|
retour cours seconde, accueil, panorama |
|
|
|
révision en cours août 2007 |
||||
|
|
|
plan 1. Une algue unicellulaire (Protiste) à "thalle" siphonné et phase reproductive cœnocytique (croissance végétative, phase reproductive, cycle de vie) 2. Un modèle expérimental réhabilité en ce début de XXIème siècle 2.1 Les premières expériences d'Hämmerling (vers 1930)
le 9/10/2007: Tous les liens vers le site de l'université de Washington (D. Mandoli) ont expiré pour une raison inconnue... j'ai provisoirement intégré les images à mon site grâce au site américain d'archivage (http://www.archive.org/web/web.php). Pour les liensvous pouvez aussi accéder aux pages des D. Mandoli par ce biais. |
||||||
|
1. Une algue unicellulaire (Protiste) à "thalle" siphonné et phase reproductive cœnocytique données principales pour Acetabularia mediterranea |
|
Acetabularia est un genre moderne qui serait apparu vers 38 Ma et dont on connaît environ 8 espèces (discutées). |
||||||
|
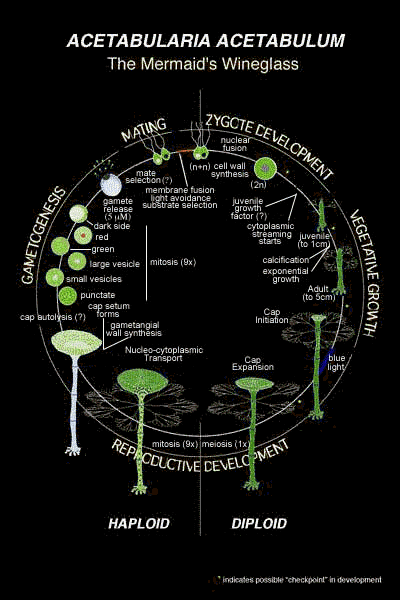
Croissance végétative |
|
|
|
|||||
| |
|
Pour des photos voir la page: |
||||||
|
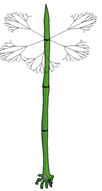
Le "thalle"
végétatif est un axe dressé
extraite de http://
faculty.washington.edu |
|
Chez Acetabularia , où
l'axe dressé peut atteindre 4 à 6 cm de
haut, les rameaux verticillés sont très minces
(et caduques) et plusieurs fois ramifiés (l'algue
ressemble à une petite prêle - Equisetum);
14 à 19 verticilles de rameaux stériles sont
observés lors de la croissance de l'axe. |
|
On considère souvent qu'Acetabularia est une algue siphonnée c'est-à-dire que l'on peut déterminer des territoires cytoplasmiques non cloisonnés qui peuvent être considérés comme autant ce cellules anucléées avant la phase de reproduction...(voir partie 2.3). Le genre Acetabularia fait partie des Dasycladales, regroupées dans les Chlorophycées cœnocytiques (thalles siphonné à plastes chlorophylliens): leur thalle, uninucléé à l'état végétatif, est constitué par un axe dressé (dans un gel calcifié) portant des ramifications verticillées (disposés en verticilles, c'est-à-dire selon un cercle autour de l'axe à chaque niveau de verticille). Des
cœnocytes ? (en anglais) |
||||
| |
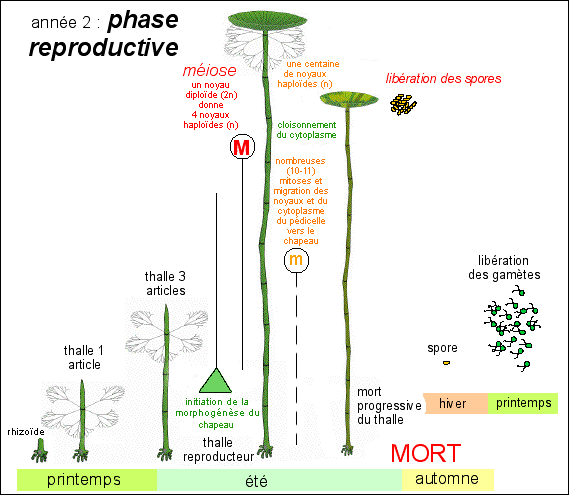
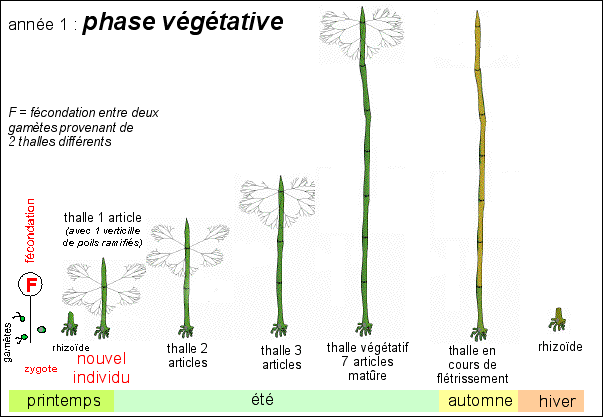
