|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
retour plan
des thèmes de spécialité,
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Le terme
"diversité" ne pose pas de problèmes mais
celui de "complémentarité" en pose. Ce terme
n'est employé que dans le titre du thème du
programme
et
n'est pas expliqué, ni dans le texte, ni dans les
documents d'accompagnement. Pour moi c'est une idée
fausse. Les métabolismes des êtres vivants ne
sont pas complémentaires. Bien
sûr, au sein d'un organisme pluricellulaire, on peut
observer des complémentarités de
métabolisme entre tissus ou entre cellules afin
d'assurer les fonctions globales du vivant, mais je ne
pense pas que cela soit l'idée que le programme
souhaite nous voir développer. Si c'est une
complémentarité à l'échelle de
l'écosystème, cela me semble plus difficile
à étayer, même si certains auteurs
insistent de plus en plus sur le côté social
de la nutrition). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Plan de cette page: |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
1.1 - On peut
se nourrir de 2 façons: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
Bases de la classe de seconde
à réinvestir : |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dans un écosystème la diversité des êtres vivants au regard de la nutrition se réduit à deux grands types : certains se nourrissent de lumière et d'air (autotrophes) et d'autres se nourrissent d'autres êtres vivants (il est donc préférable de les dire allotrophes plutôt qu'hétérotrophes). |
|
Après avoir longuement hésité, j'utiliserai dans ces pages le terme d'allotrophe. S'il vous chagrine, remplacez-le par hétérotrophe. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Un petit aperçu qui nous permet de revoir les 5 royaumes du vivant (voir cours de seconde). |
|
Remarque: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Vis-à-vis de l'écosystème, un organisme autotrophe est fondamentalement un organisme qui augmente la biomasse: il recherche le soleil et capte le flux lumineux, capte le dioxyde de carbone pour synthétiser sans cesse de la matière organique. C'est un producteur. |
|
Les bactéries (royaume des Procaryotes
ou Monères) et les unicellulaires (royaume des Protistes)
autotrophes vivent habituellement dans l'eau. Ils doivent se
trouver dans une zone éclairée (zone
euphotique des eaux douces et marines) et utilisent le
dioxyde de carbone dissous dans les eaux. Ils composent le
phytoplancton (plancton végétal). |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Vis-à-vis de l'écosystème, un organisme allotrophe est avant tout un consommateur d'autres êtres vivants; qu'il soit herbivore ou carnivore... ou détritivore. Il doit donc capturer sa proie à moins qu'il ne la trouve morte, où qu'il vive dessus. Il possède bien sûr des mécanismes chimiques très voisins de ceux de la plante. Il accroît sa biomasse en convertissant la matière organique consommée (et l'énergie qu'elle contient). Le rendement de conversion dépend beaucoup de l'aliment (rendement faible pour un aliment peu énergétique). C'est un convertisseur ou consommateur. |
|
Les champignons (royaume des Mycètes)
vivent très généralement dans l'air et
sont parasites ou symbiotes. Certains champignons capturent
des vers nématodes avec des lassos (ce sont des
prédateurs). Les animaux (royaume des Animaux)
présentent toutes les relations trophiques avec les
autres êtres vivants: consommateurs
détritivores, prédateurs, parasites et
symbiotes. |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
On considère qu'il existe 4 relations trophiques entre organismes (consommation, prédation, parasitisme, symbiose - voir cours de seconde), celles-ci pouvant être réalisées aussi bien entre autotrophes et allotrophes qu'entre allotrophes. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Remarque: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Objectifs techniques |
|
- Représenter un
écosystème sous forme d'un graphe indiquant
les types de relations trophiques. (Belin
schéma 1 p 10 et réseau p 18; exercices 4 p 25
et 6 p 25; pyramide p 20) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
1.2 - La nutrition, vue par le biochimiste, est une série de transformations (oxydoréductions, transferts de liaisons et de groupements) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
En considérant le vivant comme un système chimique, plus ou moins analogue à une solution aqueuse, la chimie ne permet que fort peu de comprendre les spécificités du vivant (ce qui fait l'originalité de la vie). Elle ne doit cependant pas être négligée ne serait-ce qu'à cause du phénomène de mode qui la soutient notamment dans les crédits de recherche. |
|
Classiquement on dit que la chimie n'a acquis son statut scientifique qu'à la fin du XVIIIème siècle (avec notamment Lavoisier, vers 1770), ce qui en fait la plus jeune des sciences expérimentales. La chimie est la science des transformations de la matière, en ce sens elle fait partie de la physique, science des corps inanimés. En leur accolant le préfixe bio- (la biochimie, la biophysique) on cherche moins à étendre des frontières qu'à montrer des résonances. En effet l'être vivant, animé (voir introduction au cours de seconde), sort du domaine de la chimie et vouloir le prendre comme objet ne peut que réduire notre compréhension du vivant. Par contre, utiliser les méthodes de la chimie pour comprendre le vivant, dans une authentique biologie moléculaire et cellulaire est plus prometteur. |
|
Base de la classe de seconde à
réinvestir : Bases de la classe de première S à réinvestir : - les molécules du vivant (voir éléments de biochimie) - les réactions d'oxydoréduction (cours de chimie) - la notion de réactions couplées - les enzymes |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Un bilan, déjà vu en seconde (que l'on va reprendre sans les couleurs, car notre but cette année et de justifier leur emploi): |
|
Étant donné leur symétrie on les réunit dans une seule équation bilan. Mais ceci n'est possible que pour les autotrophes car les allotrophes n'ont pas de photosynthèse. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Dans cette partie nous introduirons un vocabulaire de la thermodynamique pour l'étude des réactions métaboliques et s'efforçant d'énoncer des bilans au niveau de l'organisme ou des cellules sans trop se préoccuper, pour l'instant, de leur support biologique, même si nous le citerons. Je répète qu'un chimiste ne s'intéresse qu'à la matière. On commencera par la respiration, qui est peut-être plus facile à comprendre et plus générale que la photosynthèse. La nature des échanges de matière a déjà été vue en classe de seconde (TP Levures et Euglènes habituellement). |
|
Les structures et dynamiques qui, depuis un niveau supérieur de compréhension, contraignent et ordonnent cette matière lui sont inaccessibles (voir le cours de 1ère S sur les enzymes où une voie alternative au réductionnisme, qui est appelée "structuralisme", est présentée avec l'exemple de la fonction enzymatique). Je présenterai plus bas quelques ouvertures vers l'autre niveau de compréhension issu de la Théorie des catastrophes de René Thom ( une catastrophe, c'est une discontinuité phénoménologique (le bord d'un nuage, un tourbillon, le déferlement d'une vague, la naissance d'une bulle, la mitose, l'émission d'un pseudopode, le battement d'un cil...). Pour illustrer le niveau structural (géométrique) en regard du niveau réductionniste (thermodynamique) voici un extrait de Prédire n'est pas expliquer, René Thom, 1991, Eshel, Paris, dans la collection «La Question» dirigée par Émile Noël; « Mais les singularités apparaissent lorsque l'on soumet en quelque sorte l'espace à une contrainte. La manche de ma veste, si je la comprime, je fais apparaître des plis. C'est une situation générale. Cela ne relève pas de la mécanique des matériaux. J'énonce en réalité un théorème abstrait : lorsqu'un espace est soumis à une contrainte, c'est-à-dire lorsqu'on le projette sur quelque chose de plus petit que sa propre dimension, il accepte la contrainte, sauf en un certain nombre de points où il concentre, si l'on peut dire, toute son individualité première. Et c'est dans la présence de ces singularités que se fait la résistance. Le concept de singularité, c'est le moyen de subsumer en un point toute une structure globale.» Il n'y a plus de matérialité (au sens physique) : la nature du matériau n'interviennent pas dans la genèse de la forme. On comprend mieux ainsi le mot de Thom : « Le but idéal de la théorie des catastrophes, c'est de remplacer la thermodynamique par la géométrie », Thom, 1977f17.pdf, p2). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Questions / |
|
J'utilise ici le procédé pédagogique classique d'un questionnement réalisé en cours dont voici la retranscription a - Peut-on écrire un bilan pour les allotrophes ? b - Quelles sont les molécules intermédiaires "transporteuses d'énergie" ? c - D'où vient le CO2 produit par respiration et comment est consommé l'O2 lors de cette respiration ? d - Y-a-t-il d'autres substrats respiratoires que le glucose ? e - Que respire l'autotrophe ? f - Si un autotrophe peut respirer et photosynthétiser quel est le bilan global de son métabolisme ? g - D'où vient le dioxygène produit par photosynthèse ? h - Comment le CO2 est-il incorporé à des glucides ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
a - Peut-on écrire un bilan pour les allotrophes ? |
|
OUI |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
premier bilan respiratoire des allotrophes |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
On utilise judicieusement la notion d'enthalpie libre vue en cours de physique-chimie (l'enthalpie libre (notée G) est la fonction thermodynamique associée à la variation d'énergie d'un système avec variation de volume). Une transformation (et donc une variation de la fonction G entre les produits et les réactifs) avec un �G <0 est dite exergonique et une transformation avec un �G>0 est dite endergonique (pour �G = 0 on est censé avoir l'équilibre et le sens de la réaction n'est pas déterminé). Une réaction endergonique ne peut se réaliser sans apport d'énergie. |
|
L'équation globale de la respiration à un �G°' de -2840 kJ.mol-1 (le ° indiquant les conditions standard des physiciens et chimistes (à pH =0) et le ' le standard (à pH=7) spécifique aux biochimistes, on pourra bien sûr omettre cette notation en SVT en terminale !!!!), elle est donc fortement exergonique et peut fournir de l'énergie à la cellule. On se nourrit de l'énergie contenue dans notre nourriture (nous sommes des chimiotrophes). Cette énergie est "récupérée par des molécules" dans le sens où elle sert à réaliser des réactions chimiques endergoniques. Les molécules qui sont produites lors des réactions exergoniques de la respiration sont dites intermédiaires (ou "transporteuses d'énergie") car ON PENSE qu'elles peuvent être utilisées loin (relativement !!) de leur lieu de production (mais toujours DANS la cellule). Par exemple, les molécules produites dans la mitochondrie pourraient aller ensuite dans le cytoplasme cellulaire. Leur retour à un niveau d'énergie plus faible permet de réaliser des réactions chimiques variées qui sont le travail de la cellule. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
bilan complémentaire : |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
soit le bilan général: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Si on supprime par la
pensée les molécules intermédiaires, on
a bien le bilan de la nutrition |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
