontanément. |
Remarque: l'opposition soma-germen a peut-être un sens en embryologie mais cette distinction me paraît artificielle chez un adulte; il faut peut-être lier ces termes à l'association du psychologique (de l'esprit) et du somatique qui a donné le terme psychosomatique qui s'efforce de décrire l'union indissociable de l'esprit et du corps dans la nature humaine cartésienne. |
|||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
TD Reproduction chez Cœnorhabditis elegans |
Cœnorhabditis elegans
est un organisme modèle qui a
donné lieu à d'innombrables documents
disponibles sur internet (http://www.wormatlas.org/links.htm
voir films division cellulaire: caryogamie et début
de première division: http://www.mpi-cbg.de/downloads/movie/wildtypePG.mov,
plusieurs autres divisions: http://www.bio.unc.edu/faculty/salmon/lab/mafia/wt.mov,
division de blastomères isolés:
http://www.bio.unc.edu/faculty/salmon/lab/isolated.mov
|
|||||||||||||||||||||||||||
|
Ancien TP - Fécondation et premières étapes du développement embryonnaire chez l'hermelle |
||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
Théorie du programme génétique
de développement
|
Théories du développement
autonome
|
|||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
|
L'exemple classique d'Acetabularia a été choisi par les collègues ayant conçu les manuels scolaires. Mais il s'est révélé délicat. Les présentations qui en sont faites dans les manuels sont souvent par trop inexactes et empêchent alors toute réflexion. Sans pouvoir accéder aux publications historiques, on peut s'efforcer cependant de présenter des données moins dépassées : un travail en cours. |
|||||||||||||||||||||||||||
|
En travaux |
expériences de Gurdon et autres expériences de transfert nucléaire: voir page d'histoire des sciences avec articles originaux: http://www.pnas.org/misc/classics4.shtml/ 1. Spemann, H. (1938) Embryonic Development and Induction (Yale University Press, New Haven). 2. Briggs, R. & King, T. J. (1952) Proc. Natl. Acad. Sci. USA 38, 455-463. |Article| 3. National Academy Press Biographical Memoir |Article| 4. Di Berardino, M. A. and McKinnell, R. G. (2001) Differentiation 67, 59 - 62. |Article| 5. Porter, K.R. (1939) Biol Bull Woods Hole 77, 233&endash;257. 6. Briggs, R. & King, T. J. (1957) J. Embryol. Exp. Morphol. 100, 269 - 312. 7. Fishberg, M., Gurdon, J. B., & Elsdale, T. R. (1958) Nature 181, 424. 8. Westhusin, M. et al. (2002) Nature 415, 859. |Article| 9. Hochedlinger, K. & Jaenisch, R. (2002) Nature 415, 1035 - 1038. |Article| 10.Wilmut, I. et al. (1997) Nature 385, 810-813. 11. Human Genome Project Fact Sheet 12. Shiels, P.G. et al. (1999) Nature 399, 316 - 317. |Article| 13. Jaenisch, R. et al. (2002) Proc. Natl. Acad. Sci. USA 99, 12889 - 12894. |Article| 14. Committee on Science, Engineering, and Public Policy, Board on Life Sciences (2002) Scientific and Medical Aspects of Human Reproductive Cloning (National Academies Press, Washington, DC). |Report| 15. Dennis, C. (2002) Nature 419, 334 - 336. |Article| 16. Gurdon, J.B., & Byrne, J.A. (2003) Proc. Natl. Acad. Sci. USA 100, 8048-8052. |Article| 3.5 Parenté et diversité des organismes La parenté des organismes résulte de la reproduction. La diversité repose sur une idée qui est celle de l'évolution: les êtres vivants se sont transformés au cours des temps géologiques. |