La croissance végétative dure de un à deux ans avant de laisser la place à la phase reproductrice. La fécondation à lieu au printemps lorsque l'eau est chaude (en Méditerranée !). Le zygote croît et donne en été un thalle d'une quinzaine de verticilles. Le thalle végétatif se flétrit à l'automne et seul le rhizoïde persiste en hiver. Un nouveau thalle croît au printemps. Le thalle passe ensuite à la phase reproductrice et forme un chapeau (voir ci-dessous). Habituellement Acetabularia a donc un cycle de vie bisannuel (qui se déroule sur deux ans), la seconde année correspondant à la phase reproductrice. Parfois le cycle est trisannuel. |
|||||||
|
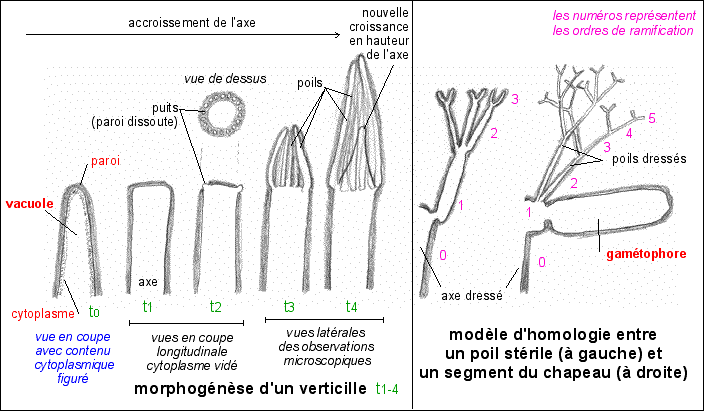
gamétophore
est employé comme synonyme de gamétange Redessiné
d'après Acetabularia: A Unicellular
Model for Understanding Subcellular
Localization and Morphogenesis during
Development, Jacques Dumais, Kyle Serikawa and
Dina F Mandoli, J Plant Growth Regul (2000)
19:253-264, figures 4 et 5 |
||||||||
|
Phase reproductive (du printemps
à l'été |
|
Le "thalle" reproductif comporte un chapeau.  extraite de http://faculty.washington.edu /mandoli/images/ loops-large.gif |
|
Le "thalle" reproductif n'est pas unicellulaire mais c'est un cœnocyte, c'est-à-dire un organisme possédant plusieurs noyaux pour un seul territoire cytoplasmique. Lors de la fructification (initiée par la lumière bleue), il se forme au sommet de l'axe un nouveau verticille de rameaux épaissis (gamétophores) qui constituent, par coalescence latérale, les secteurs rayonnant d'un disque en forme de parasol ou de champignon (d'où le surnom de "Mermaid's Wineglass" - le verre-à-vin des sirènes). Voir ci-dessus (dessins de droite). un modèle de l'origine possible de cette morphologie |
|
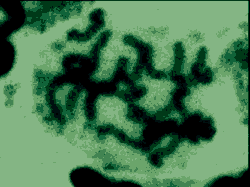
C'est ici qu'intervient la méiose.
Le noyau diploïde augmente de taille
(près de 100µm de diamètre) puis diminue
légèrement. Les chromosomes ne sont visibles que
sous forme de fins filaments, d'abord non accolés puis
accolés sous forme de bivalents (voir montage
ci-contre). |
||
|
|
|
Une question importante est de savoir si la genèse du chapeau commence avant la méiose ou si c'est le contraire. Apparemment c'est bien la genèse du chapeau qui est initiée par la lumière bleue avant que la méiose ne soit lancée. La preuve étant donnée par des expérience de genèse du chapeau sans noyau (voir plus bas). On notera ici que méiose et mitose sont bien des phénomènes nucléaires qui n'impliquent pas ici de cytodiérèse, d'où l'intérêt de considérer séparément la mitose et la séparation des cellules filles lors du phénomène de division (voir cours de seconde et de 1èreS). |
|
Les spores, entourées d'une paroi
cellulaire épaisse pourvue d'une fente circulaire
délimitant un opercule (clapet), sont
libérées par destruction de la plante
mère.