BILAN nutritionnel des allotrophes: aliments (substances organiques représentées par le glucose), comburant (le dioxygène), déchets (le dioxyde de carbone et l'eau), travail |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Remarque: |
|
Avec notre parti-pris chimique, il est évident que l'on ne connaît pas de travail qui ne passe pas par la matière (il faut des molécules pour l'exprimer). L'énergie chimique ne peut se transférer qu'à une molécule qui peut se transformer par ajout ou perte de groupement (travail chimique), se déplacer (travail mécanique), se charger électriquement ou déplacer des charges électriques (travail électrique). Mais tout le travail de la cellule ne passe pas forcément par cette énergie chimique. Par exemple, il y a le travail des forces extérieures qui agissent sur (contraignent) le cytoplasme et les structures cytoplasmiques. Un système vivant n'a pas que de l'énergie interne tirée de sa nourriture matérielle. Peut-être pourrait-on dire qu'il y a d'autres formes de nourriture que la nourriture matérielle. Placée dans un champ de gravité la cellule peut l'exploiter pour en tirer de l'énergie. Placée dans un champ thermique, l'organisme sait en prélever une partie et l'accumuler. Gravité et chaleur sont tout autant de nourritures. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
b - Quelles sont les molécules intermédiaires "transporteuses d'énergie" ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Exercice n°7 p 83 (Belin) |
|
La glycolyse est un ensemble de réactions chimiques qui ont lieu dans le cytoplasme de toutes les cellules eucaryotes. |
|
On considère que la liaison énergétique entre le phosphate inorganique et l'ADP nécessite ou fournit de 50 à 65 kJ.mol-1 dans les conditions intracellulaires (on a calculé 51,8 pour le globule rouge intact alors que la valeur est de 30,5 kJ.mol-1 dans les conditions standard). |
|
La seule subtilité de l'exercice nécessite de faire attention au nombre de molécules impliquées. Un sucre en C6 est le point de départ et deux molécules en C3 sont les points d'arrivée. Une présentation des formules développées des molécules serait bien plus pédagogique. (glucose et fructose en C6; dihydroxyacétone, glycéraldéhyde et glycérates en C3). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Pour des �G voisins de 0 les réactions se font avec des enzymes. Les �G voisins de 30 kJ-.mol-1 nécessitent une molécule d'ATP. Et inversement, les �G voisins de -30 kJ.mol-1 fournissent une molécule d'ATP. Soit au total 2 molécules consommées et 4 produites, ce qui fait un bilan net de 2 produites par molécule de glucose transformée en 2 pyruvates. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Questions complémentaires |
|
Écrire en terme de bilan l'équation de la glycolyse (la formule d'un pyruvate est CH3COCOOOH). La compléter sous forme d'une somme de deux réactions couplées en ajoutant le transfert d'énergie chimique de liaison. |
|
glycolyse C6H12O6 2 CH3COCOOH 2 (ADP + Pi) 2 ATP |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Pourquoi l'équation n'est-elle pas équilibrée ? Ajouter les molécules nécessaires pour équilibrer l'équation. |
|
On notera les transporteurs d'énergie d'oxydoréduction R sous la forme oxydée et RH2 sous la forme réduite; je préfère cette notation à Rox et Rréd du fait de la présence des protons en plus des électrons dans la forme réduite.
Pour donner une idée de l'énergie des réactions d'oxydoréduction où sont impliqués les coenzymes on peut citer par exemple le �G°' de l'oxydation du NADH+ H+ (forme RH2) en NAD+ (forme R, NAD signifiant nicotinamide adénine dinucléotide) qui est de -220 kJ.mol-1. Ce qu'il faut comparer à la déphosphorylation de l'ATP en ADP qui a un �G°' voisin de -50 kJ.mol-1 (voir ci-dessus). |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Résumé |
|
Les molécules intermédiaires "transporteuses d'énergie" sont d'une part les nucléotides énergétiques ou mieux nucléotides triphosphate (ATP et GTP principalement qui sont hydrolysés en ADP, AMP ou GDP...) et d'autre part des molécules dites "transporteurs d'électrons et de protons" ou mieux coenzymes d'oxydoréduction car ce sont de petites molécules comportant des nucléotides (très modifiés) et associées à des protéines qui interviennent comme enzymes dans les réactions d'oxydoréduction. |
|
Il est souvent commode de
parler de pouvoir réducteur de la
cellule, désignant ainsi le nombre de coenzymes
réduites disponibles à un instant
donné. Possédant des molécules
intermédiaires énergétiques
réduites, une cellule peut oxyder d'autres substrats,
ce qui est le plus souvent nécessaire dans les
synthèses. |
|
Les réactions du catabolisme sont principalement des oxydations, qui permettent de récupérer des coenzymes réduits, et des réactions exergoniques qui permettent de récupérer de l'énergie chimique de liaison sous forme de nucléotides triphosphate. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Deux exemples (bien évidemment pas à apprendre) pour ne pas avoir des réactions imaginaires: |
|
AldPG --> ABPG lors de la glycolyse (et permet de récupérer un transporteur réduit)
Pour les
élèves TRÈS ATTENTIFS, |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
pyruvate ---> éthanol lors de la phase
terminale de la fermentation alcoolique (qui
fait suite à la glycolyse)
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Comme il est regrettable
que les enseignants de biologie aient pris la fâcheuse
habitude de ne pas écrire les demi-réactions
avec les électrons, nous allons y remédier
pour une fois. Cela est
dû, je pense, à la complexité des
réactions (et à leur méconnaissance).
Car on écrit souvent des bilans et non de
véritables réactions. On pourrait y
remédier si on nous allégeait nettement le
programme . Il faudrait alors bien sûr se contenter de
quelques réactions et non pas de la respiration tout
entière.
réduction de
l'acétaldéhyde oxydation du coenzyme oxydoréduction |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Compléments sur l'ATP |
|
Si l'on fait un bilan de la fermentation alcoolique, on obtient
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
c - D'où vient le CO2 produit par respiration et comment est consommé l'O2 lors de cette respiration ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Nous avons vu que la glycolyse a lieu dans l'hyaloplasme des cellules et produit du pyruvate. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Analyse de documents (Belin A1 p 72, Didier Doc1 p 116) |
|
Le pyruvate est
oxydé en CO2 et H2O dans les
mitochondries. Ces réactions complexes comprennent: |
|
Remarque: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
La plupart des réactions chimiques proposées pour la cellule ont été mises en évidence sur des fractions cellulaires broyées centrifugées, solubilisées... parfois marquées radioactivement... bref une chimie de l'homogène (du tube à essai) alors que le cytoplasme est fondamentalement hétérogène, et organisé alors que les méthodes sont destructives de structures fines du cytoplasme. Ce qui nous amènera à une autre voie de recherche. En tout cas, cela justifie que l'on donne aux élèves les résultats de ces expériences sans vous en montrer les techniques. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bilan général biochimique de la fermentation alcoolique et de la respiration (même bilan avec la fermentation lactique) (Belin p 81) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fermentation alcoolique |
|
glycolyse C6H12O6 2 CH3COCOOH 2
(ADP + Pi) 2 ATP 2 R 2
RH2 phase finale 2 CH3CH2OH
+ 2 CO2 2 CH3COCOOH Le bilan est de 2 moles d'ATP
produites par mole de glucose consommée;
tous les transporteurs réduits (2 RH2)
sont réoxydés (2 R) et donc aucun
n'est utilisable par la cellule. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Respiration |
|
glycolyse C6H12O6 2 CH3COCOOH 2
(ADP + Pi) 2 ATP 2 R 2
RH2 phase
mitochondriale matrice <------
10 RH2 10 R 2 ATP 2 (ADP + Pi) 6
CO2 2 CH3COCOOH
+ 4 H2O crêtes phosphorylation oxydative 6
O2 12
H2O 12 RH2 12 R 32
(ADP + Pi) 32 ATP Le bilan est de 36 moles ATP
(32+2 +2) produites par mole de glucose
oxydée; dans ce bilan tous les
transporteurs réduits (12 RH2)
ont été réoxydés (12
R) mais certains peuvent servir dans le
cytoplasme ou dans la mitochondrie pour
des synthèses. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
On s'est efforcé en seconde d'utiliser le terme ventilation pour les mouvements de gaz au niveau des organes ou de l'organisme. |
|
Le terme respiration signifie respiration cellulaire . Elle existe pour tous les eucaryotes, même si certaines cellules peuvent perdre cette capacité et notamment acquérir la capacité de fermenter. Les cellules musculaires par exemple présentent une fermentation de type lactique (qui produit de l'acide lactique mais pas de CO2 - non au programme). Les levures comme Saccharomyces cerevisiae, ont un métabolisme mixte: fermentaire (alcoolique) ou respiratoire, selon les conditions de milieu. On les utilise grandement dans l'industrie alimentaire (pain, boissons fermentées, produits laitiers fermentés.... |
|
On notera enfin qu'une cellule qui fermente (et ne respire pas, du moins à un moment donné) peut vivre en anaérobiose dans un milieu sans dioxygène. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
d - Y-a-t-il d'autres substrats respiratoires que le glucose ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Oui (indirectement) et non (directement). |
|
Les réactions chimiques présentées ici ne sont bien sûr pas exhaustives, il existe plusieurs autres voies alternatives, mais pas si nombreuses que cela. De même, de nombreuses molécules peuvent directement rentrer dans le cycle de Krebs par exemple (où en être retirées). Bref, le métabolisme forme un réseau d'équilibres reliés les uns aux autres. |
|
Remarque: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Revenons maintenant à l'équation-bilan double respiration-photosynthèse des autotrophes. |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
e - Que respire l'autotrophe ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Les molécules organiques qu'il synthétise lui-même. |
|
Sa respiration se fait, comme pour les allotrophes dans son cytoplasme (glycolyse) puis dans ses mitochondries. Le substrat est toujours le glucose qui est le plus couramment issu de l'hydrolyse de l'amidon que la cellule fabrique et stocke transitoirement dans ses chloroplastes (par photosynthèse). |
|
Les cellules non chlorophylliennes reçoivent le glucose sous forme de saccharose (voir cours de 1ère S - annexe de biochimie) qui circule dans la sève élaborée (Belin schéma p 37, nous y reviendrons dans la partie 2). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
f - Si un autotrophe peut respirer et photosynthétiser quel est le bilan global de son métabolisme ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Au niveau de l'organisme, on pourrait croire qu'il est assez facile de comparer les parts respectives de la respiration et de la photosynthèse du fait des gaz produits et consommés. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
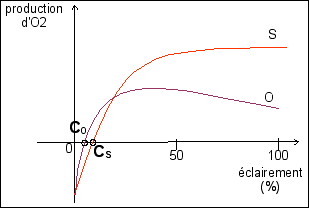
Analyse de courbes: consommation-production de dioxygène par des plantes chlorophylliennes. (elles sont expliquées dans l'exercice corrigé Belin p 96) |
|
 |
|
Rappels de seconde sur la technique de l'analyse de courbes. La mesure de la production de dioxygène (qui est appelée photosynthèse nette) en fonction de l'éclairement permet de le mettre en évidence que photosynthèse et respiration sont en compétition dans une plante chlorophyllienne. Toutes les cellules ne peuvent pas photosynthétiser mais toutes respirent. Cs et Co indiquent les points de compensation entre photosynthèse et respiration pour les deux plantes S et O. Définissez mathématiquement et biologiquement le point de compensation C.