|||||||||||||||||||||||||||
|
Voir fiche : Qu'est-ce que l'évolution ? |
3.5.1 - Le concept d'évolution affirme que les espèces se sont transformées au cours des temps géologiques |
|||||||||||||||||||||||||||
|
voir fiche Qu'est-ce qu'une espèce ? |
Les mécanismes de l'évolution sont développées au sein de différentes théories de l'évolution. |
|||||||||||||||||||||||||||
|
voir fiche nommer-classer |
|
|||||||||||||||||||||||||||
|
|
Cette classification doit pouvoir rendre compte de l'évolution des espèces au cours des temps géologiques. Les espèces dérivant probablement les unes des autres (la phylogénie, mot créé dans le sens actuel par Haeckel, vient du latin phylum= la lignée, ou du grec phulon = la race; et génie, genèse = la génération ou de genus = le genre), les mécanismes évolutifs doivent s'appuyer sur des facteurs de diversité à partir de points communs à l'ensemble du vivant. |
|||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||
|
Un exemple: |
diblastiques: |
Spongiaires
(éponges), |
||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
triblastiques: |
acœlomates: |
Plathelminthes (vers plats)... |
||||||||||||||||||||||||||
|
pseudocœlomates: |
Rotifères, |
|||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
N.B. dans cette
classification, toujours en usage, les critères
principaux sont d'origine embryonnaire. Ce sont eux qui
déterminent les types cellulaires et les
symétries. La phylogénie est sous-jacente si
l'on prend en compte la complexification et la
diversification des tissus entre la première ligne et
la dernière ligne du tableau. |
|
|
||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
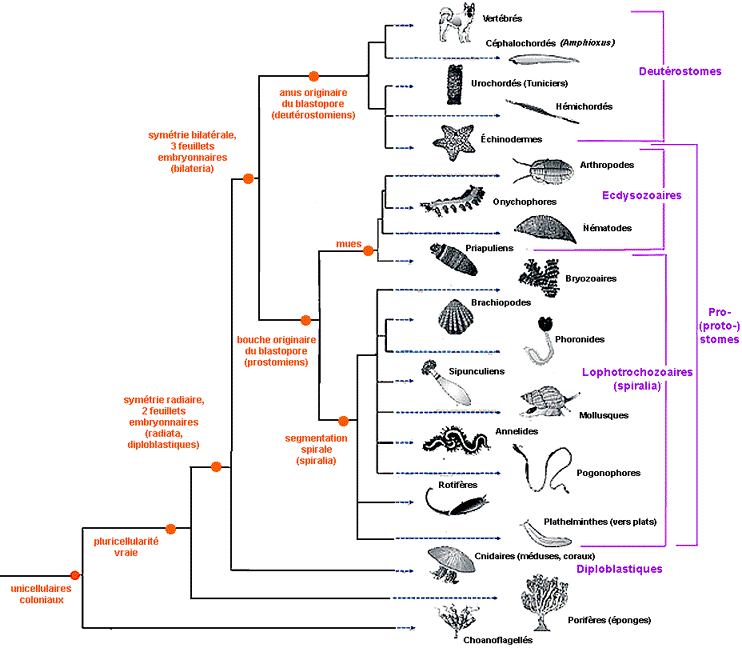
Classification phylogénétique cladiste des animaux (d'après Biologie du développement; S.F. Gilbert, De Boeck, 2004, fig. 2.21 et http://www.americanscientist. org/content/AMSCI/ AMSCI/Image/ FullImage_ 20041123152054_307.jpg ) Pour le texte (en anglais) voir par exemple: The origin of body plan (http://www.americanscientist. org/template/AssetDetail/ assetid/39001/ page/3#39113) |
 |
|||||||||||||||||||||||||||
|
Si l'on reprend les mêmes groupes que dans la classification évolutive ci-dessus on obtient le tableau suivant, qui est aussi mémorisable pour un élève de seconde : |
|
On voit que le point essentiel est la
disparition du critère "cœlomate /
pseudocœlomate / acœlomate" comme
élément classificatoire principal. |
||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||
|
On
peut penser qu'il est encore trop tôt, en seconde,
pour commencer à discuter du sens des
classifications. Et pourtant, en terminale, on s'appuiera
sur des classifications sans avoir pris le temps de les
justifier. Voici quelques repères dans la jungle des
classifications cladistes, à partir d'un article de
référence Table 1 de l'article
nommer, classer : une fiche de seconde le cladisme, approfondissement |
Pour ce qui est des 5
royaumes: Pour les Procaryotes, le Bergey's Manual (l'outil papier de systématique des bactéries à l'usage des microbiologistes) est plus ou moins relayé par le site : http://www.bacterio. cict.fr/ index.html
|
|
