|
retour plan du cours, accueil, panorama |
|
Une fois la notion de cellule et de travail du vivant précisé (voir chapitre précédent), ce chapitre présente l'organisme humain au travail lors d'un effort. Les cellules sont classiquement regroupées en tissus qui sont "des ensembles de cellules concourant à une même fonction". Les organes étant à leur tour composés de tissus, chaque organe, morphologiquement défini, concourant à une ou plusieurs fonctions. Les organes étant eux-mêmes regroupés en appareils. |
|
Les cellules d'un même organe et de
même origine embryonnaire appartiennent à une
population. |
||
|
|
|
|
||||
|
Plan |
|
1. Qui travaille ? : l'organisme et les
organes |
|
|
||
|
|
|
|
||||
|
1. Qui travaille ? : l'organisme et les
organes
|
|
|
||||
|
|
|
Un effort physique est un travail: il demande de l'énergie et est réalisé par des cellules qui sont les travailleurs. Mais les cellules travaillent ensemble. On dit que ce travail est coordonné. On dit aussi que ce travail intègre, c'est-à-dire est la somme de, tous les travaux de chaque cellule individuelle. Comme un effort est un travail complexe il met en jeu non seulement de très nombreuses cellules mais aussi des organes et des appareils différents (en fait, de presque tous les appareils de l'organisme). |
|
Le
travail de
relation le plus visible
est un travail mécanique réalisé
par l'appareil locomoteur qui comprend les
muscles et le système nerveux. Les
muscles qui permettent de soulever un objet ou de marcher ou
courir sont les muscles qui actionnent les bras et les
jambes, ce sont des muscles squelettiques car ils
sont aussi rattachés aux squelette des membres. Ils
sont composées de cellules musculaires
striées (les cellules qui se contractent ou se
relâchent), de cellules conjonctives (les
cellules élastiques qui accrochent les muscles au
squelette et forment notamment les tendons), de cellules
des vaisseaux sanguins qui irriguent le muscle et de
cellule nerveuses des nerfs qui innervent le muscle.
Si l'on y regarde de plus près il est évident
que le travail mécanique met aussi en jeu des
travaux chimiques (nutritifs par exemple) et
électriques (nerveux par exemple). |
|
Le
travail de nutrition est
réalisé par l'appareil digestif qui
absorbe les nutriments qui seront consommés par les
cellules (ses cellules sont les entérocytes), par
l'appareil ventilatoire et l'appareil
circulatoire qui assurent les échanges gazeux
entre le sang et l'air et entre les cellules et le sang
(leurs cellules principales sont les pneumocytes et les
cellules des vaisseaux sanguins respectivement) et enfin par
l'appareil excréteur rénal qui
règle la composition du sang mais aussi de la
peau qui, grâce à sa riche irrigation
sanguine et grâce aux glandes sudoripares, rejette de
l'énergie thermique excédentaire ainsi que
quelques ions (transpiration). Le travail de nutrition n'est
pas uniquement chimique mais aussi mécanique
(la pompe cardiaque par exemple) et même
électrique ("commande" nerveuse de très
nombreux organes).
|
|
|
|
|
||||
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
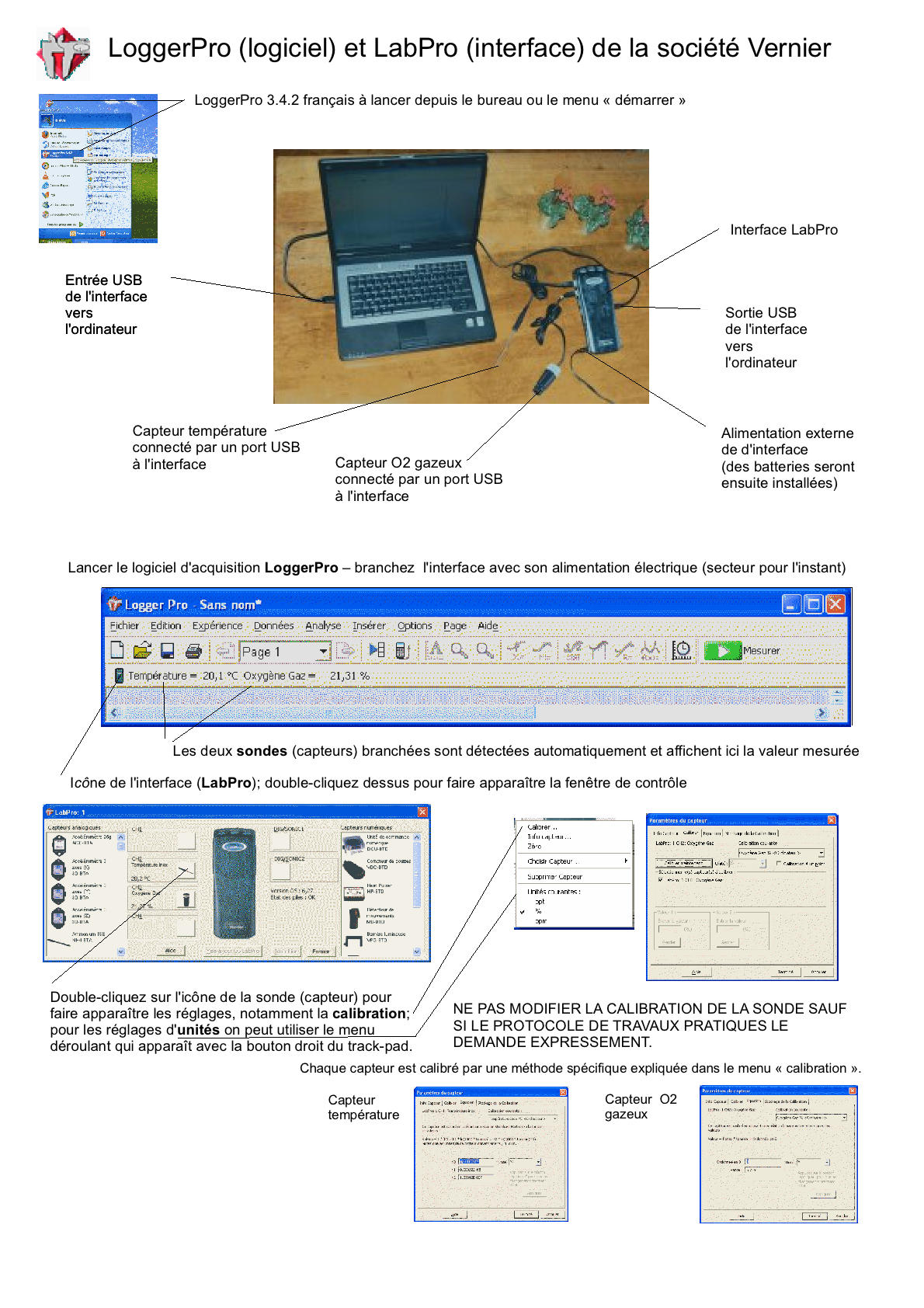
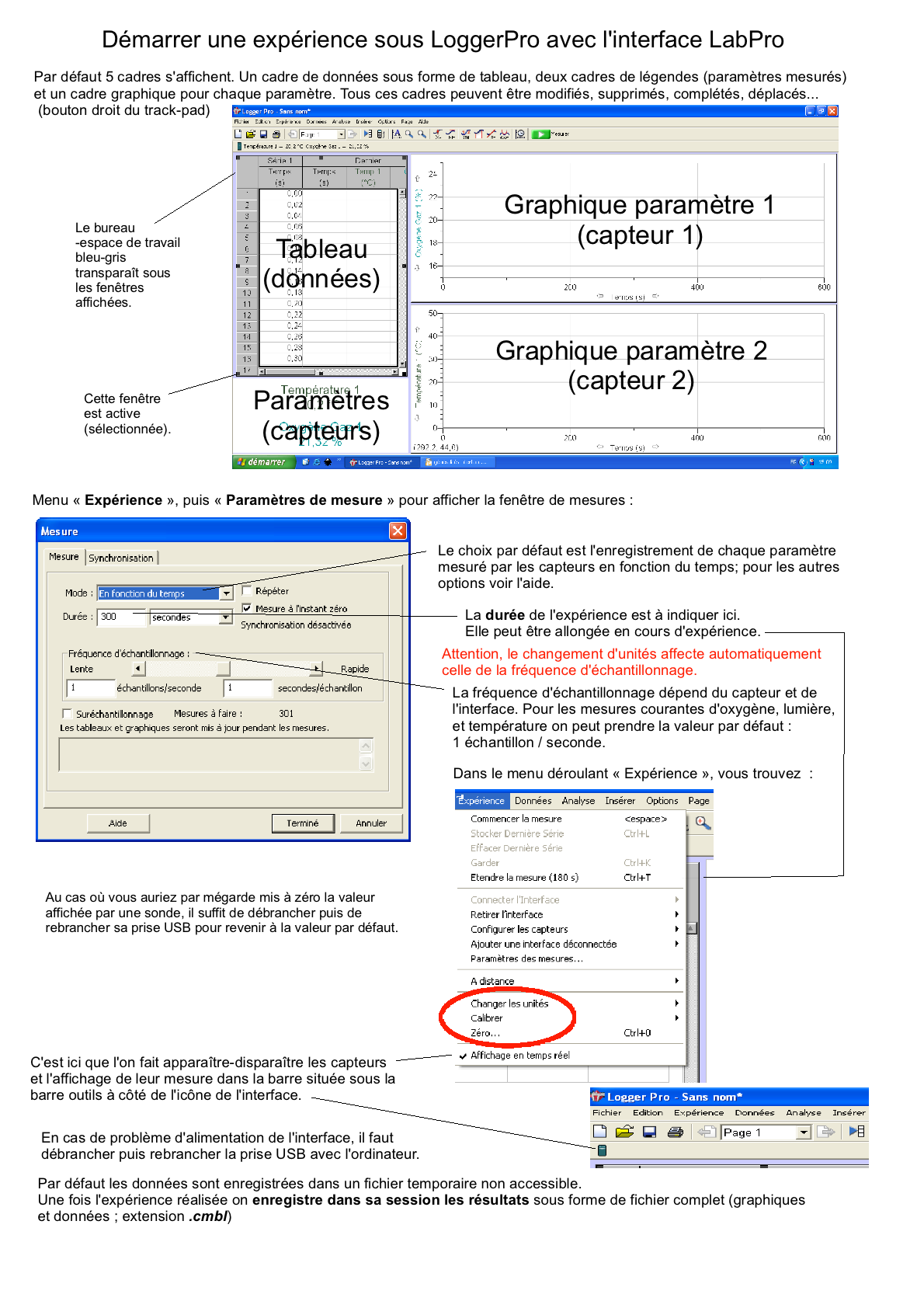
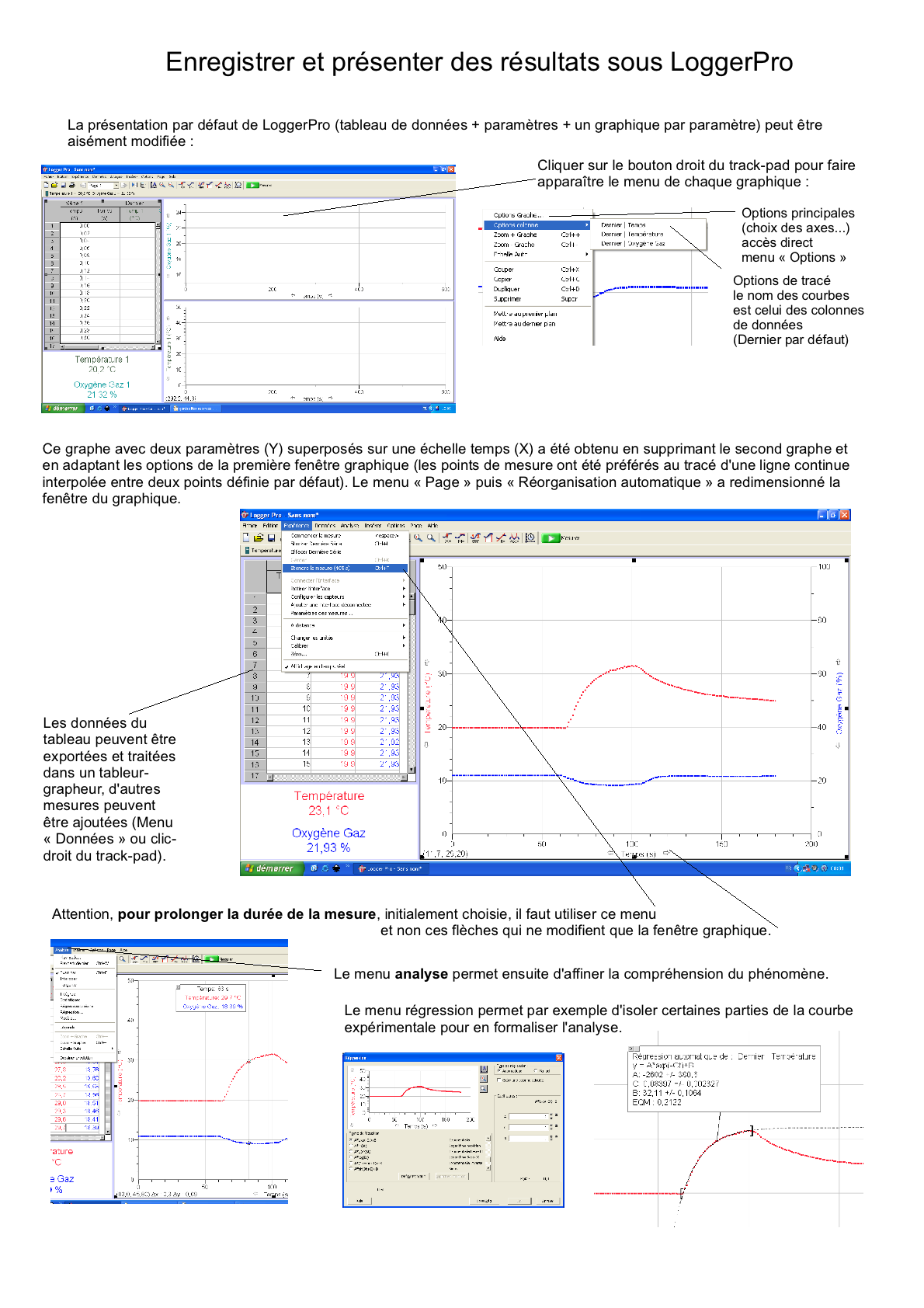
Mon avant-dernier lycée utilise depuis 2007 le logiciel LoggerPro et les capteurs de la firme Vernier (commercialisé en France par Calibration); nous disposons de 14 PC portables (la région a refusé l'équipement Mac) mais je travaille à mon domicile sur Mac Fiches découvertes sur le logiciel et les capteurs: Généralités interface (fichier .odg, pdf, png) Démarrer une expérience (fichier .odg, pdf, png) Présenter des résultats (fichier .odg, pdf, png) Vous pouvez télécharger une version démo EN ANGLAIS (http://perso.orange.fr/ calibration/logger.html) du logiciel pour voir les fiches TP |
|
Le terme de
paramètre semble avoir
été employé pour la première
fois par Leibniz (dans le
développement d'une famille de courbes). C'est aussi
Leibniz qui, dans un manuscrit de 1684, utilise le mot