La tomate est une plante héliophile (qui aime le soleil), son point de compensation se situe entre 700 et 1.000 lux. Les fougères ou les mousses sont sciaphiles (aiment l'ombre) et ont des points de compensation aux alentours de 100 Lux. Expliquez. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Mais le bilan est difficile à établir, car il peut y avoir un décalage dans le temps et dans l'espace entre les différentes réactions chimiques: les gaz ou les intermédiaires métaboliques peuvent être stockés et les réactions différées dans le temps. |
|
Ce qui est certain est qu'une cellule chlorophyllienne d'une plante peut photosynthétiser à la lumière et respire toujours. À l'opposé une cellule non chlorophyllienne ne peut que respirer. On notera donc que les termes autotrophes et allotrophes ne s'appliquent qu'aux organismes et non aux cellules d'un pluricellulaire (même si on le fait parfois de façon erronée). |
|
J'insiste, car il n'est pas rare de voir des questions, même dans des sujets de bac, sur les "cellules hétérotrophes des autotrophes". Je suis partisan sans hésitation de "corriger" la phrase et de préciser, à l'attention du correcteur, que l'on a compris que l'on parle de cellules d'un autotrophe qui sont non photosynthétiques mais qui respirent ou fermentent à partir de nutriments qui viennent des parties photosynthétiques de la plante. On pourrait ajouter que même les cellules photosynthétiques reçoivent des éléments nutritifs (au sens "d'intervenant dans le métabolisme de nutrition") en provenance d'autres cellules, ne serait-ce que lors des changements de saison... |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
g - D'où vient le dioxygène produit par photosynthèse ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Analyse d'une expérience fondamentale utilisant des isotopes radioactifs (Ruben et Kamen, 1940) (Belin p 51 B3, Nathan p 20, 2) |
|
Une suspension d'algues vertes unicellulaires fortement éclairées en présence de CO2 non radioactif, mais dans l'eau lourde (marquée à l'oxygène 18O) produit du dioxygène marqué au 18O. |
|
On peut donc améliorer l'équation bilan de la photosynthèse en colorant l'oxygène de l'eau et celui du dioxygène produit
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Les réactions chimiques qui conduisent à la production du dioxygène à partir de l'eau sont connues sous le nom de photooxydation de l'eau. Comme toute réaction chimique d'oxydation, elle nécessite une réduction couplée. |
|
Exercice complémentaire: Nathan n°2 p 29, Didier p 130 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Analyse d'une expérience sur la réaction de Hill (1941) à partir d'une suspension de chloroplastes (Belin p 50 A1) |
|
On a donc un accepteur d'électrons et de protons (réactif de Hill = 2,6 dichlorophénol indopténol - ici le ferrocyanure de K) qui, lorsqu'il est réduit, permet l'oxydation de l'eau et donc le dégagement de dioxygène. On a donc une réaction fondamentale à la lumière dans le chloroplaste (plus précisément au niveau de complexes dans les membranes des thylakoïdes): |
|
Cette réaction est toujours appelée "réaction de Hill" en hommage à son découvreur. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
La cellule serait bien inefficace si elle n'en profitait pas pour récupérer de l'énergie chimique de liaison. Les mécanismes en sont complexes et nécessitent aussi la lumière. Le phénomène général est nommé photophoshorylation. Le bilan n'est pas facilement calculable (certaines réactions sont cycliques avec uniquement formation d'ATP sans réduction de transporteurs, d'autres produisent les deux types d'énergie...) mais est de l'ordre de 2 à 4 molécules d'ATP par molécule d'eau oxydée. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Le bilan global est donc approximativement : |
|
au niveau des thylakoïdes des chloroplastes à la lumière (phase photochimique).
2 H2O O2 2 R 2 RH2 3 ou 4(ADP + Pi) 3 ou 4 ATP |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
h - Comment le CO2 est-il incorporé à des glucides ? |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Analyse de documents (Belin p 52 A1-2 et p 53 B3-4) - Expérience de Calvin (1961) - courbe plus exacte du Didier exercice n°5 p 106 |
|
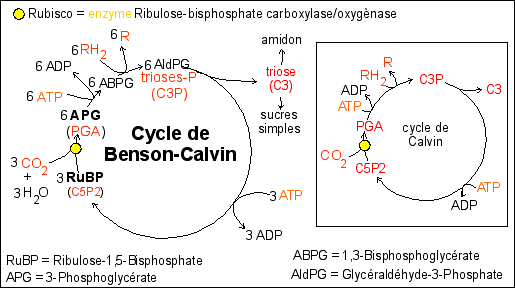
La matrice du chloroplaste comporte donc un cycle (cycle de Benson-Calvin) comportant environ 15 composés dont le principal est le Ribulose-1,5-Bisphosphate (RuBP ou C5P2 sur votre livre Belin) sur lequel le CO2 est fixé pour donner deux molécules de 3-Phosphoglycérate (APG ou PGA en anglais). Les molécules d'APG sont phosphorylées (ce qui nécessite 3 ATP). La réduction a ensuite lieu (c'est l'APG qui est réduit en AldPG, ce qui est la réaction inverse de celle qui se passe lors de la glycolyse - voir ci-dessus) et utilise le pouvoir réducteur des RH2 produits pendant la phase photochimique. Des sucres sont produits (certains sont stockés dans le chloroplaste sous forme d'amidon et d'autres sont exportés vers le cytoplasme de la cellule). Une dernière étape régénère le RuBP ce qui demande aussi de l'ATP. |
|
Le bilan global pour un tour de cycle est de l'ordre de 3CO2 fixés, 6 RH2 réoxydés en R, 9 ATP consommés. Pour former un glucide à 6 carbones, on peut donc considérer qu'il faut:
6 CO2 C6H12O6
+ 6 H2O 12 RH2 12 R 18 ATP 18 (ADP + Pi) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Remarque: |
|
L'encadré à droite ne
présente strictement que les éléments
au programme à mémoriser avec les sigles
demandés par le programme. Exercice complémentaire: Belin p 62 n°7 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bilan général de la photosynthèse |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
phase photochimique (membranes des thylakoïdes des
chloroplastes) 12 H2O 6 O2 18 à 24
(ADP + Pi) 18 à 24 ATP 12 R 12 RH2 phase chimique (matrice des chloroplastes) 18 (ADP + Pi) 18 ATP 6 H2O
+ C6H12O6 6 CO2 |
|
Tous les transporteurs réduits pendant la phase photochimique (12 RH2) sont réoxydés pendant la phase chimique (12 R), mais toutes les molécules d'ATP ne sont pas forcément consommées. Le bilan énergétique est donc de l'ordre de 6 moles d'ATP par mole de glucide produite (soit une mole d'ATP par mole d'O2 libérée). |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Remarque: |
|
(photo)oxydation de l'eau qui correspondrait à
réduction du
1,5-Bisphosphoglycérate (ou
APG) en AldPG |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Exercice |
|
On en revient donc maintenant à
l'équation bilan
des autotrophes vue en
seconde. |
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
En ce qui concerne la respiration (1), même s'il y a oxydation totale du glucose, les 6 carbones ne sont pas simultanément rejetés sous forme de CO2. Certains s'incorporent à des molécules du cycle de Krebs et peuvent naviguer dans le métabolisme de la cellule et n'être oxydés que bien plus tard. |
|
C'est la même chose pour la photosynthèse (2): le CO2 incorporé ne se fixe que sur un corps en C5 du cycle de Benson-Calvin pour donner un corps en C3 qui possède le carbone incorporé et un autre qui n'en possède pas de nouveau. Le carbone incorporé peut se retrouver ensuite dans un acide aminé dans les protéines ou dans un lipide. Il n'y a aucun glucide composé de 6 CO2 fixés ensemble qui se forme. |
|
En ce sens, les bilans sont faux, même si, statistiquement , on peut affirmer qu'au bout d'un grand nombre de cycles les carbones d'un glucide proviennent de l'air. De la même manière, les
électrons pris à l'eau lors de la
photooxydation se retrouvent sur les transporteurs
réduits et donc finalement sur l'AldPG, mais ils ne
peuvent que très occasionnellement se retrouver dans
les glucides formés. Aucune molécule de
glucose formée ne possède des protons tous
issus de l'eau. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Enfin, l'intérêt de superposer ces deux bilans reste limité, car l'équilibre ou la réciprocité ainsi suggérés finalement ne correspondent qu'à la nature des molécules mises en jeu. D'où provient cette similitude ? Non de l'unité du vivant (des métabolismes voisins) mais justement de la non-spécificité au regard de la matière. La diversité des métabolismes est autre part. Comme nous l'avons vu, l'autotrophe est un producteur et l'allotrophe un consommateur. Il faut donc revenir à l'organisme si l'on veut comprendre quelque chose à la nutrition. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Laissons maintenant les chimistes et parlons en biologiste. |
|
|
||||||||||||
|
En grec «métabolè» signifie «changement» |
|
Il est temps d'affirmer que le métabolisme n'est pas un ensemble de réactions chimiques dans un milieu particulier. La vie n'est pas (seulement) un ensemble de réactions chimiques. |
|
Le métabolisme c'est le mouvement de la vie. Ce sont des transformations de la matière particulières propres à la vie (vitales) mais aussi des dynamiques. |
|