|
||||
 |
|
Une représentation de la vie d'un individu de l'espèce Acetabularia mediterranea comme "bisannuelle" et qui meurt donc après la phase reproductive qui intervient à l'été de la seconde année (la phase végétative peut durer 2 ans et le cycle devient alors trisannuel). |
||||||
|
La croissance est assez lente, environ une semaine pour un article avec un verticille de poils ramifiés. Le passage phase végétative - phase reproductive n'a pas été détaillé ici (voir ci-dessus les schémas de détail; et voir ci-dessous pour les changements de morphologie de l'apex de l'axe). |
|
|
||||||
| |
|
|
|
|
||||
| |
|
|
|
Si A. mediterranea (=acetabulum) ne possède qu'un seul chapeau, d'autres espèces, comme A. crenulata en possèdent plusieurs par allongement du pédicelle qui génère plusieurs verticilles de loges donnant des chapeaux successifs. |
|
Chez A. crenulata les loges forment des rayons qui sont séparés du pied mère par des parois alors que chez A. mediterranea les loges communiquent entre elles et avec le pied mère. |
||
| |
|
|
||||||
|
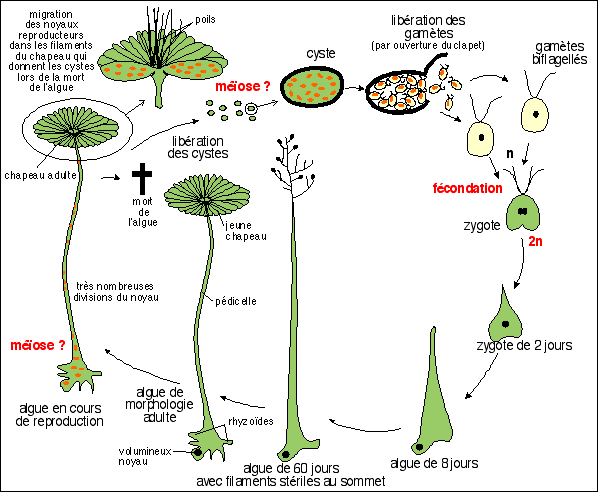
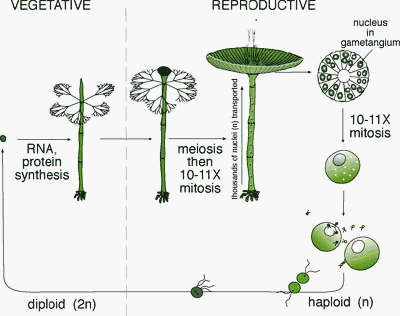
Cycle de vie |
|
Cycle simplifié d'Acetabularia
acetabulum
un cycle plus complexe : |
|
 |
||||
| |
|
|
||||||
|
|
|
Cycle classique présentant des inexactitudes:
|
|
Les poils ramifiés en
verticilles manquent (seuls les poils verticaux ont
été représentés),
voir d'autres représentations. Par exemple l'image de
D. Mandoli (voir article cité plus bas): De plus la méiose, présentée ici comme hypothétique, a été mise en évidence depuis et est bien localisée au niveau de la base de l'axe à la fin de la phase reproductive lorsque le chapeau est déjà formé (De DN, Berger S. 1990. Karyology of Acetabularia mediterranea, Protoplasma 155:19-28; Runft and Mandoli 1996; Shihira Ishikawa 1984). Enfin, les deux types de gamètes (+/- ou mâle/femelle) ne peuvent pas provenir du même cyste, contrairement à ce qui est représenté ici. Par analogie avec les plantes on peut dire qu'Acetabularia est donc un organisme "monoïque" (qui présente les cellules sexuelles de deux sexes dans des "organes" différents). |
||||
|
2. Un modèle expérimental réhabilité en ce début de XXIème siècle |
|
|
||||||
| |
|
Sources: Elaboration of body plan and phase change during development of Acetabularia: How is the complex architecture of a giant unicell built? Dina F. Mandoli,1998, Annual Review of Plant Physiology & Plant Molecular Biology, 49:173-198 |
||||||
|
Pour un bibliographie et des
données récentes (en anglais), |
|
Le laboratoire propose même des kits (fort chers) pour l'enseignement.... cela serait une bonne idée de les développer en France. |
||||||
|
avantages |
|
inconvénients |
||||||
|
|
||||||||
|
* une taille exceptionnelle favorisant les expérimentations (sections, greffes, électrophysiologie, études des facteurs nutritifs...), les observations (gradients...) |
|
|
|
|
||||
|
* cicatrisation extrêmement facile après section |
(cette propriété n'est pas du tout générale, surtout pour une cellule unique) La cicatrisation n'est pas homogène : 50% des cellules blessées au niveau du rhizoïde (non calcifié) meurent. Les jeunes cellules cicatrisent plus rapidement et plus facilement mais une blessure empêche souvent le stade reproducteur. Les cellules en phase de reproduction cicatrisent lentement et difficilement mais atteignent alors une reproduction complète. D'autre part le K+ en solution favorise la cicatrisation - c'est aussi un facteur de coagulation dans le sang humain - (voir par exemple: Wounds incurred in routine cell culture prolong the duration of the life cycle of Acetabularia acetabulum and require K+ to heal). |
|||||||
|
* culture relativement facile depuis les années 2000 |
* difficultés de culture avant les
années 2000, ce qui a provoqué son abandon vers
les années 1980, mais il y a, depuis l'année
2000, un regain d'intérêt. |
|
(voir les travaux de Mandoli notamment (1998); il existe des sociétés qui commercialisent les souches et milieux de culture : c'est par exemple un service de l'université d Austin (Texas) http://www.utex.org/) |