fonction dans le sens où on le comprend
actuellement (ou presque): « toute quantité
qui varierait d'un point à un autre d'une courbe
» (on lui doit aussi les mots de variable et
constante). (d'après M. Kline, Mathematical
Thought from Ancient to Modern Times, New York, Oxford
University Press, 1972, p. 339-340.; cité dans les
notes de Paraboles et catastrophes, Réné
Thom) |
|
Remarque: |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
2.1.1 Les mouvements ventilatoires et le rythme ventilatoire Un cycle ventilatoire comprend une inspiration (active) et une expiration (passive). Le volume courant est le volume d'air renouvelé dans les poumons à chaque cycle ventilatoire. Lors d'exercices physiques un volume supplémentaire peut être mobilisé (venant du volume résiduel et de l'espace mort). |
|
Remarque: |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
pulmonaire
total
(6 L) |
|
Les volumes pulmonaires de la colonne de gauche sont estimés AU REPOS. Il est essentiel de noter qu'au cours d'un exercice physique l'espace mort diminue pour atteindre moins de 1/20ème du volume courant et devient donc négligeable. AU REPOS, à chaque inspiration, 0,35 L (volume renouvelé) d'air "frais" se mélangent aux 1,65 L (1,5 L de réserve expiratoire + 0,15 L de l'espace mort) et donc il n'y a un renouvellement que du cinquième de l'air pulmonaire. Lors d'un exercice le renouvellement atteint près de 0,5 L sur 1,5 L soit 1/3. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fiche TP (nécessite LoggerPro) volumes ventilatoires (vide).cmbl |
|
Le rythme ventilatoire, ou plutôt la fréquence ventilatoire, nombre de cycles ventilatoires par unité de temps, augmente bien sûr avec l'activité physique. (Rythme est un mot très général pour caractériser des phénomènes périodiques ou même des mouvements temporels très divers). Le paramètre le plus utilisé pour comparer la ventilation chez différents organismes est le volume de ventilation qui est égal au produit de la fréquence ventilatoire avec le volume courant. Le débit ventilatoire (DV ou ventilation (noté V° voir ci-contre-->), exprimé en L.min-1 ou L/min) est égal au volume courant (VC) multiplié par le fréquence ventilatoire (FV) : |
|
Exercices et étude de documents (Belin p 121 et p160-163, Bordas p 154-155)
Le
débit
est représenté ici
par V° mais il devrait être noté avec un V
majuscule surmonté d'un point : |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
La fréquence ventilatoire est sous
le contrôle d'un centre bulbaire respiratoire
relié par des nerfs aux principaux muscles de
l'inspiration: les muscles intercostaux et le diaphragme. La
seule phase active ventilatoire involontaire est
l'inspiration. C'est un réflexe (contraction
involontaire sous l'effet d'une stimulation) mais il peut
être contrôlé par les centres
supérieurs (cerveau). Lors
d'une expiration active volontaire les muscles abdominaux
sont mis à contribution en refoulant les
viscères et donc le diaphragme vers le haut.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
En 2005 le "Prix du Ministère" de La Recherche a été attribué à Bernard Sapoval, Maddalena Felici , Marcel Filoche , Benjamin Mauroy et Ewald Rudolf Weibel pour leurs travaux sur la "Physique de la respiration chez les mammifères : efficacité ou robustesse" (http://www.larecherche.fr/ html/ LPLR/ laureats/ 2005.jsp# ministere: je conseille de lire la présentation faite lors de la remise du prix: http://www.larecherche.fr /html /2008/pdf/ sapoval.pdf) On peut aussi voir la conférence de Bernard Sapoval (Laboratoire de physique de la matière condensée, Ecole Polytechnique: http://pmc.polytechnique.fr/ groupes/irregularite/activites/ hydropoumon/ hydropoumon.htm ) : Physique de la respiration accessible sur le site de l'ENS, diffusion des savoirs (http://www.diffusion.ens.fr/ index.php?res=conf&idconf=230), et d'autre part, pour une approche plus biologique orientée vers le développement, je conseille les travaux de Vincent Fleury et notamment son article dans Génétiquement indéterminé, 2007, Quæ (Morphogenèse des structures arborisées et conditions physico-chimiques d'une croissance biologique auto-organisée, pp 111-140). |