Si l'on utilise le vocabulaire des biologistes, il y a trois dynamiques, 3 grandes fonctions (ou travail): il existe un métabolisme nutritionnel, un métabolisme relationnel et un métabolisme reproducteur; ce qui produit le mouvement (ce qui maintient stables les dynamiques) ce sont les fonctions. Nous nous intéresserons au travail de nutrition. |
||||||||||
| |
|
1.3 - Se nourrir est une fonction homéostatique |
||||||||||||||
| |
|
|
|
Bases de la classe de seconde à réinvestir : |
|
les trois types de travail du vivant
(ou grandes fonctions): nutrition,
reproduction et relation |
||||||||||
| |
|
L'unité du vivant repose moins sur l'universalité de la chimie (qui justement la réunit au non-vivant) que sur sa dynamique propre que l'on regroupe sous le terme de fonctions globales ou de travail. Seuls les systèmes vivants sont capables de se nourrir, de se reproduire et communiquer entre eux (voir cours de seconde). |
|
On a coutume de dire que le travail de nutrition consiste à échanger de la matière et de l'énergie avec le milieu extérieur. Mais le terme d'échange est inapproprié étant donné la radicale différence qu'il existe entre l'être vivant et son milieu et l'inégalité des volumes échangés. L'organisme vit au dépens de son milieu: il le consomme même s'il en fait partie. Il ne cesse de lutter pour exister en dehors de lui (ou être enfanté par lui), malgré les contraintes qui l'assaillent. Il est donc plus exact de dire que le travail de nutrition assure le maintien (la stabilité) de la forme de l'organisme (structure et dynamique) . |
|
La stabilité est une des caractéristiques des dynamiques du vivant. Ce n'est pas le système vivant qui est stable (ou immobile) mais sa dynamique, c'est-à-dire ses fonctions. On dit encore que l'organisme est en équilibre dynamique (qu'il fonctionne), ce qui est le propre d'un système en homéostase. Ce terme vient du grec homéo (ou homo = identique) et stase qui signifie position (voir cours de 1èreS). La propriété d'un système en homéostase est l'homéostasie. Un système homéostatique est un système qui est stable face à de petites perturbations. Nous avons vu en seconde que le premier principe de dynamique du vivant (ou principe d'homéostasie) affirme qu'un être vivant est un système homéostatique (ou en homéostase) et donc que les fonctions du vivant sont STABLES (résistantes à de petites perturbations). |
||||||||||
| |
|
|
|
|
||||||||||||
| |
|
Si l'on utilise un vocabulaire analogue à celui des économistes ont peut dire que, pour survivre et par la nutrition, l'organisme vivant compense ses pertes de matière et d'énergie et s'autorise un renouvellement de ses structures (toute structure dynamique est en perpétuel renouvellement: ce qui est vrai pour les molécules, mais aussi pour les formes - et la forme est vraiment contrainte par le type de nourriture et le mode de nutrition -). Le bilan peut donc s'établir comme une somme de besoins qui doit couvrir les dépenses. |
|
On remarquera que du point de vue physiologique (du fonctionnement) la nutrition comprend la capture de la nourriture (matière et énergie), son assimilation (ses transformations éventuelles) et le rejet des déchets produits. Mais on peut aussi considérer que les systèmes de communication entre cellules qui assurent le maintien de l'intégrité de l'organisme font partie de ce travail de nutrition: ce sont certains des mécanismes que l'on classe actuellement dans les mécanismes immunitaires. |
|
Pour illustrer la diversité des modes
de nutrition, on peut s'attacher à l'anatomie,
à la morphologie...et à tant d'autres
critères classificatoires. Habituellement on classe
les animaux en fonction du type d'aliments qu'ils
ingèrent (liquide ou solide, micro et macrophage) et
du type de comportement mis en jeu pour leur capture
(filtreurs, chasseurs, suceurs nécrophages....). |
||||||||||
| |
|
|
|
|
||||||||||||
|
Un peu de "thomisme"... "Deviens ce que tu manges". On s'identifie à sa nourriture. |
|
Cette réflexion, peut-être anthropomorphique au départ (j'ajouterais que l'homme ne se nourrit pas que de pain, mais aussi de parole...), à des prolongements très justes en biologie (René Thom l'emploie principalement pour l'animal qui s'identifie à sa proie: "le prédateur affamé est sa proie"; voir la notion de lacet de prédation dans l'œuvre de Thom; voir aussi Les chemins du sens à travers les sciences, AL, [1984, 9. 3] (1984f9.pdf); auquel on peut aussi ajouter le travail de Philippe LACORRE, Sur un nouveau type de représentation catastrophiste pour les modélisations en biologie et sciences cognitive, Intellectica, 1997/1, 24, pp. 109-140). |
|
La nutrition est la satisfaction d'un besoin. Si l'on représente le métabolisme comme une dynamique correspondant à une forme dans un espace de grande dimension (voir page sur les modèles), un manque de nourriture est une instabilité. La fonction de nutrition globale peut être approchée, localement (par exemple au niveau d'une cellule), par un potentiel et l'acte de se nourrir par le retour du système à une valeur minimale du potentiel. La nutrition est toujours accomplie lorsque l'organisme minimise son potentiel. |
|
La nutrition allotrophe est une catastrophe de capture: deux actants, le prédateur et la proie, n'en font plus qu'un, à la fin de l'action. Il y a ici le cycle fondamental d'un retour à un niveau stable, antérieur à la capture et à l'assimilation (1967f8.pdf, p3s). |
||||||||||
|
2. Être autotrophe |
|
|
|
Bases de la classe de seconde à
réinvestir : Bases de la classe de première S à réinvestir : Données succinctes de morphogénèse végétale (partie non traitée dans le cours de 1ère S sur ce site, étant donné son aspect exagérément ambitieux - voir commentaires sur l'ancienne page de 1èresS, nouvelle page); on reprendra donc ici les éléments d'anatomie ou de morphologie végétale utiles. |
||||||||||||
|
Généralités: |
|
Pour un autotrophe, qui se nourrit d'air et de lumière on peut considérer qu'il existe trois actions à réaliser: boire, ventiler, capter la lumière. Selon son milieu de vie et sa taille, ces actions sont réalisées par des structures plus ou moins élaborées. |
|
(J'utilise ventiler et non pas respirer pour la raison évidente que les échanges gazeux chez un autotrophes ne se servent évidemment pas uniquement à la respiration cellulaire, mais aussi à la photosynthèse. Ensuite on pourrait ajouter les phénomènes d'excrétion, mais ils sont, chez un autotrophes beaucoup moins importants que pour un allotrophe, car les autotrophes augmentent bien davantage leur taille (ne cessent de croître ou/et de se diviser) et les rejets des autotrophes sont très faibles, à moins que l'on ne considère l'eau qui présente un véritable flux à travers la plante). |
|
|||||||||||
| |
|
2.1 Une bactérie bleue (Procaryote, Cyanophycée ou Cyanobactéries) |
||||||||||||||
|
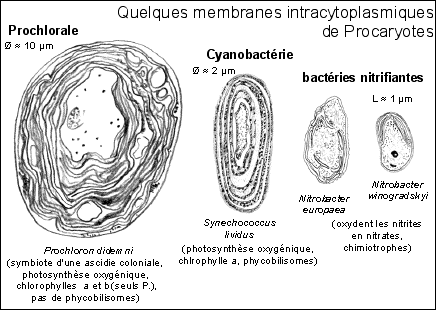
Schéma de comparaison entre la structure (l'ultrastructure quand elle est dévoilée à partir de coupes colorées observées au ME) d'un chloroplaste (Belin p 46, 55, 59, 86) et d'une Cyanophycée (Didier p 136, Les cyanobactéries, d'étonnants procaryotes autotrophes, Marc-André Selosse, Biologie-Géologie (Bulletin de l'APBG), n°3-1996, 481-529 ). |
|
|
||||||||||||||
| |
Ces Procaryotes aquatiques (souvent mutualistes) forment le plus souvent des colonies (masse mucilagineuse, trichomes (filaments formés de files de cellules issues de la division d'une cellule initiale) ou parenchymes (feuillets)...). |
|
Il existe plusieurs types cellulaires pour une même espèce (les hétérocystes sont des cellules spécialisées dans la fixation du diazote atmosphérique, mais il existe de nombreux autres types cellulaires : baeocytes mobiles, akinètes de résistance, nécridies en dégénérescence, hormogonies riches en vacuoles à gaz...). |
|
Plus de 2.000 espèces avaient été décrites (mais on les réduit actuellement à près de 62 espèces et 24 genres). |
|||||||||||
| |
|
|
|
|||||||||||||
|
Boire et ventiler |
|
On PENSE que les gaz, l'eau et les solutés, diffusent à travers la paroi et la membrane plasmique puis dans le cytoplasme sans qu'il soit nécessaire de posséder un système d'échange et de transport des gaz ou des liquides. On notera cependant que, comme chez toute cellule procaryote, il n'y a pas de mouvements intracytoplasmiques de vésicules. Les substances transportées dans le cytoplasme le sont donc individuellement par des molécules transporteuses, soit diffusent seules, ce qui n'est pas sans poser d'intéressants problèmes de compréhension quant aux mécanismes permettant d'atteindre des vitesses de réaction métaboliques raisonnables (regroupement des enzymes en complexes, chanelling - système de transfert des composés d'une chaîne métabolique aux différentes enzymes regroupées...). |
||||||||||||||
|
Capter la lumière |
|
Capter la lumière donne lieu à des empilements membranaires voisins de ceux des chloroplastes. On les qualifie d'ailleurs de thylakoïdes et on leur attribue le même rôle que les membranes des grana des chloroplastes. Cependant, toutes les bactéries photosynthétiques ne présentent pas des thylakoïdes et les membranes internes existent chez des bactéries non photosynthétiques.