|||||
|
* son génome est petit par rapport à la taille de la cellule : 0,92 pg d'ADN (ce qui est l'équivalent du génome de la tomate et 4,6 fois plus grand que celui d'Arabidopsis thaliana, un autre modèle bien connu des généticiens). Une cellule de plante vasculaire a en moyenne 3,0 pg d'ADN (plantes annuelles) ou 8,5 pg d'ADN (plantes bisannuelles et pérennes). |
* étant donné le nombre très élevé de gamètes (2-6 millions - faire le calcul avec 1 méiose puis 10 mitoses suivies, dans le gamétophore, de 10 autres mitoses: soit 4 X 210 +10 = 4,2 millions - voir cycle en anglais ci-dessus) produits par une seule cellule, on estime à 1,8 à 5,6 µg d'ADN produit dans un cycle par une génération (là aussi, faites le calcul avec 4 millions de gamètes à 0,92 pg d'ADN par gamète - voir ci-contre), ce qui est gigantesque; là encore ce métabolisme est assez particulier à cet organisme et le fait considérer comme primitif. On peut aussi ajouter à ce chiffre la quantité énorme d'ADN synthétisée pour les millions de chloroplastes et de mitochondries. * mais le problème principal reste que cette espèce s'auto-féconde très mal, ce qui rend les études de croisements génétiques plus difficiles (95% des descendants d'une autofécondation ne se développent pas en culture). D.F. Mandoli a réussi à suivre pendant 7 ans sur 10 générations des lignées pratiquement isogénétiques. |
|||||||
|
Des expériences de transformation ont été réalisées (transfert de noyau, injections de génomes étrangers, notamment viraux...) mais les résultats sont encore partiels. |
||||||||
|
* la proportion de bases GC n'est pas inhabituelle ni pour le génome nucléaire, ni pour le génome mitochondrial, ni pour le génome chloroplastique (probablement aux alentours de 44% pour le noyau), ce qui permet d'utiliser les techniques habituelles de clonage des gènes. |
|
|
|
|
||||
|
Des expériences de croisement avec recherche de "mutants" présentant des morphologies "anormales" ont été menées. Mais peu de résultats ont pu être interprétés dans le cadre d'une hérédité chromosomique mendélienne. |
||||||||
| |
|
* le nombre de chromosomes n'est pas encore connu avec certitude; le chiffre le plus couramment accepté est de 40 chromosomes, notamment suite aux travaux de De et Berger (De DN, Berger S. 1990. Karyology of Acetabularia mediterranea, Protoplasma 155:19-28) qui ont observé la méiose avec la formation de 20 bivalents. les chromosomes sont très fins et difficilement observables. |
||||||
|
* le très grand noyau (50-120 µm pour le noyau diploïde), situé dans une cellule relativement transparente, se prête à des observations microscopiques lors de phases de division |
* 3800 copies de gènes d'ARNr ont été trouvées (400 serait un chiffre plus habituel pour une algue verte) ainsi que 32 nucléoles dans le noyau géant, ce qui laisse présager un métabolisme inhabituel des acides nucléiques (en relation avec la phase cœnocytique reproductive). |
|||||||
| |
|
|
* le code génétique d'A. acetabulum n'est pas standard (code n°6 des macronucléus des ciliés et des Dasycladacées dont font partie les acétabulaires) : si le codon UGA est bien un codon stop les deux autres codons-stop standard codent pour la glutamine. Cette différence rend difficile les manipulations génétiques habituelles et nécessite l'usage d'une souche spéciale bactérienne (page sur le code génétique) |
|||||
| |
|
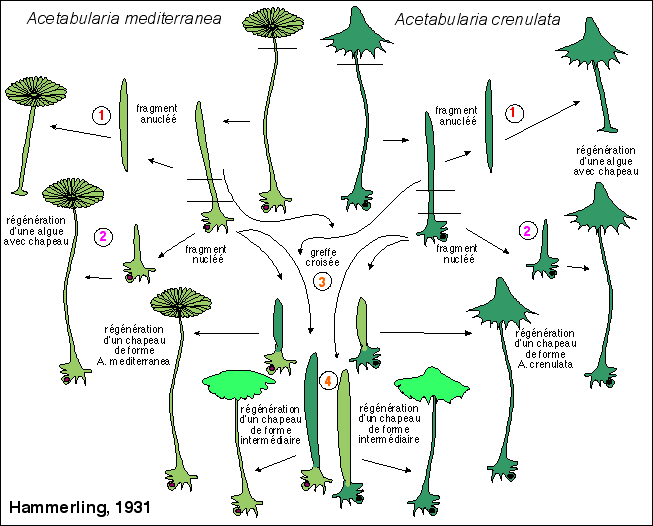
2.1 Les premières expériences d'Hämmerling (vers 1930) Paradoxalement les expériences relatées ici ont permis de découvrir l'importance du noyau dans la morphogenèse - le rôle de cet organite étant totalement inconnu à l'époque des premières expériences - et de postuler la présence de facteurs morphogénétiques issus du noyau (ou en relation étroite avec lui). Ces facteurs seront identifiés bien plus tard comme étant les ARNm. Enfin ces ARNm deviendront finalement des ARN au sens large (étant donné les nombreuses variétés d'acides ribonucléiques). |
||||||
|
Dans certaines conditions des fragments
anucléés (1)
et nucléés (2)
peuvent régénérer (mais pas toujours...)