|
Propos de biophysiciens
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
2.1.2 La consommation en dioxygène et
V°O2max
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fiche TP (nécessite LoggerPro) consommation dioxygène dioxyde carbone effort (vide).cmbl |
|
On mesure la concentration en
dioxygène et en dioxyde de carbone de l'air
expiré (en % et en ppm)
et en en déduit la
consommation cumulée de
dioxygène et le rejet cumulé
de dioxyde de carbone (en
L.h-1 ou
mL.mn-1).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
L'intensité respiratoire
(IR) est le volume de dioxygène
consommé par unité de temps et de masse (en
L.min-1kg-1). |
|
Après un effort l'organisme continue de consommer plus de dioxygène qu'au repos. Cette surconsommation est appelée "dette en oxygène" et correspond probablement à l'inertie du système métabolique et à la reconstitution des réserves énergétiques de l'organisme. |
|
V°O2max
est un excellent indicateur de la capacité
d'effort d'un sportif
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
(Belin p 121, Bordas p 122). |
|
2.1.3 Le rythme cardiaque
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Le cœur isolé de ses nerfs bat à un rythme autonome élevé (± 89 battements.min-1). Le système parasympathique (PS) diminue en permanence le rythme cardiaque (c'est un tonus cardiomodérateur) plus intensément que ne l'accélère le système orthosympathique (OS) (tonus cardioaccélérateur). (Belin, tableau 4 p 158; rythme autonome (89 battements.min-1); rythme avec l'OS et le PS (49 battements.min-1); rythme avec le PS seul = tonus cardiomodérateur (44 battements.min-1); rythme avec l'OS seul = tonus cardioaccélérateur (84 battements.min-1).) |
|
Exercices et
études de documents: la révolution
cardiaque (Belin p 140, Bordas p 126) et son contrôle
nerveux (Belin p158 -159, Bordas p 152-153; et exercice
n° 1 p 167)
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
Remarques: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Le terme pression artérielle désigne couramment la pression moyenne (au-delà de la pression atmosphérique) qui règne dans les grosses artères et MESURÉE à l'aide d'un sphygmomanomètre (ou brassard gonflable associé à un manomètre et un stéthoscope). On mesure bien évidemment soit la pression au bras en position couchée, soit la pression au bras en position debout ou assise, à hauteur du cœur, de façon à s'affranchir de l'énergie gravitaire. |
|
On donne deux valeurs: la valeur la plus
élevée est la pression systolique
(Pas) et la pression la moins
élevée: la pression diastolique
(Pad). On notera que cette valeur n'est pas la moyenne arithmétique des deux valeurs extrêmes (120/80 mmd'Hg = 93 mmd'Hg et non 100 mm d'Hg) mais qu'elle tient compte de la durée de la diastole qui est supérieure à celle de la systole.
|
|
On parle d'hypertension lorsque la pression artérielle se situe ne permanence au-dessus* des valeurs normales.
norme de la pression artérielle chez les sujets de plus de 18 ans (tableau 20.1, Wilmore et Costill, 2006, Physiologie du sport et de l'exercice, De Boeck Université) * il n'y a pas de maladie de l'hypotension (valeur située en permanence en dessous des valeurs normales): l'hypotension n'est pas un état stable à la différence de l'hypertension. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
2.1.5 La nutrition et la demande en glucose
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
La teneur en glucose du sang (glycémie) est un paramètre sanguin et donc interne en fait, même si les apports alimentaires peuvent intervenir pendant l'effort. Les muscles squelettiques en activité comme le cœur ou le système nerveux consomment du glucose en grande quantité. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