Remarque (voir endosymbiose ci-dessous) Les Prochlorales, avec leur systèmes photosynthétiques à chlorophylle a et b, sont proches des chloroplastes des eucaryotes. Ils se rapprochent des Cyanobactéries par la séquence de leurs ARNr (composants des ribosomes). |
|
 |
||||||||||||
| |
|
2.2 Une algue verte unicellulaire (Protiste) |
||||||||||||||
|
|
|
Les Chlorophytes (algues vertes unicellulaires isolées (comme ici) ou le plus souvent en colonies (comme les Enteromorphes de nos marées vertes)...) présentent plus de 7500 espèces ; elles possèdent les chlorophylles a et b et réalisent une photosynthèse oxygénique. Leurs réserves sont habituellement de l'amidon, mais peuvent aussi être composées d'autres sucres polymérisés (fructanes par exemple, polymères de fructose). La paroi composée de cellulose et de mannanes (polymères de mannose, isomère du glucose) est souvent minéralisée par du carbonate de calcium. Toutes les algues vertes sont classées dans le royaume des Protistes. Seules les algues rouges et brunes restent dans le royaume des plantes étant considérées comme de véritables pluricellulaires. Les algues rouges possèdent des phycobilisomes et possèdent la chlorophylle a seule (parfois la d). |
|
Culture de quelques unicellulaires de l'eau croupie, Elodée (unicellulaire coloniale: "pseudo"-pluricellulaire du royaume des Protistes). Expérience d'Engelmans.... Extraction de chloroplastes ? (Cabomba ?); non réalisable sans centrifugeuse. mouvements de cyclose (déplacements) |
||||||||||||
| |
|
La nouveauté de la cellule eucaryote est la présence de vésicules qui peuvent transporter des substances (liquides ou gazeuses) et bien sûr des chloroplastes et mitochondries. |
|
On notera que le dioxygène est PARTICULIÈREMENT TOXIQUE (oxydant). Il ne peut pas être libéré dans la cellule sans précautions. |
||||||||||||
| |
On a émis l'hypothèse que les mitochondries et les chloroplastes des cellules eucaryotes dériveraient de Procaryotes symbiotiques. |
|
La formulation du programme peut faire sourire: "La mitochondrie et le chloroplaste proviennent probablement de bactéries qu'une cellule hôte ancestrale aurait adoptées comme endosymbiotes." D'une part à cause de la simultanéité sous-entendue et d'autre part étant donné la part énorme de l'hypothèse dans cette interprétation, telle qu'elle est actuellement proposée. Cela dit, l'idée est séduisante et peut-être plausible. Il reste à l'argumenter de façon scientifique ce qui n'est pas une mince affaire puisqu'il s'agit de phénomènes passés. (<---) |
|||||||||||||
| |
|
2.3 Une Plante |
||||||||||||||
|
Anatomie
d'une plante en relation avec le travail de nutrition: |
|
Plant de tomate ou de Pomme de
terre, racines, tige, feuilles (Belin p 41); |
||||||||||||||
|
|
||||||||||||||||
|
Études complémentaires |
|
Ultrastructure de trois cellules de
Plante: des cellules avec des plastes (nom
générique pour désigner les organites
pouvant évoluer en chloroplastes)
très variés |
|
* (cellule méristèmatique
de la racine de Pois (paroi souple primaire,
proplastes avec grain d'amidon, rapport
nucléocytoplasmique élevé), |
||||||||||||
| |
|
|
||||||||||||||
| |
Une plante est un organisme autotrophe aérien (même si quelques plantes sont aquatiques nécessitant des systèmes particuliers d'approvisionnement en gaz et de reproduction) et pluricellulaire. |
|
La ventilation est donc habituellement facile et peut se réaliser par un flux gazeux qui pénètre et ressort par de nombreux orifices (les stomates). Les gaz circulent entre les cellules d'un tissu lâche (le parenchyme aérifère) et dans les espaces naturels (les méats) laissés entre les cellules des tissus plus denses comme les épidermes. |
|
Toutes les cellules respirant, elles ont besoin de gaz (dioxygène). Celles pratiquant la photosynthèse sont par contre dépendantes de la teneur en CO2. Celle de l'air est faible (0,04%) mais il existe d'innombrables mécanismes biochimiques qui permettent d'augmenter la quantité de CO2 réellement disponible au niveau des centres pigmentaires photosynthétiques, notamment du fait des équilibres avec les formes dissoutes de ce gaz: HCO3- et CO32-. Comme nous aurons à revoir ce point dans d'autres parties du programme autant en parler tout de suite (pour des détails voir page sur les carbonates). |
|||||||||||
| |
|
|
||||||||||||||
|
les carbonates (site iufm) |
|
Le dioxyde de carbone atmosphérique est tout d'abord dissous dans l'eau. Mais il ne reste pas sous forme de gaz et réagit avec les éléments de l'eau: les ions H+ (proton ou H3O+) et les ions OH- (hydroxy). Ces réactions sont à la base de la formation des sédiments carbonatés qui donnent notamment toutes les roches calcaires.... Dans une cellule le transport du CO2 est donc principalement celui d'ions bicarbonate (HCO3-), ce qui se fait en liaison avec différentes substances organiques. |
|
Le dioxyde de carbone est en équilibre avec l' acide carbonique: H2CO3. L'acide carbonique en solution se dissocie en 2 H+, les protons, et HCO3- , l'ion bicarbonate ou hydrogénocarbonate. |
|
Cet ion peut, plus difficilement et très transitoirement, se dissocier en H+ et en CO32-, l'ion carbonate : |
||||||||||
| |
|
|
|
|||||||||||||
| |
|
Le dioxygène est par contre très toxique dès qu'il donne naissance à des radicaux libres oxygénés comme l'anion superoxyde O2- ou le radical hydroxyle HO-. Divers systèmes "anti-radicaux libres" sont connus comme le monoxyde d'azote. |
||||||||||||||
|
Amzallag, L'homme végétal. Pour une autonomie du vivant, 2003, Albin Michel |
|
On lit parfois que la concentration en dioxyde de carbone de l'air, très faible (moins de 0,04%), est un facteur limitant pour la plante. Si l'on augmente sous serre cette concentration, la production primaire (masse de la plante) augmente en effet souvent. Mais cette augmentation est très transitoire. |
|
Les effets d'augmentation de la teneur en dioxyde de carbone de l'air ont été étudiés précisément chez la Jacinthe d'eau (voir références in Amzallag,p 32; article: http://www.plantphysiol.org/ cgi/content/abstract/ 82/2/528 ) et si l'on a effectivement une prolifération de jeunes pousses, le taux de croissance revient, en 4 semaines, à la valeur initiale. Il semblerait que chaque plante possède un taux de croissance propre et que, sous l'action de changements environnementaux, elle puisse s'adapter, plus ou moins rapidement, afin de revenir à un rythme qui lui convienne. Il est faux de dire que le CO2 est un facteur limitant pour la plante (la notion même de facteur limitant est fortement suspecte, voir Amzallag, p 30s). |
|
Par contre, il est nettement plus intéressant de considérer que chaque plante est capable de réguler sa dynamique de croissance, en fonction de son volume racinaire, et par rapport au volume de ses parties aériennes: une régulation morphologique en quelque sorte (voir Amzallag, p33s).
|
||||||||||
| |
|
|
||||||||||||||
|
Capter la lumière
|
|
La captation de l'énergie lumineuse est augmentée par des surfaces que la plante expose aux rayons lumineux (les feuilles sont des fines lames fonctionnelles des deux côtés, mais leur orientation n'est pas aléatoire). |
|
La face inférieure possède plus de stomates que la face supérieure habituellement, ce que l'on relie à une diminution de la transpiration (départ de vapeur d'eau par les stomates) du côté de la face la plus exposée aux rayons lumineux et supposée être la plus chaude. Les cellules photosynthétiques sont situées dans la partie centrale de la feuille (parenchyme chlorophyllien). |
|
Il existe de nombreuses plantes qui ont des parenchymes photosynthétiques au niveau de leur tige. Vous avez tous remarqué qu'une pomme de terre (qui est une tige souterraine chargée de réserves) verdit si elle exposée à la lumière) : les cellules superficielles voient leurs plastes devenir photosynthétiques. |
||||||||||
|
|
|
Comme pour la nutrition carbonée de la plante, je veux ici aussi mettre en garde contre les idées simplistes concernant la prétendue "lutte pour la vie" ou, plus simplement l'idée de compétition, que l'on trouve assez souvent comme explication. La lumière est bien la source d'énergie commune aux autotrophes, mais il est toujours fécond de considérer qu'il existe un véritable dialogue social entre une plante et ses congénères dans un milieu donné, dans l'utilisation des ressources disponibles. Voici une des expériences rapportées par N. Amzallag qui vont dans ce sens : |
|
Portulaca est une petite plante à croissance rapide qui couvre les sols nus. Des chercheurs ont étudié les effets, sur sa croissance, de la présence de voisins dans son environnement (d'après Amzallag, p 37).