une algue complète avec chapeau reproducteur (mais sans
noyau et donc vouée à la mort pour le fragment
anucléé...bien sûr). Si un fragment anucléé est
greffé sur un fragment nucléé, et
lorsqu'une algue complète est alors
régénérée, le nouveau chapeau
présente les caractéristiques morphologiques et
structurales de l'espèce qui a fourni le fragment
nucléé
(3). Mais, selon la taille du fragment anucléé (et selon sa position par rapport au chapeau et au noyau - l'apex du pédicelle étant la zone la plus déterminante pour un type de chapeau donné), le chapeau régénéré présente des caractéristiques plus ou moins intermédiaires (4) entre les caractéristiques des algues qui ont fourni l'un ou l'autre des fragments (on parle de "degré de pénétration"). |
|
|
||||||
| |
|
Les interactions entre deux fragments nucléés peuvent aussi être mis en évidence par des expériences complémentaires de greffes de fragments nucléés dont voici un exemple: |
|
|
||||
| |
|
N'ayant pour l'instant pas accès aux publications originales, il me paraît très difficile de faire une relation et une interprétation à partir de connaissances modernes sur l'information génétique. Nucleo-Cytoplasmic Interactions in Acetabularia and other Cells, J Haemmerling, Annual Review of Plant Physiology, June 1963, Vol. 14, Pages 65-92 (doi: 10.1146/annurev.pp.14.060163.000433) |
||||||
| |
|
2.2 Autres données historiques Les chercheurs sont nombreux à avoir travaillé sur le modèle Acetabularia. Je placerai ici les données historiques qui pourraient être utilisées dans des cours. |
|
|
||||
|
(en anglais) Jean Brachet and his school, Henri Alexandre, Int. J. Dev. Biol., 36, 29-41 (1992) |
|
|
|
<< De la
paternité des concepts nouveaux qui flottent dans
l'air du temps L'ADN chloroplastique
>> Remarque: chez Acetabularia, on s'est aperçu que près de 50% des plastes ne possédaient pas d'ADN. Cette particularité est maintenant expliquée par une hérédité plastidiale monoparentale, résultant d'une incompatibilité entre le génome nucléaire du zygote et celui de l'une des lignées plastidiales. Cette incompatibilité entraînant la dégénérescence du nucléoïde des plastes issus de l'un des gamètes (Biologie végétale, t. 1, D. Robert et J.C. Roland, Doin, 1989, p 180). |
|
|
||
| |
|
2.3 L'état des lieux en 1998 d'après D.F. Mandoli |
|
|
||||
| |
|
|||||||
| |
|
|
||||||
| |
|
|
|
|||||
| |
|
La partie suivante comporte ce que l'on peut qualifier d'œillères : une morphologie est reliée à une mutation et à un gène (c'est toujours le problème de la méthode sélective), mais si l'approche reste conventionnelle elle n'est est pas moins un point de départ. Pour témoin la dernière phrase de l'extrait. |
||||||
|
références numérotées de l'article
Pour la comparaison avec la Drosophile, voir les gènes de polarité de l'œuf dans l'ancienne page sur le développement |
|
|
|
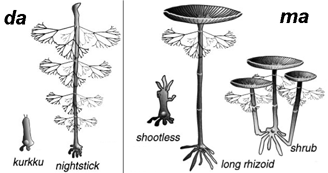 (Figures 6 et 7 de l'article, simplifiées, montrant les principales morphologies classées dans les phénotypes da (développement arrêté) et ma (morphologie arrêtée). |
||||
| |
|
|
||||||
| |
|
2.4 La compréhension classique de la morphogénèse à l'aide de la biologie moléculaire reste une question ouverte J'ai choisi de présenter quelques résultats sous la forme d'une traduction d'un paragraphe d'un article publié en 2000. La complexité inventée dans le cadre d'un vision génético-moléculaire rend la présentation synthétique des résultats quasiment impossible (je renvoie au site de D.F. Mandoli pour des données plus récentes, toutes en anglais...). Il va sans dire que je préfère l'approche du chapitre suivant. |
|
Le laboratoire de D. F. Mandoli s'est lancé dans la course à la biologie moléculaire mais on peut quand même trouver sur le site quelques documents avec des données plus générales ayant davantage de sens. |
||||
|
La localisation de déterminants : la clef de la compréhension de la morphogénèse et du développement d'un unicellulaire ? «Localized Determinants: The Key to Morphogenesis and Development in a Unicell?» extrait de Acetabularia: A Unicellular Model for Understanding Subcellular Localization and Morphogenesis during Development, Jacques Dumais, Kyle Serikawa and Dina F Mandoli, J Plant Growth Regul (2000) 19, p 259-260 |