2.2.1 les paramètres cardiaques |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Les paramètres cardiaques sont: |
|
|
|
Exercice
d'application: |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
2.2.2 les paramètres pulmonaires |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
En plus de la fréquence ventilatoire externe, il faut ajouter trois paramètres pulmonaires internes : |
|
|
|
Étude de documents: (Belin p 142-143 et p 147, Bordas p 128-129 et 130-131)
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
Étude de documents: (Belin p 147, Bordas p 123) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Physiologie du sport et de l'exercice, Wilmore et Costill, De Boeck, 2006, p 222s |
|
graphique 1
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Physiologie animale, Adaptation et milieux de vie, Schmidt-Nielsen, Dunod, 1998, pp33s
page spéciale sur les carbonates sur le site associé |
|
Il est très facile de mettre en
évidence la sensibilité des Mammifères
au CO2 de l'air pulmonaire en ajoutant du
CO2 dans l'air inspiré. Pour une
concentration de 2,5% de CO2
(par rapport au 0,04%
habituels) la ventilation est presque
doublée. A plus fortes concentrations le
CO2 a un effet narcotique. Le dioxygène a
également un effet sur la ventilation mais il est
beaucoup plus faible (pour une variation de 21% à
18,5%, l'effet est quasiment indécelable). |
|
On pense donc que le centre ventilatoire
bulbaire est directement sensible à la
pCO2 du sang (et du liquide
céphalo-rachidien) et non à la
pO2. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
On explique ainsi les inspirations profondes que les nageurs en apnée pratiquent avant la plongée, non pas par une charge supplémentaire de dioxygène dans le sang, mais par une élimination supérieure du CO2 sanguin, ce qui a pour effet de diminuer le réflexe ventilatoire (dont le centre est au niveau du bulbe) et augmenter ainsi la durée de l'apnée. En effet la pCO2 reste alors suffisamment faible pendant la plongée pour ne pas déclencher le réflexe d'inspiration. Il va sans dire que cette pratique est fort dangereuse, car une trop forte baisse de la pO2 sanguine peut provoquer une perte de connaissance conduisant ainsi à la noyade. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
Vasculaire vient du latin vascula = petit vase et désigne tout ce qui a trait aux vaisseaux, aussi bien sanguins (veineux et artériels) que lymphatiques. On utilise aussi le préfixe "vaso" pour désigner les vaisseaux. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
La résistance
périphérique est la force qui s'oppose
à l'écoulement du sang dans le réseau
artériolaire et capillaire. Elle dépend de la
constriction des artérioles et de la fermeture des
sphincters (muscles circulaires pouvant fermer un conduit en
se contractant). La vasoconstriction ou la vasodilatation sont sous contrôle nerveux et hormonal (par des substances chimiques transportées par le sang). Une hormone est une substance chimique sécrétée par des cellules spécialisées (dites endocrines), transportée par le sang, et qui agit sur des cellules cibles particulières (pourvues de récepteurs à cette hormone). |
 Structure transversale simplifiée des principaux vaisseaux |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
(Belin p 145 et 147; Bordas exercice n°2 p 143; sphincters à ajouter au schéma p 115). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
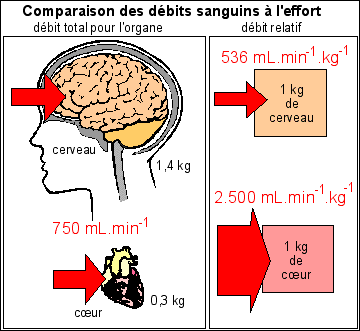
Le débit sanguin au niveau de chaque organe (débit total du sang perfusant l'organe) reflète la distribution de la masse sanguine. (Belin p 144). Pour pouvoir être comparés utilement entre organes, les débits doivent être corrigés par les débits relatifs (rapportés à la masse de l'organe). |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Explication de la notion de débit relatif (à la masse) |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Un
débit
est un volume (ou une masse) de fluide qui passe à