1 -panneaux verts de petite
taille entourant les plants (ne
portant pas d'ombre, mais simulant la
présence de jeunes plantules au voisinage
en réfléchissant (faiblement) un
spectre appauvri en radiations rouges) 2 -panneaux peints en noir
et toujours de petite taille (qui
absorbent toute l'étendue du spectre de
la lumière et ne
réfléchissent donc pratiquement
pas la lumière) 3 - exposition à une
lumière enrichie en radiations rouges (qui
sont habituellement absorbées par la
photosynthèse et donc par
d'éventuels voisins) |
||||||||||||
| |
|
|
|
|
||||||||||||
|
Distribuer/stocker les nutriments |
|
La matière organique synthétisée par la plante dans les cellules du parenchyme permet la nutrition de la totalité des cellules ce qui nécessite un système de circulation des nutriments. |
|
C''est la sève élaborée qui distribue principalement le saccharose comme nutriment énergétique. |
|
De nombreuses plantes ont des organes de stockage (tiges modifiées comme les tubercules ou racines modifiées, comme les rhizomes...). |
||||||||||
| |
|
|
|
|
||||||||||||
|
Boire |
|
L'eau est habituellement captée par les racines et conduite dans des "tuyaux" formés de cellules mortes (cellule du xylème qui donne le bois). Les sels minéraux sont aussi captés par les cellules des poils absorbants des racines et conduits avec l'eau en formant la sève brute. |
|
L'eau monte ainsi vers tous les organes de la plante et est évacuée pour la plus grande part au niveau des stomates sous forme gazeuse : c'est l'évapotranspiration. |
|
On notera que de très nombreux arbres ont des racines qui se développent en étroite association avec des champignons (mycorhizes) qui leur fournissent l'eau en échange de différents composés, mais aussi assurent un véritable réseau nutritif entre végétaux d'un même milieu (voir ci-dessous). |
||||||||||
|
Amzallag, L'homme végétal. Pour une autonomie du vivant, 2003, Albin Michel |
|
|
|
Voici une petite expérience (in Amzallag, p 34) qui est intéressante à analyser, toujours dans l'idée d'une autonomie de la plante : |
||||||||||||
|
|
|
Le milieu humide permet de garder les racines vivantes, mais elles ne peuvent absorber d'eau. Si l'on se contente des étapes (1) et (2) de l'expérience (a et b étant à comparer, a étant considérée comme témoin), on est amené à émettre l'hypothèse que, dans le cas b, il y a un déficit hydrique DÛ à un manque d'eau, au niveau des racines du compartiment de gauche, et DONC, à émettre l'hypothèse élargie d'un déficit GLOBAL pour toute la plante. C'est sans aucun doute cette extension du raisonnement à toute la plante qui est erroné. En effet, dans l'expérience complémentaire (b3), ont voit que si l'on coupe les racines vivantes, mais qui ne plongent pas dans l'eau (on pourrait aussi imaginer dans la nature qu'elles se dessèchent naturellement et meurent), on observe une reprise de croissance, montrant ainsi que ce n'était pas un déficit GLOBAL en eau qui était la cause de la baisse de croissance, mais une "décision" adaptative de la plante de modérer sa croissance. Avec les mots d'Amzallag: «la croissance de ces plantes est avant tout régulée par des facteurs internes à la plante, bien en deçà des "capacités de production"». |
||||||||||||||
| |
|
La plante est donc un organisme qui possède deux caractéristiques dynamiques majeures qui lui sont propres. |
||||||||||||||
| |
|
* elle pompe une grande quantité d'eau qui la traverse et qui ressort; il y a un véritable flux hydrique à travers la plante. C'est comme cela que l'on explique que la plante a un tel besoin d'eau. Si elle ne boit pas elle se flétrit, ne peut maintenir sa pression cellulaire et meurt (c'est le moment de réinvestir ce que vous avez vu en première S sur la mérèse et l'auxèse (ancien cours). On pense que c'est aussi ce qui explique l'extraordinaire capacité de la plante à croître: on peut même observer à l'œil nu l'ouverture d'un bourgeon où le déploiement d'une crosse de fougère, ou même la germination de certaines graines. Cette dynamique linéaire orientée est associée à des structures tubulaires ou ramifiées (anastomosées) (poils, tissus conducteurs, tubes avec des orifices, tige, pétiole...). Le flux gazeux est moins net et ne semble en tout cas pas orienté. Les stomates sont très nombreux, mais leur ouverture/fermeture est contrôlée. On ne connaît pas actuellement, me semble-t-il, de système qui entraîne le mouvement des gaz dans la plante. Étant donné la faible densité de l'air par rapport à l'eau, on imagine que les mouvements de diffusion sont suffisants pour assurer le renouvellement de l'air des espaces aérifères. Il y a certainement ici des choses à découvrir. |
|
* elle s'étend verticalement et horizontalement dans l'air pour capter la lumière; elle présente des surfaces plus ou moins planes aux rayons lumineux; on pourrait dire que c'est une dynamique de croissance plane. Cependant, il est bien sûr aussi important de noter qu'à la différence des animaux (ou des unicellulaires autotrophes qui peuvent être flagellés), elle ne se déplace pas (du moins rapidement; car elle est en effet capable, par différents moyens de reproduction sexuée et asexuée, de coloniser un milieu). Ces deux dynamiques - avec leurs structures - sont associées et conduisent à la diversité des formes végétales. La ressemblance des structures végétales avec les fractales (répétition de motifs identiques à différents échelles d'observation) a été maintes fois soulignée. |
||||||||||||
| |
|
|
|
|||||||||||||
|
Extrait du Chapitre 4 d'Esquisse d'une Sémiophysique, René Thom, 1988 (ch4. Embryologie animale) |
|
« Comme l'a déjà
observé Aristote, un caractère important
d'un être vivant est sa divisibilité ; si
l'on peut couper un organisme artificiellement, et
maintenir en vie les fragments simplement en les plongeant
dans un liquide nourricier, alors il n'y a pas «
forme » au sens strict. Tel est le cas des
Végétaux, dont on sait qu'ils peuvent
régénérer à partir d'un cal ne
comptant guère plus que cinq ou six cellules. (...) |
|
La self-similarité qui caractérise les fractals « réguliers » joue donc un rôle essentiel dans la morphologie végétale. Les modèles mathématiques connus proposés pour la forme des plantes comme ceux de la phyllotaxie (la suite de Fibonacci pour les angles des primordiaux sur l'apex de la tige) 1, ou les modèles de Lindenmayer pour la croissance des algues filamenteuses 2, ou encore ceux de Francis Hallé pour la structure des arbres 3 reposent tous sur cette self-similarité - en principe infinie - qui se manifeste par la réinjection d'un champ morphogénétique dans lui-même. De là vient que les botanistes acceptent volontiers l'idée d'une mathématisation sous-jacente à la morphologie végétale, une idée qui choque beaucoup les spécialistes des Animaux. |
|
On observera cependant que même chez les Végétaux, il n'y a pas reproduction indéfinie de la même structure ramifiante. Comme « l'infini n'accède au réel que plongé dans le continu », on se heurte de toute manière à la structure moléculaire de la matière (en fait, bien avant, à la structure cellulaire de toute matière vivante). De plus, l'apparition de la « fonction » biologique se manifeste par l'arrêt du processus génératif : formation de la feuille chez la plante à feuilles, « capteur solaire » transverse aux rayons lumineux, arrêt de la ramification par la sexualité sur l'apex floral (et sur le carpophore des champignons) porteurs du « message » germinal. Ce schéma formel d'une générativité indéfinie stoppée ou modulée par la manifestation de la fonction biologique, se retrouvera chez les Animaux métamériques. Chez les Animaux marins sessiles qui se nourrissent d'un plancton indifférencié, on retrouve presque la morphologie ramifiante du Végétal : il leur suffit de diriger le courant dans un filtre approprié pour capturer leur nourriture (le courant est parfois créé par l'animal lui-même). Ce n'est pas sans raison que ces animaux étaient appelés zoophytes par les naturalistes du XVIIIe siècle. |
||||||||||
| |
|
|
|
|||||||||||||
| |
Toujours à propos des Végétaux, on observera que la plante possède deux structures ramifiantes quasi symétriques par rapport au sol, interface terre-air : l'appareil foliaire et la racine, cette dernière ayant pour fonction l'absorption de l'humidité, alors que les feuilles capturent la lumière. La tige et ses ramifications servent d'intermédiaire, et la circulation à double sens (dans le coeur et l'aubier, xylème et phloème) n'est pas sans rappeler le cycle d'hystérésis attribué par nous au mésoderme animal. Si l'on peut considérer les tissus radiciel et foliaire comme deux sortes d'endoderme, on ne trouvera pas d'homologue végétal à l'ectoderme - sauf peut-être dans les structures florales et ovariennes, qui simulent le transport à longue distance. (Observer à ce propos que les plantes carnivores (Drosera) capturent leurs proies (des insectes) dans des ovaires modifiés.) |
|
Peut-être la racine correspond-elle aussi au besoin de fixation, donc de « capturer l'espace », ce qui n'est possible qu'en phase solide, en terre. On ne peut en tout cas - en considérant la plante - qu'admirer l'intuition d'Aristote en sa théorie des éléments : dans la plante on trouve les quatre éléments : terre, eau, air et feu (la lumière), et la plante est un mixte cinétique de ces quatre éléments, la terre fournissant l'axe de cette rotation, la lumière fournissant l'énergie motrice, et l'eau et l'air étant des fluides vecteurs de l'énergie mise en jeu (Fig. 4.1) .» |
|
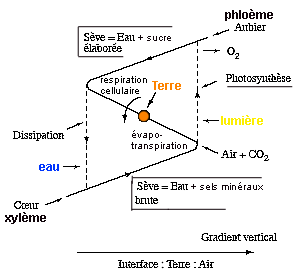 Figure 4.1 de l'ouvrage (très légèrement modifiée) |
|||||||||||
| |
|
|
||||||||||||||
|
Remarque : |
|