|
In situ hybridization with poly(U) probes indicates that mRNA in A. acetabulum is present in a gradient with abundance highest in the base near the nucleus (Garcia and Dazy 1986). Messenger RNAs of some genes are localized (Serikawa and Mandoli 1999; Serikawa and others 2001; Vogel 1998), and enzyme (uridine diphosphate glucose pyrophosphorylase (UDPG-P) and others) activities, can be asymmetrically distrib- uted along the stalk (Zetsche and others 1970). » |
|
|
||||
| |
|
2.5 Pistes pédagogiques, théoriques et encouragements à utiliser ce modèle exceptionnel |
|
|
||||
| |
|
|
|
+ de l'histoire des sciences à la biologie contemporaine du XXIème siècle |
||||
| |
|
|
|
Les expériences d'Hämmerling et leurs interprétations habituelles que l'on peut trouver dans les manuels scolaires ont été simplifiées pour cadrer avec la théorie de l'information génétique et du programme de développement. Mais on peut essayer de replacer correctement chaque élément à son niveau. Je vous propose une démarche en 3 étapes : |
||||
| |
|
1. Présentation historique des expériences d'Hämmerling |
||||||
|
En travaux |
|
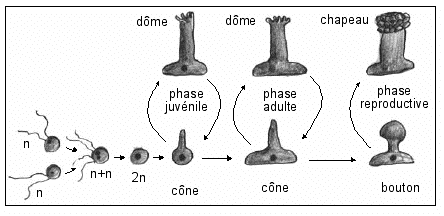
Historiquement les expériences d'Hämmerling suggèrent un rôle du noyau entièrement nouveau en 1936 : le noyau reproductif * comme organite qui contrôle les caractéristiques de la morphogénèse reproductive (et, en élargissant les résultats interprétés dans le cadre de la morphogénèse reproductrice (du chapeau reproducteur), à la mophogénèse complète puis enfin à la morphogénèse d'autres organismes... on dira par la suite que le noyau contrôle la morphogénèse). Son transfert entre espèces voisines, au même état de développement, permet de transférer, plus ou moins partiellement les caractéristiques morphologiques de l'appareil reproducteur, d'une espèce à une autre. * la plupart des dessins illustratifs que l'on trouve dans les manuels sont TROMPEURS puisqu'ils donnent l'impression que l'on transfère des noyaux entre thalles reproducteurs mûrs (à chapeau) alors que les jeunes thalles reproducteurs, chez lesquels le transfert est possible, sont les thalles chez qui la méiose n'est pas initiée,.et donc qui n'ont pas de chapeau (voir cycle ci-dessus et biologie dans la première partie). |
||||||
| |
|
1. Situer les connaissances historiques dans un tableau chronologique . |
|
2. Bien préciser les techniques connues et la nouveauté des réussites techniques. Un transfert de noyau n'est pas une mince prouesse. |
|
3. Analyse OUVERTE de ces expériences et propositions d'expériences complémentaires pour éprouver les hypothèses émises. |
||
|
Un article en anglais très
orienté vers la théorie de l'information
génétique par un des pères des
expériences de clonage: |
|
2. Présentation de quelques résultats modernes de transfert de noyau très médiatisés (expériences de clonage sur la brebis par exemple) |
||||||
| |
3. Comparaisons des techniques, résultats et implications théoriques. |
|||||||
| |
La première différence
essentielle est que le noyau de l'acétabulaire est un
noyau d'un thalle reproducteur dans un état très
particulier qui n'est pas le même que celui d'une
cellule différenciée. |
|
Chez Acetabularia on transfert TOUTE l'information génétique "reproductrice" et totipotente de l'organisme, dans l'autre cas seulement celle d'UNE cellule, adulte et DIFFÉRENCIÉE. Ce deux informations ne sont pas du tout identiques sauf si on les considère comme matériellement et significativement équivalentes (ce qui était le cas dans la théorie de l'information génétique mais on sait maintenant que ces deux informations ne sont pas les mêmes.. d'où la prouesse et la quasi impossibilité de réussir une telle manipulation). C'est pour cela que l'on parle maintenant de pluripotence pour qualifier une cellule capable, malgré une certaine différenciation, de se dédifférencier pour donner différents types cellulaires (in vitro). La théorie de l'information génétique voit ce phénomène comme une reprogrammation du noyau par le cytoplasme. |