travers un organe par unité de temps. Il
s'écrit donc en mL.min-1 par exemple. Pour pouvoir comparer l'irrigation de chaque cellule appartenant à deux organes différents, on divise par la masse de l'organe et l'on compare donc l'irrigation de deux masses identiques d'organes. |
|
 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Ici les
débits
relatifs sont donnés en
mL.kg-1.min-1, ce qui veut dire que
l'on compare les débits d'1 kg de chaque
organe: le cerveau pesant 1,4 kg, son débit
relatif est de 750/1,4 mL.kg-1.min-1 =
536 mL.kg-1.min-1; ce qui revient
à dire qu' 1kg de cerveau reçoit un
débite de 536 mL.min-1. On peut faire de
même avec le cœur qui reçoit un
débit de 750 mL.kg-1.min-1
à l'effort, mais comme sa masse est de 0,3 kg (300g),
son débit relatif, c'est-à-dire correspondant
à 1 kg de cœur, est de 750/0,3
mL.kg-1.min-1 = 2.500
mL.kg-1.min-1; ce qui revient à
dire que 1 kg de cœur reçoit un débit de
2500 mL.min-1. On peut alors comparer les
débits relatifs entre 1 kg de cerveau et 1 kg de
cœur à l'effort et l'on trouve une
différence de 536 à 2500 mL.min-1,
ce qui est bien plus parlant physiologiquement et surtout
bien plus éclairant que l'égalité des
débits à l'effort (750 mL.min-1)
trouvée sur la deuxième ligne du tableau.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Analyse du tableau |
|
Description |
|
Au repos, les organes recevant le
débit le plus faible ne sont pas les reins mais
l'abdomen (29 mL.kg-1.min-1 ) et aussi
les muscles (41 mL.kg-1.min-1). Le
débit maximal passe en fait par les reins (3667
mL.kg-1.min-1); cerveau, cœur et
peau ont des débits importants mais
intermédiaires entre ceux des reins et des
muscles. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Interprétation - Explication |
|
Le débit sanguin est la masse de
sang qui irrigue les cellules d'un organe par unité
de temps. Plus le débit est important plus les
cellules peuvent avoir de nutriments à leur
disposition et évacuer leurs déchets. On
établit donc une relation directe entre
activité et débit sanguin. Le débit
étant directement proportionnel à
l'activité (le travail) d'un organe. Les
variations de débit sont considérées
comme étant commandées par l'organe
lui-même qui adapte la vasoconstriction
artériolaire et la fermeture des sphincters à
son activité métabolique. Le rein représente un organe à part car le sang qui perfuse à travers est filtré, ce qui veut dire que ce sang ne sert pas uniquement et directement à la nutrition de l'organe, comme c'est le cas pour les autres débits. Un débit de 1,1 L.min-1 pour les reins signifie que tout le volume de sang de l'homme (~6 L) passe à travers les reins en moins de 6 min. Cependant tout le sang qui perfuse le rein ne passe pas le filtre proprement dit (en donnant l'urine primitive) car seul le plasma (sang débarrassé de ses cellules) passe à hauteur de 180 L par jour, soit 30 fois le volume sanguin tout de même. La majorité de l'eau est réabsorbée (voir cours général pour des précisions). Le rein est l'organe de filtration du sang qui travaille moins à l'effort qu'au repos. Le débit superficiel au niveau de la peau est élevé même au repos et devient supérieur à celui des muscles lors de l'effort, car la peau effectue un travail spécifique: l'évacuation de l'énergie thermique métabolique (chaleur produite par les réactions chimiques et la contraction musculaire). C'est le réseau artériolaire et capillaire superficiel qui permet l'échange thermique entre le sang et le milieu extérieur. On notera que la transpiration, qui nécessite la consommation d'une énergie de vaporisation (ou chaleur latente de vaporisation), permet aussi grandement de diminuer la température sanguine superficielle (ou pulmonaire) et donc d'évacuer de la chaleur. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Pour des données
classiques |
|
La physique de la géométrie
de l'arborisation artério-veineuse est aussi fort
étudiée, comme celle de l'arbre bronchique.