Cette intéressante partie est abordée par tous les manuels scolaires (puisque l'on fait extraire les pigments chlorophylliens aux élèves) mais n'est pas à proprement parler dans le programme. Elle n'a pas trop trouvé sa place définitive dans mon cours, je la mets provisoirement ici. TP - Extraction et chromatographie des pigments photosynthétiques d'une feuille - Spectre d'absorption et spectre d'action d'un pigment (Belin p 48-49: Nathan 18-19) |
||||||||||||||
| |
|
Les pigments chlorophylliens NE SONT PAS des substances chimiques magiques ou vivantes; la vie reste une fonction globale; les molécules ont une fonction locale. Elles sont capables, dans des structures vivantes très complexes (que l'on ne sait PAS reproduire in vitro), de transformer de l'énergie lumineuse (arrivée d'un photon) en énergie chimique (dans l'ATP et dans des RH2). Le mécanisme que l'on appelle transduction (du latin trans = à travers et ducere = conduire) est compris actuellement comme un transfert d'énergie d'oxydoréduction sous l'activation de la lumière. |
|
Un complexe photoexcitable à atomes de manganèse (Mn) est tout d'abord responsable de la photo-oxydation de l'eau. Les électrons sont ensuite récupérés successivement par deux gros complexe ou photosystèmes (celui qui porte le n°II est excité principalement à 680 nm et le n°I est excité principalement à 700 nm) comprenant notamment la chlorophylle a qui est le pigment photoexcitable central. Les photosystèmes, en récupérant les électrons en provenance indirecte de l'eau, et à la lumière (sous certaines longueurs d'onde) , passent à un état excité (et réduit) et peuvent alors transférer leurs électrons à des transporteurs d'un potentiel d'oxydoréduction inférieur (l'énergie d'oxydoréduction et donc les électrons sont transférés selon des potentiels décroissants). |
|
Lors de la photolyse de l'eau et lors des transferts d'électrons le long de ce que l'on appelle la chaîne photosynthétique, des molécules d'ADP sont phosphorylées en ATP et donc de l'énergie chimique de liaison est produite. Le dernier accepteur d'électrons est un transporteur (le NADPH). Comme les réactions chimiques se font sans consommation des différentes molécules transporteuses d'électrons (qui sont régénérées) on pourrait tout simplement dire que ce sont des réactions enzymatiques entraînées par la lumière. L'ATP et les NADPH produits lors de la phase lumineuse sont utilisés lors de la phase chimique dans le stroma du chloroplaste. |
||||||||||
| |
|
|
|
|
||||||||||||
| |
|
Je n'ai pas voulu placer cette partie dans la biochimie car il est certain que ce n'est pas la seule approche possible de la photosynthèse. L'approche biochimique est arrivée en quelque sorte à une impasse. La complexité biochimique ne permet pas de comprendre le pourquoi des structures. Par quelles "forces" ou plutôt quelles contraintes les molécules sont-elles regroupées dans des systèmes aussi ordonnés (par exemple le fait que les pigments soient disposés dans ce que l'on appelle des antennes collectrices) ? Il est temps de s'intéresser aux dynamiques pour expliquer les structures. Ce qui n'est pas un mince travail. |
|
Voici quelques pistes de
réflexion (pour des élèves de classes
post-bac). |
||||||||||||
| |
|
|
|
|
||||||||||||
|
3. Être allotrophe |
|
Bases de la classe de seconde à
réinvestir : Bases de la classe de première S à réinvestir : Les enzymes. |
||||||||||||||
| |
|
Un allotrophe se nourrit des autres. Il se nourrit de proies, vivantes (qu'il capture ou sur lesquelles il se fixe) ou mortes (qu'il doit trouver). Il y a toujours une phase d'ingestion-digestion de la nourriture. En dernière analyse le point le plus essentiel est finalement l'état physique de l'aliment: soit un aliment liquide, qu'il faut absorber avec l'eau, soit un aliment solide, qu'il faut alors capturer. |
|
On peut donc résumer les trois actions nutritives des allotrophes: absorber un aliment liquide ou capturer un aliment solide, boire et ventiler. |
|
|||||||||||
| |
|
3.1 Deux protistes mobiles capturant des aliments solides |
||||||||||||||
| |
|
La phagotrophie est la capacité à se nourrir d'éléments nutritifs NON DISSOUS ou SOLIDES (on parle d' "éléments figurés"); ils sont ingérés PAR DES VÉSICULES. |
|
Elle se différencie de l'osmotrophie (très mal nommée car l'osmose désigne les mouvements d'eau) qui consiste à se nourrir d'éléments EN SOLUTION ou LIQUIDES ; ils sont à l'état moléculaire sans que des vésicules capturent les particules alimentaires (c'est le mode de nutrition que l'on attribue au procaryotes allotrophes puisqu'il n'y a pas de mouvement ni de vésicule dans leur cytoplasme). |
|
Il existe deux autres mots qui désignent la
taille des proies PAR RAPPORT à la taille de
l'organisme qui les ingèrent. On distingue ainsi les
macrophages (qui capturent des proies de
grande taille, par rapport à leur propre taille) et
les microphages qui capturent des proies de
très petite taille. Appliquée à une
cellule, la microphagie est l'osmotrophie, mais ce terme
n'est pas employé dans ce sens. |
||||||||||
|
La paramécie |
|
|
|
|
||||||||||||
| |
La paramécie (Paramecium) se déplace activement. Son comportement alimentaire est sans aucun doute la chasse de poursuite ou au moins la captation de particules alimentaires par déplacement actif dans le milieu. |
|
Elle se nourrit de bactéries et de débris organiques qu'elle amène grâce aux battements de nombreux cils (et de cils agglomérés formant une membrane ondulante) dans un entonnoir au fond duquel des vacuoles digestives se forment (par invagination de la membrane plasmique) en englobant les proies (endocytose). Ces vacuoles suivent ensuite un trajet dans le cytoplasme le long duquel elles reçoivent le contenu de petites vésicules contenant des enzymes. |
|
Les éléments nutritifs sont absorbés au cours du transit des vacuoles par de petites vésicules (pinocytose) ou peut-être directement de la vacuole digestive vers le cytoplasme (?). En fin de parcours les vacuoles digestives déversent leur contenu résiduel vers l'extérieur (exocytose) au niveau d'une zone particulière. |
|||||||||||
| |
|
Taille voisine de 250 µm et donc souvent visible à l'œil nu - le pouvoir séparateur de l'œil étant de 0,2 mm; recouverts de cils - pour pouvoir les observer il faut assécher partiellement l'espace entre lame et lamelle afin de les immobiliser ; l'utilisation du rouge neutre dilué, colorant vital (s'il reste à l'extérieur ou tant que le pigment ne se trouve pas dans le cytoplasme), permet d'observer le trajet des particules de pigment depuis le milieu extérieur vers les vacuoles digestives puis vers le cytoplasme... à ce moment a cellule intoxiquée meurt. Dans les macérations végétales en décomposition il est plus fréquent d'observer des Colpodes (Ciliés plus petits d'environ 120 µm et en forme de haricots) et des Vorticelles de taille voisine mais accrochés aux débris végétaux par un pédoncule qui peut être replié en hélice ou détendu et projeter ainsi rapidement la cellule à environ 600µm de son point d'attache; on notera que les mouvements paraissent d'autant plus rapide que le grossissement du microscope nous fait observer des distances très petites qui sont donc parcourues dans des temps très brefs (mais qui ne correspondent pas à des vitesses extrêmes: en effet, notre cerveau est habitué à évaluer des vitesses dans l'espace sensible, et non avec un agrandissement de l'espace; par exemple si le pédoncule se rétracte en 0,5 s en parcourant par exemple 500 µm cela fait une vitesse de déplacement de (500 µm / 0,5 s =) 3,6 m.h-1, ce qui n'a rien d'extraordinaire pour un organisme comme le notre. De la même manière lorsque l'on voit une Paramécie traverser le champ du microscope à toute vitesse sa vitesse de déplacement est de l'ordre de quelques mètres à l'heure tout au plus dans notre espace. La vraie question est de savoir comment évaluer le temps pour un micro-organisme; si l'on se réfère à son échelle de durée de vie (mettons un heure avant de se diviser) cela correspond pour nous à quelques 20 à 80 ans et donc un rapport de 1h/20 à 80 ans = 1h/(20 à 80)x365x24h= 1 à 4.10-6. Si l'on multiplie la distance parcourue par le même facteur cela fait donc, proportionnellement à l'homme une distance non pas de 500µm mais de 0,5 à 2 km, toujours parcourue en 0,5 s. Il y a donc bien une vitesse très élevée. |
||||||||||||||
|
des Foraminifères |
|
Des Foraminifères récoltés sur des algues rouges dans les petites mares entre les rochers de la côte bretonne. |
|
Fixés les foraminifères pratiquent une chasse à l'affût si l'on considère leurs proies mobiles ou de captation de particules sans système spécifique de filtration (mais on peut aussi considérer que les filopodes (fins prolongements cytoplasmiques) anastomosés (réunis par des ponts cytoplasmiques) constituent un filet filtreur). Ils se déplacent aussi avec leurs filopodes. |
|
Ce sont des unicellulaires de quelques centaines de µm dans leur plus grande dimension, entourés d'une "coquille" calcaire appelée test mais perforé. Le cytoplasme des filopodes sort par des pores. Ce sont les filopodes qui captent les particules alimentaires (: autres unicellulaires de petite taille, bactéries, particules organiques...). Des vésicules de phagocytose se forment et amènent les particules au centre du cytoplasme protégé par le test. On pense que des mécanismes similaires à ceux qui interviennent pour la Paramécie ont lieu lors de la digestion. |
||||||||||
|
cours de seconde : |
|