Un des articles de référence datant de 2005 et donnant une classification des eucaryotes (en anglais évidemment) ; libre accès en 09/2008 : Sina M. ADL et al. (2005) The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists The Journal of Eukaryotic Microbiology 52 (5) , 399-451 doi: 10.1111/j.1550-7408.2005. 00053.x Comme les classifications cladistes restent relatives (les cladogrammes ne sont pas comparables entre eux) il est nécessaire de souligner que l'idée d'un cladogramme UNIQUE du vivant (et donc d'une seule classification) est très éloignée des fondements du cladisme. Les arbres phylogénétiques cladistes ne sont PAS des illustrations de l'histoire des êtres vivants (voir fiche nommer-classer) et donc pas un "arbre de vie", qui reste un rêve pour un phylogéniste non relativiste mais pas pour un cladiste. Il faudrait davantage voir ces cladogrammes comme des zooms réalisés avec différents appareillages optiques -filtrants certaines longueurs d'onde - et donnant chacun une vue originale, et partielle, des relations de parenté existant entre les êtres vivants actuels ou passés. De plus il faut souligner que l'empressement des phylogénéticiens utilisant l'outil cladistique à contester la validité de tel ou tel phylum de définition ancienne repose la plupart du temps sur des résultats d'études moléculaires (très couramment sur des ADN associés à des ARN ribosomiaux par exemple). Certes, les caractères moléculaires ne sont pas les seuls pris en compte, mais ce sont souvent eux qui sont déterminants pour le choix des groupes (ingroup et outgroup) qui sont à la base de la méthode cladiste. Cette multiplicité et ce relativisme revèlent davantage, à mon sens, l'absence d'une théorie de l'évolution moléculaire (plus ou moins réduite à un mutationnisme aléatoire car la sémiologie de l'ADN n'est pas une question simple, voir le cours de 1èreS), qu'une véritable avancée dans la compréhension de l'évolution, et donc de classifications phylogénétiques pertinentes. Il ne faut pas oublier que la cladistique ne repose sur aucun mécanisme évolutif particulier et se satisferait d'une théorie évolutive moins mécaniciste et moléculariste (voir théories de l'évolution). |
|||||||||||||||||||||||||
|
Si je pense que l'introduction de la systématique phylogénétique (cladistique) dans l'enseignement était nécessaire, je ne pense pas que cela ait été fait avec une prudence suffisante, principalement du fait des implications philosophiques des classifications. En désaccord avec les idées de certains collègues, qui sont diffusées par les sites associés à l'INRP , j'affirme la pertinence des classifications naturelles basées sur des observations et des analogies (et tout particulièrement au collège et dans le primaire) permettant de fournir aux élèves des bases mémorisables. Vouloir déraciner de l'esprit enfantin l'idée d'une classification naturelle est une idéologie inacceptable (voir nommer-classer). D'ailleurs les activités pédagogiques proposées dans ce sens n'ont souvent qu'un rapport lointain avec la méthode cladiste (voir par exemple : http://acces.inrp.fr/evolution/evolution/relations-de-parente/enseigner) par le fait même que les éléments fondamentaux de la méthode: définition des groupes et polarisation des caractères, est TOTALEMENT inaccessible à l'élève : ces exercices tournent à l'acquisition de la pratique d'un logiciel (phylogène) et s'éloignent des objectifs d'observation et d'analyse d'une leçon de science expérimentale. |
||||||||||||||||||||||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}