D'une part la mise en place lors du développement
(croissance dendritique; voir par
exemple les travaux de Vincent Fleury (article dans
Génétiquement
indéterminé, 2007, Quæ
(Morphogenèse des structures arborisées et
conditions physico-chimiques d'une croissance biologique
auto-organisée, pp
111-140)) et d'autre part la
physique de l'écoulement du fluide sanguin dans cet
incroyable réseau (27 ordres de grandeur entre
l'aorte et les plus fins capillaires).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
en travaux 2010 Wilmore et Costill, 2006, Physiologie du sport et de l'exercice, De Boeck Université |
|
La pression artérielle (définition ci-dessus) augmente fortement à l'exercice. Si une valeur de 200 mmd'Hg est courante on observe fréquemment chez des athlètes très entraînés mais non hypertendus des valeurs de 240 à 250 mm d'Hg à l'exercice. |
|
L'augmentation de la Pam (voir sigles ci-dessus) est essentiellement due à celle de la Pas. L'augmentation de la Pas s'explique principalement par l'augmentation du débit cardiaque alors que le Pad change peu à l'effort. Une augmentation de plus de 15 mm d'Hg de la Pad est considérée comme pathologique et constitue un critère obligeant à stopper toute épreuve d'effort à visée diagnostique. |
|
On notera que les exercices sollicitant les membres supérieurs plutôt que les membres inférieurs font nettement plus augmenter la pression artérielle. On pense que les membres supérieurs possèdent une masse musculaire moindre et un volume circulant plus faible et donc une résistance à l'écoulement du sang plus forte que les membres inférieurs. Du fait de l'irrigation cardiaque (artères coronaires) et comme la consommation myocardique (du tissu musculaire du cœur) de dioxygène est proportionnelle au débit sanguin du myocarde (qui varie lui-même de façon directement proportionnelle à la fréquence cardiaque et à la pression artérielle systolique) tout exercice physique réalisé avec les bras augmente fortement la travail du myocarde. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
On peut noter une légère baisse de la Pas lors d'exercices sous-maximaux lorsque l'effort est prolongé. Elle serait due à une vasodilatation des artérioles musculaires diminuant la résistance périphérique totale. |
|
Lors d'exercices de force (haltérophilie ...) on peut atteindre des pressions artérielles de 480/350 mm d'Hg. Les athlètes réalisent alors souvent la manœuvre de Valsalva qui consiste en une expiration volontaire, bouche, nez et glotte fermés. Cette manœuvre a pour effet d'augmenter la pression intrathoracique et donc la pression dans le reste du corps puisque la pression intrathoracique ainsi augmentée s'ajoute à la valeur habituelle de la pression à laquelle sont soumis les vaisseaux au niveau du cœur. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

 graphique
2
graphique
2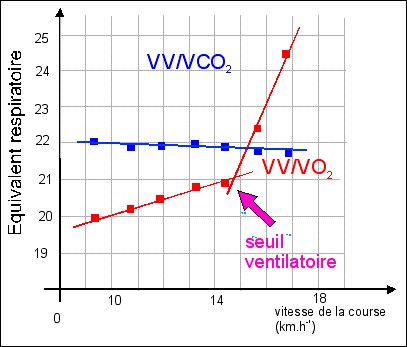
|
|
|
|
|
3.1 Bref historique: de l'analogie
mécanique à la
systémique
|
||||||||||||||||||||||||||||
|
On qualifie de mécanisme ou de mécanicisme une pensée philosophique qui explique la Nature par les lois des mouvements des corps matériels. |
|
Cette philosophie a eu un
développement très important au
XVIIème siécle mais de nos jours on
peut encore dire que le mécanicisme fait partie de
nos modes de pensée intuitifs : On pourrait dire que cette philosophie
s'est diluée dans le matérialisme
athée. « Et
véritablement l'on peut fort bien comparer les nerfs
de la machine que je vous décrits, aux tuyaux des
machines de ces fontaines ; ses muscles et ses tendons,
aux autres divers engins & ressorts qui servent à
les mouvoir ; ses esprits animaux à l'eau qui
les remuë, dont le cœur est la source, & les
concavitez du cerveau sont les regars... » |
|
 |
||||||||||||||||||||||||||||
|
E.U.V12, 2007, articles
"mécanisme" et "cybernétique")
|
|
|
||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
 Un réseau d'expression génétique au cours du développemnt... |
|
La cybernétique
(du grec "kubernêsis" =
gouverner) ou science des
échanges d'information a été, depuis le
milieu jusqu'à la fin du XXème
siècle, une clef de compréhension du
fonctionnement de l'organisme. La
biocybernétique s'est intéressée
à la logique des systèmes biologiques en se
focalisant d'une part sur la recherche d'un programme
(cet aspect a été absorbé par
la biologie moléculaire avec sa notion de
programme génétique) et d'autre part
sur la notion d'automate abstrait (plus ou
moins intégrée au sein d'une biologie
des systèmes ou systémique).
|
|
On considère Norbert Wiener , auteur de Cybernetics or Control and Communication in the Animal and the Machine (1948) comme le père de la cybernétique. Dans son ouvrage Wiener fait la synthèse de toute une série de recherches poursuivies dans le domaine des mathématiques pures (théorie de la prédiction statistique), dans le domaine de la technologie (machines à calculer, télécommunications), dans ceux de la biologie et de la psychologie, et jette les bases d'une science nouvelle, à support mathématique, destinée à couvrir tous les phénomènes qui, d'une manière ou d'une autre, mettent en jeu des mécanismes de traitement de l'information. |
||||||||||||||||||||||||||||
 Le paysage épigénétique de Waddington (1956) |
|
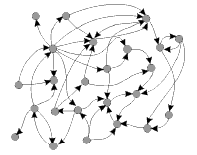 Un réseau génétique stylisé |
|
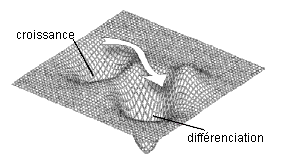 Un paysage avec 2 attracteurs |
||||||||||||||||||||||||||||
|
Waddington est avec René Thom un des fondateurs d'une vision continue des phénomènes du vivant (voir page sur les modèles thomiens) (in D. Ingber, Int. J. Dev. Biol. 50: 255-266 (2006)) |
|
Les deux illustrations
présentées ci-dessus permettent de saisir
intuitivement la différence entre les
modèles discontinus (à partir de
réseaux) et les modèles continus
(à partir de fonctions, d'attracteurs et de
champs). |
||||||||||||||||||||||||||||||
|
|
|
|
|
|||||||||||||||||||||||||||||
|
|
|
3.2.1 L'effort met en œuvre des fonctions en vue
d'un travail
|
|
|
||||||||||||||||||||||||||||
|
|
|
En physiologie, comme en physique, un effort c'est la mise en œuvre de forces en vue d'un travail.. En langage courant l'effort peut mobiliser des forces comme pour un travail intellectuel. Les forces, en biologie sont des fonctions. |