|||||
| |
|
Plaçons nous maintenant dans le cadre d'une théorie plus moderne qui considère que les caractéristiques de l'espèce, la vie en un mot, émergent comme des propriétés originales (que l'on présente habituellement sous la forme de 3 fonctions: de nutrition, de relation et de reproduction) à partir des systèmes de l'organismes qui comprennent des éléments matériels (molécules et structures...) mais aussi immatériels (forces, champs...). Dans ce cas il n'y a pas de programme mais une vie émergente à partir du système vivant total qui est décrit complètement, pour un unicellulaire, à l'aide du concept de cellule. |
|
Les expériences d'Hämmerling et certains résultats complémentaires obtenus depuis prennent alors un tout autre sens. Le noyau à l'état reproducteur est prêt à réaliser une méiose suivie de nombreuses mitoses. Son transfert dans une cellule dans le même état physiologique n'empêche pas, dans certains cas, ceux où le transfert de noyau réussit - ce qui n'est pas le cas général -, l'utilisation par le cytoplasme de l'information génétique qui est contenue dans ce même noyau. Les éléments matériels et immatériels supports du développement sont suffisamment similaires pour que l'échange de noyau puisse se faire. Les caractères du développement qui différent entre la cellule originale et la cellule ayant reçu un noyau étranger, reposent donc de façon directe ou indirecte sur la manipulation réalisée. Si la morphologie de l'appareil reproducteur semble partiellement liée au noyau il n'est pas possible de préciser exactement quel est le support matériel ou immatériel de cette orientation du développement. Les ARN se sont révélés bien décevants. Dire que le noyau comporte une "information morphogénétique" laisse comme un goût de mystification étant donné que l'on peut obtenir la même morphologie SANS noyau. |
|
Dans les expériences de clonage on est confronté d'abord au même type de raisonnement puisque certains caractères du donneur du noyau se retrouvent chez l'organisme issu de la manipulation. Mais des éléments viennent s'ajouter : d'abord le développement est TRÈS RARE : la poursuite du développement après transfert de noyau est inférieure à 1%. Ensuite les animaux transformés présentent des ANOMALIES de développement plus ou moins graves. Ces résultats demandent, dans le cadre d'une interprétation ouverte, bien plus que la simple invocation de différences mineures dans l'information génétique. |
||
| |
|
|
|
|
||||
| |
|
|
|
+ des modèles mathématiques et physiques |
||||
| |
|
Jacques Dumais est un des
chercheurs qui a travaillé sur la
morphogénèse chez Acetabularia. |
|
Un article en anglais accessible: Can mechanics control pattern formation in plants? Jacques Dumais, 2007 La page How complex can a cell be ? est plus spécifiquement dédiée à la présentation du mystère de la forme des verticilles de poils d'Acetabularia. Sa page sur la croissance apicale (Mechanic modeling of tip growth) peut tout à fait s'intéger dans un cours de 1èreS (on est loin du contrôle génétique). |
||||
|
En travaux |
|
|
|
|
||||
| |
|
|
|
+ remise en question de l'unicellularité |
||||
| |
C'est non seulement le "thalle" végétatif qui peut être régénéré par un fragment de l'axe mais aussi le "thalle reproducteur"; Ces fragments sont incapables de reproduction sexuée, étant dépourvus de noyau, mais ils peuvent être multipliés. En fait ce type de multiplication végétative serait banal pour un organisme pluricellulaire. Cela pose donc la question de la pertinence de l'unicellularité de l'organisme, basée principalement sur l'observation d'un noyau. |
|
Ce n'est peut-être pas un hasard que le modèle le plus employé en génétique moléculaire dans le royaume animal soit l'embryon de drosophile qui est, dans les premières heures du développement embryonnaire (qui ne dure qu'un jour), un système unicellulaire syncytial (voir ancienne page sur le développement) qui présente des points communs avec Acetabularia. Est-ce une chance ou bien est-ce un obstacle du fait du caractère trop original de ces organismes ?