|
Géométriser les fonctions vitales c'est comprendre la vie. Même si cette compréhension repose sur des approximations inhérentes à tout modèle comme l'analyticité des fonctions considérées. C'est un bel horizon qui s'ouvre à la biologie théorique. (Pour des collègues qui liraient ces pages, je renvoie à une page en construction sur la théorie des modèles de René Thom). |
||||||||||||||||||||||||||||
|
|
|
3.2.2 Le premier principe de dynamique du vivant ou
principe
d'homéostasie:
les fonctions du vivant sont
stables (résistantes à de petites
pertubations)
|
|
|
||||||||||||||||||||||||||||
|
|
|
En physiologie on a l'habitude de considérer que l'organisme est en équilibre dynamique : on dit qu'il fonctionne, ou encore qu'il y a homéostase. Ce terme vient du grec homéo (ou homo = identique) et stase qui signifie position (voir cours de 1èreS). La propriété d'un système en homéostase est l'homéostasie. Un système homéostatique est un système qui est stable face à de petites perturbations. |
|
Il y a une incontestable
stabilité de l'être vivant qui maintient
sa forme et se déplace par exemple en consommant de
l'énergie qu'il doit trouver dans ses aliments. Du
point de vue physiologique, c'est-à-dire du point de
vue du fonctionnement, ce qui est stable c'est la
fonction. L'organisme maintient une stabilité de
la fonction. |
|
Par analogie avec le
premier principe de la physique (formulé comme
l'équivalence chaleur-travail ou encore le principe
de conservation de l'énergie - cinétique et
potentielle), l'homéostasie est une
propriété du vivant qui peut être
formulée sous la forme d'un principe. Voir
cours
de
1èreS
pour quelques données sur ce principe. |
||||||||||||||||||||||||||
|
|
|
1er principe de dynamique du vivant = principe d'homéostasie « un être vivant est un
système homéostatique ou en homéostase
» |
|
|||||||||||||||||||||||||||||
|
|
|
Par exemple l'approvisionnement en dioxygène d'un muscle doit être maintenu à un niveau qui dépend de l'activité de ce muscle. Comme pour la forme dans l'espace qui est maintenue stable lors du mouvement (nous ne sommes pas des protoplastes mais notre squelette articulé et nos muscles nous permettent à la fois de changer de forme sans changer la forme globale de notre corps - on reconnaît un homme qui court comme homme aussi facilement qu'un homme allongé...), la physiologie métabolique peut être considérée comme une forme qui est maintenue stable dans un espace de grande dimension (qui comprend tous les paramètres physiologiques). On peut dire que la fonction est déployée dans l'espace des régulations. La forme de cette fonction est une géométrie qui reste stable (qui ne change pas malgré l'effort): ce que l'on qualifie d'homéostasie. |
|
Pour l'effort, l'espace de régulation comprend, outre le temps, des paramètres comme le type d'exercice, le sexe, la taille, l'âge, l'apport alimentaire, la capacité vitale pulmonaire, le volume d'éjection systolique, la fréquence cardiaque au repos, le V°O2 max et tant d'autres. Certains de ces paramètres sont quantitatifs (âge, intensité de l'exercice) mais d'autres sont qualitatifs (comme le sexe ou le type d'exercice...) et ont donc une variation discontinue. Il est clair qu'on ne sait pas actuellement représenter une telle forme. Mais dans chaque situation physiologique il serait à mon avis très profitable de commencer à utiliser les modèles mathématiques déjà disponibles (Pour modéliser une fonction que l'on suppose inaccessible dans sa complexité, il faut bien sûr faire des approximations. La plus courante, utilisée par la théorie des catastrophes, consiste à considérer toutes les fonctions comme des potentiels que la vie cherche à minimiser (ce que Thom appelle "la convention de Maxwell"). On obtient alors un modèle d'une sorte d'espace vital composé de gradients vitaux représentés par des fonctions - voir page en construction: cette convention d'approximation ne devrait pas faire peur aux physiciens. Les potentiels sont décrits mathématiquement par des conflits d'attracteurs (ce sont des objets mathématiques qui caractérisent des régimes asymptotiques, c'est-à-dire qui tendent vers une certaine stabilité). |
|
|||||||||||||||||||||||||||
|
|
|
Il est important de comprendre que l'organisme ne maintient pas stable des paramètres mais bien des fonctions. Ensuite cette stabilité ne se situe pas dans l'espace-temps euclidien (à 4 dimensions) mais dans un espace beaucoup plus complexe qui dépend de nombreux paramètres (et donc à n dimensions, n>4). |
||||||||||||||||||||||||||||||
|
|
|
3.2.3 Recherche des fonctions stables au cours de l'effort |
|
|
||||||||||||||||||||||||||||
|
|
|
|
||||||||||||||||||||||||||||||
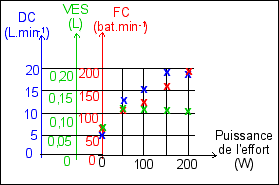
|
exemple 1
* on aimerait savoir sur quel types de
sujets ces mesures ont été faites, avec quel
type d'effort et avec quelle incertitude ces
résultats sont donnés |
|
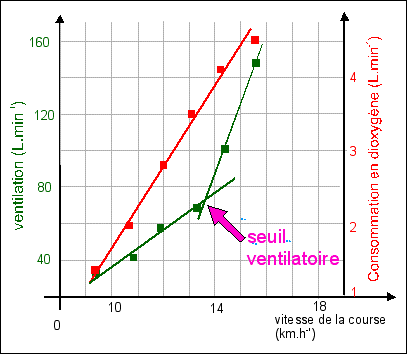
Rien ne permet de dire que la variation de débit cardiaque, de fréquence cardiaque ou de volume d'éjection systolique, est une variation régulière en fonction de l'augmentation de la puissance de l'effort. C'est-à-dire que la pente de la courbe (qu'il ne faut pas tracer) réunissant les points de mesures pour l'une des trois grandeurs mesurées (DC, VES ou FC), n'est pas significative. Elle ne permet que de visualiser des écarts de valeurs entre deux points de mesure. Ce que l'on compare ce sont les valeurs relatives des 3 grandeurs pour chaque puissance d'effort; ce ne sont pas les variations, en considérant de façon fausse que le temps est représenté en abscisses. Entre 0 et 50W par exemple la variation de la fréquence cardiaque n'est pas "moins forte" que celle du débit cardiaque. Ce ne sont pas les mêmes unités et les variations ne sont donc pas comparables, numériquement. Si l'on avait une évolution en fonction du temps on pourrait penser que l'on mesure la vitesse d'un phénomène et donc que les pentes seraient comparables. |
||||||||||||||||||||||||||||||
|
|
|
Le DC est très
nettement augmenté lors d'un effort physique. Il
augmente d'autant plus que l'effort physique est important
mais pas au delà d'un certain seuil, probablement
situé entre 150 et 200W pour ce sujet.