|
|
Du point de vue morphologique Acetabularia est un "thalle" composé de plusieurs "organes". Le problème principal vient de ce que la membrane est censée délimiter la cellule. Or, du point de vue dynamique, une membrane n'est que le résultat de l'affrontement de deux dynamiques, son absence est donc à expliquer en terme de dynamique. En ce sens la dynamique au sein d'Acetabularia est peut-être bien plus complexe que celle d'un œuf de drosophile, du fait de la morphologie, sauf à considérer que l'ensemble de cette morphologie est prédéterminée. Il est plus simple de penser que la morphologie résulte des dynamiques. Ces intéressantes questions sont abordés par exemple dans l'ouvrage de Vincent Fleury (De l'œuf à l'éternité - Le sens de l'évolution, Vincent Fleury, Flammarion, 2006) mais pour d'autres organismes. Plusieurs travaux sont en cours pour s'efforcer de trouver une organisation subcellulaire: quelques exemples - Calcification and measurements of net proton and oxygen flux reveal subcellular domains in Acetabularia acetabulum, Serikawa, K.A et al., Planta Sept 2000. v. 211 (4), p. 474-483 - Asymmetric subcellular mRNA distribution correlates with carbonic anhydrase activity in Acetabularia acetabulum, Serikawa KA et al., Plant Physiol. 2001 Feb;125(2):900-11) |
|||
| |
|
Je me permet de donner ma méthode pour trouver sur internet les publications scientifiques d'un auteur (sans avoir accès aux bases payantes, ce qui est malheureusement le cas pour les enseignants du secondaire) : il suffit de taper le nom de cet auteur sous Google Scholar; par exemple pour K.A. Serikawa: http://scholar.google.com /scholar?q=%22author%3 AK.%20A.+author%3A Serikawa%22, ce qui permet de savoir que l'on a affaire à un moléculariste. Une fois que l'on a le nom de l'article, on copie l'intégralité de son titre sous Google et l'on cherche à trouver le site de l'auteur (ou un autre) qui permet PARFOIS le téléchargement gratuit de l'article... Évidemment, il faut comprendre l'anglais. Mais je cite un propos qui est attribué à Claude Allègre, alors ministre de l'éducation nationale: l'anglais n'est plus une langue étrangère... je pense aussi que c'est une langue internationale nécessaire à tout scientifique. |
|
|||||
| |
|
|
|
+ questionnement sur la reproduction |
||||
|
reproduction (cours 2de) |
|
Si, dans le monde vivant, la phase de
reproduction est souvent associée à la formation
de gamètes (cellules sexuelles
spécialisées), on peut noter que les organes
reproducteurs (gamétanges, gamétophores,
gonades...) sont souvent formés bien avant. |
|
Il n'est pas pas surprenant d'observer chez Acetabularia
une telle séparation temporelle. |
||||
| |
|
|
|
+ le noyau renferme les chromosomes, organites de la division; pourquoi sont-ils si difficiles à observer ? |
||||
| |
Comme nous nous sommes efforcés de le présenter dès la seconde, la dynamique d'apparition des chromosomes est probablement la clef de la compréhension de la catastrophe de division cellulaire. |
|
Dans cette structure pseudo-unicellulaire qu'est le thalle d'Acetabularia, il n'est pas anodin de noter que justement les chromosomes ont une forme inhabituelle (finesse...) et que les phénomènes de méiose et de mitoses (très nombreuses) sont très localisées dans le temps et l'espace, mais que justement ces phénomènes (voir ci-dessus) n'ont pas lieu au sein du chapeau que l'on peut considérer comme une structure reproductrice. |
|
Il faut aussi rappeler que les divisions des unicellulaires sont souvent atypiques et que nombre d'organismes unicellulaires n'ont toujours pas de méiose connue. Acetabularia n'est probablement pas un modèle simple pour la compréhension de la division cellulaire. |
|||
| |
|
En conclusion Acetabularia est un modèle que l'on sait aisément cultiver, même si son développement est lent. Doué de capacité de régénération, il supporte très bien les amputations et greffes. Son organisation est suffisamment différenciée et ses phases végétatives et reproductives suffisamment distinctes pour qu'on puisse en faire un modèle très intéressant de morphogenèse. Son unicellularité est un avantage pour les études moléculaires qui en sont évidemment simplifiées. Mais c'est aussi un écueil car il est évident que les approximations faites sur le transport des molécules ou l'homogénéité des compartiments cellulaires ne sont plus valables pour un organisme de cette taille, si tant est qu'elles ont été valables pour des cellules de petite taille (voir page sur le cellule). |
||||||