|
|
Le VES est
augmenté lors de l'effort mais d'une valeur qui
semble quasiment indépendante la puissance de
l'effort (elle aurait même tendance à diminuer
par rapport à cette valeur pour l'effort le plus
important).
|
|
La FC augmente avec la
puissance de l'effort.
|
||||||||||||||||||||||||||
|
|
|
Finalement, que nous
apprennent ces graphes ? |
||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
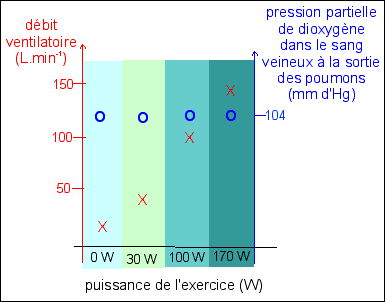
Exemple 2
|
|
|
|
|
||||||||||||||||||||||||||||
|
On ne connaît pas l'âge, le sexe, le type d'éffort, la durée de l'effort, les conditions individuelles physiologiques... d'après Bordas p 131 (ou p 125 de la nouvelle édition, CORRIGÉ) |
|
Description Comme la variation de la puissance de
l'exercice n'est pas régulière, on ne peut pas
vraiment tracer une courbe - dans ce graphique - qui exprime
une relation entre débit ventilatoire et puissance de
l'exercice. Il y a peut-être une fonction mais elle
n'est pas représentable sur ce graphique.
On peut cependant
dire que le débit ventilatoire augmente avec la
puissance de l'effort. |
||||||||||||||||||||||||||||||
|
|
|
Explication Cette fois on est plus près de la fonction qui est bien celle de la prise en charge de dioxygène par le sang au niveau du poumon. Il y a bien une adaptabilité de la ventilation pulmonaire, mais on ne sait pas mettre en relation le débit ventilatoire et la pression partielle de dioxygène au niveau des vaisseaux issus des poumons. Une fois encore la fonction est autre part et on doit se contenter d'une compréhension vague du travail de nutrition. |
||||||||||||||||||||||||||||||
|
|
|
|
||||||||||||||||||||||||||||||
|
|
|
a - Quelques caractériques des fonctions |
|
Les fonctions vitales ne sont jamais
constantes mais stables. La constance c'est la mort.
|
|
Les fonctions sont rarement
accessibles.
|
||||||||||||||||||||||||||
|
|
|
b. Quelques fonctions stables |
|
Lors d'un effort physique les fonctions stables sont reliées au travail de nutrition et de relation: en voici quelques-unes:
|
||||||||||||||||||||||||||||
|
|
|
|
|
|||||||||||||||||||||||||||||
|
|
|
3.3.1 Effort statique et effort dynamique mettent en jeu des fonctions différentes |
|
|
||||||||||||||||||||||||||||
|
|
|
On distingue deux types d'efforts physiques: |
|
* l'effort statique dont l'exemple type est l'exercice d'haltérophilie. Il se fait à respiration bloquée (ce qui est dangereux); il provoque une élévation de la pression artérielle (voir pression artérielle &3a) et une diminution de la pression de retour veineux au cœur. La fonction principale qui est de contracter des muscles ce qui nécessite un travail nerveux et musculaire. Les autres fonctions sont alors inhibées (partiellement évidemment). |
|
* l'effort dynamique dont l'exemple type est la course. Il se fait avec une respiration contrôlée et on observe une adaptation de l'appareil cardio-circulatoire. La fonction principale est de fournir de l'énergie aux muscles (squelettiques) selon leur demande tout en maintenant les apports aux autres organes en fonctionnement comme le cœur mais aussi le cerveau (et les organes des sens). Il est aussi nécessaire d'évacuer le trop plein de chaleur produit par la contraction musculaire. |
||||||||||||||||||||||||||
|
|
|
L'échauffement, ou exercice
préparatoire, en augmentant la température de
fonctionnement des muscles principalement: cœur et
muscles squelettiques, permet d'augmenter le
V°O2 max et la contractibilité des
fibres cardiaques (Hachette, n°7 p 271)
|
||||||||||||||||||||||||||||||
|
|
|
3.3.2 L'entraînement peut modifier durablement les fonctions mais aussi les structures |
|
|
||||||||||||||||||||||||||||
|
|
|
Un exercice physique régulier et progressif (entraînement) est suceptible de modifier la structure des muscles cardiaques et squelettiques (Belin p 172-173). Cette modification, possible chez l'adulte bien après le développement, est qualifiée de plasticité pour les cellules et organes. Dans le cas des muscles on considère qu'il n'y a pas de renouvellement cellulaire c'est-à-dire pas de divisions. On explique donc la plasticité par le remaniement moléculaire permanent des cellules adapté à leur activité. Par exemple la cellule musculaire d'un muscle squelettique fortement sollicité voit aussi bien sa synthèse de protéines contractiles augmenter (ce qui change sa taille) que son équipement enzymatique changer. |
|
Certains paramètres, comme le
V°O2 max ou la débit cardiaque
présentent une nette modification à la suite
de l'entraînement. Nous seulement quand à laure
valeur au repos mais aussi quand à leur variation en
fonction de la puissance de l'effort.
|
||||||||||||||||||||||||||||
|
|
|
* Pour l'appareil ventilatoire il existe
des contraintes physiques (géométriques)
déterminantes. Tous les humains n'ont pas la
même géométrie pulmonaire et donc la
même efficacité (de
toutes petites différences géométriques
conduisent à de grandes différences
physiologiques en terme
d'efficacité). Un
déséquilibre acquis
(par exemple du fait d'une tabagie
précoce - avant 12 ans, âge de la
maturité pulmonaire - empêchant un
développement symétrique et/ou complet de
l'arborescence pulmonaire) peut avoir
pour conséquence un handicap définitif. * Pour l'appareil circulatoire il est possible qu'il existe une plasticité plus grande que pour l'appareil ventilatoire. |
|
"Physique de la respiration chez les mammifères : efficacité ou robustesse" (http://www.larecherche.fr/ html/ LPLR/ laureats/ 2005.jsp# ministere présentation faite par Bernard Sapoval lors de la remise des prix "La Recherche" 2005: http://www.larecherche.fr /html /2008/pdf/ sapoval.pdf) |
||||||||||||||||||||||||||||
|
|
|
|
||||||||||||||||||||||||||||||
{kind=link}
{kind=link}
{kind=link}