Remarque: Il existe un film extraordinaire que l'on trouve notamment dans le CDRom accompagnant l'édition 2005 de Biologie Moléculaire de la cellule, Alberts et al., Médecine-Sciences-Flammarion. Ce sont Gustavo Pesce et Peter Walter (Institut Médical Howard Hughes - Université de Californie, San Francisco) qui ont filmé les changements de forme du réseau mitochondrial d'une cellule de levure (que l'on classerait maintenant dans les Protistes allotrophes par absorption) à l'aide d'un microscope confocal et d'une sonde fluorescente associée à la séquence signal spécifique des mitochondries que portent certaines protéines impliquées dans le trafic des protéines (ce sont des protéines de type chaperonines qui se fixent sur des récepteurs spécifiques des mitochondries mais d'autres séquences sont dirigées vers d'autres compartiments "membranaires"). J'ai placé dans le cours de seconde une animation extraite des pages de Nunnari. Des images voisines sont disponibles à l'adresse http://www.ucsf.edu/sedat/mito.html (il y a une image gif animée mais les liens de cette page ne fonctionnent pas) et http://www.mcb.ucdavis.edu/faculty-labs/nunnari/ (le film le plus complet étant la page de Jodi Nunnari (Section of Molecular and Cellular Biology - University of California, Davis; mais sont format est "avi" et n'est donc pas lisible sur toutes les machines.... les formats des films et des sons sont un jungle commerciale: bon courage) à l'adresse http://www.mcb.ucdavis.edu/faculty-labs/nunnari/mitoavi.avi). La dynamique mitochondriale, impliquée dans les mécanismes respiratoires cellulaires n'est pas pour moi un problème de chimie (mais avant tout un problème de dynamique d'interface ou membranaire.... (Des chercheurs intéressés peuvent consulter à ce sujet l'article de Yves Bouligand: Remarks on the geometry of micelles, bilayers and cell membranes, Y. BOULIGAND, 1999, Liquid Crystals, vol. 26, n°4, 501-515 qui est juste cité avec deux petites figures extraites de l'article dans mon ancienne page sur les modèles). Le film de la dynamique mitochondriale in vivo est sans aucun doute un argument fort pour montrer combien la dynamique cellulaire, notamment celle des organites, est un point d'entrée passionnant pour la compréhension du fonctionnement cellulaire. |
||||||||||||||
| |
|
3.2 Un champignon : une pelote multicellulaire filamenteuse absorbant des aliments liquides |
||||||||||||||
| |
|
Les mycètes sont des eucaryotes dont les cellules forment habituellement des filaments (mycélium) qui sont appelés hyphes si les cellules sont séparées et siphons ou tubes si les cellules ne sont pas séparées, l'organisme présentant alors un unique cytoplasme avec d'innombrables noyaux: c'est ce que l'on nomme une structure cœnocytique (des cloisons de soutien percées peuvent apparaître). Les filaments peuvent localement s'associer en pelotes ou en files très serrées ou encore en cordons structurés. En coupe, les structures ainsi formées ressemblent à s'y méprendre à certains tissus des plantes, notamment aux racines. Les cellules des mycètes possèdent une paroi et des vacuoles dont la taille augmente avec l'âge des cellules. Aucun plaste n'a jamais été observé. L'appareil de Golgi comme on le connaît manque parfois. Les mitoses, chez les formes les plus complexes, sont atypiques, sans intervention des centrioles et l'enveloppe nucléaire peut persister, totalement ou partiellement. |
|
Si l'on présente habituellement les champignons comme saprophytes (se nourrissant de matière organique en décomposition: du grec sapro = pourri), de très nombreux champignons sont parasites et symbiotes mais au niveau cellulaire la nutrition semble avant tout être sous la dépendance de la présence d'une paroi, comme pour les plantes ou les bactéries. Les aliments doivent traverser la paroi. Cette paroi est de nature variée et surtout complexe (les glucides semblent cependant y dominer), cellulosique (comme chez les plantes et de nombreux protistes), chitineuse (comme la cuticule des arthropodes), kératineuse (comme la peau, la carapace ou les ongles et les poils des vertébrés)... Il ne faut pas croire qu'elle constitue un obstacle ni rigide ni imperméable. C'est plutôt une protection souple et une sorte de milieu intermédiaire. Les enzymes permettant une digestion extracellulaire (exoenzymes) semblent être libérées (par exocytose) dans cette paroi, à proximité directe de la membrane plasmique (et non pas excrétées dans le milieu, loin du filament, habituellement). |
|
Les produits de la digestion sont ensuite absorbés à travers la membrane ("osmotrophie") ou par des vésicules de pinocytose (vésicules d'endocytose de toute petite taille). Certains champignons sont ainsi capables de dégrader la lignine (qui est le composé qui rigidifie les parois des cellules du bois des plantes, c'est un des composés les plus résistants, bien plus résistant que la cellulose). On qualifie les mycètes d'absorbeurs.
Ils développent une surface d'absorption proprement
phénoménale. |
||||||||||
|
associations symbiotiques, parasites ou tout simplement riches relations sociales nutritives |
|
Sans prétendre explorer sérieusement le sujet, je tiens à souligner ici l'importance des relations symbiotiques nutritionnelles tout en ouvrant à des explications plus fondées expérimentalement que les considérations habituelles qui tiennent davantage de l'a priori et de l'anthropomorphisme que de la preuve expérimentale, certes, difficile à atteindre. Presque toutes les plantes terrestres ont
des racines colonisées par des champignons
microscopiques. On a vite fait, en notant les
échanges nutritionnels entre le mycorhize et la
plante (eau, sels minéraux...) d'affirmer que l'on a
une symbiose au sens strict : une association avec
bénéfices réciproques.
Ainsi la plante profiterait du volume de drainage
supplémentaire avec le filament mycorhizien, ainsi
que des éventuels ions; et le mycorhize se servirait
dans les nutriments organiques de la plante. |
|
* Tout d'abord, les plantes ne sont pas limitées dans leur croissance par la disponibilité des ressources, sauf cas de stress hydrique particulier : elles ont une dynamique de croissance qui leur est propre, selon leur espèce, et qui inclut les périodes de disponibilité en eau (l'énergie solaire est habituellement abondante et le carbone alimentaire est en quantité adéquate dans l'air pour leur mécanisme de photosynthèse). La présence d'un mycorhize, ne lui apporte pas forcément grand chose. |
|
* La photosynthèse semble être plutôt stimulée chez des plantes dont les racines sont envahies de mycorhizes (voir références in Amzallag, p 41); ce qui ne conduit pas à une interprétation unique... * Certaines plantes peuvent aller jusqu'à perdre certains métabolismes essentiels : on parle de symbiose obligatoire avec perte de fonctions, comme chez les Orchidées dont les graines ne peuvent germer sans la présence d'un champignon (celui-ci colonise les cellules de l'hôte : c'est un endomycorhize). * Le réseau relationnel entre les
mycorhizes et les plantes associées, parfois
d'espèces très différentes, tend
à élargir notre conception un peu
étroite de la nutrition solitaire optimale de chaque
individu, en la remplaçant par une vision plus
équilibrée où les organismes utilisent
conjointement les mêmes ressources. |
||||||||||
|
Amzallag, L'homme végétal. Pour une autonomie du vivant, 2003, Albin Michel |
|
|
|
|||||||||||||
| |
|
3.3 Des animaux qui absorbent des aliments liquides ou/et qui capturent des aliments solides Si l'on voulait équilibrer le cours il faudrait ici présenter les systèmes ventilatoires, circulatoires, digestifs et excréteurs des animaux. Je n'ose imaginer que les concepteurs du programme considèrent que ce sont des acquis ?! (Je renvoie à d'autres pages générales pour les esprits curieux). |
||||||||||||||
| |
|
Les animaux ont des types d'aliments variés et donc des comportements alimentaires qui ne le sont pas moins. On trouve des filtreurs (les lamellibranches comme la moule qui filtrent par leurs branchies mais aussi la baleine qui récolte le krill avec ses fanons...), des absorbeurs (de l'étoile de mer qui digère ses proies en dévaginant son estomac aux insectes suceurs de sève ou même au colibri qui se nourrit de nectar ou de chair de fruits liquéfiée...), des chasseurs (de la seiche au lion...)... |
|
Remarque: |
||||||||||||
|
4. Le cycle bio-géochimique du carbone |
|
Base de la classe de seconde à
réinvestir : Base de la classe de première S à réinvestir : - éléments de biochimie: glucides, lipides, protides et acides nucléiques |
||||||||||||||
| |
|
Le travail de nutrition regroupe l'ensemble des échanges de matière et d'énergie au sein de l'organisme et entre l'organisme et son milieu. Le carbone étant l'atome spécifique de la matière vivante il est légitime d'essayer de comprendre les relations "nutritives" entre les êtres vivants et les milieux environnants, à l'échelle de la planète, à partir de l'étude des formes de cet atome et des changements entre ces formes. |
|
Les prémisses de l'interprétation de tels cycles pourraient être résumés par l'ancien adage: "rien ne se crée, tout se transforme"; ce qui se traduit ici par des bilans de matière et d'énergie entre réservoirs à l'échelle de la terre. Dans le cycle ci-dessous seuls les bilans de matière sont envisagés. La planète est décomposée en réservoirs (évalués en Gt: une gigatonne = 1 Gt = 109 tonnes de carbone) où telle ou telle forme chimique prédomine et les échanges (notés flux de carbone en Gt par an) sont évalués par de savants (et plus ou moins hypothétiques) modèles. |
|
|||||||||||
|
(d'après Sciences de la Terre et de l'Univers, col, Vuibert, 2000 - Fig 9,2) |
|
|
||||||||||||||
| |
|
Ce cycle sera étudié à nouveau dans le thème de spécialité sur les climats du passé. Nous nous intéressons ici aux formes chimiques du carbone et aux relations entre ces formes. |
|
Remarque: |
||||||||||||
| |
|
|
|
|
||||||||||||
| |
|
Les formes chimiques du carbone sont:
Les relations entre les formes du carbone peuvent s'étudier à deux niveaux:
|
||||||||||||||
|
Objectifs techniques particuliers à cette partie : |
|
- Schématiser le bilan
énergétique et matériel au niveau d'un
organisme de type producteur et de type consommateur ou d'un
écosystème (Belin
p 16, 21) . |
||||||||||||||
| |
|
|
||||||||||||||
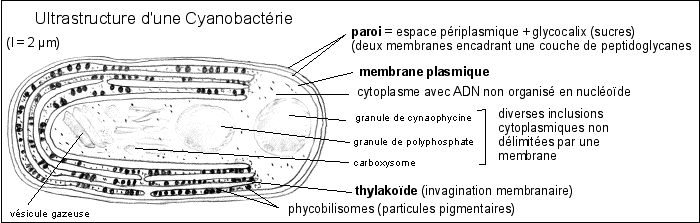
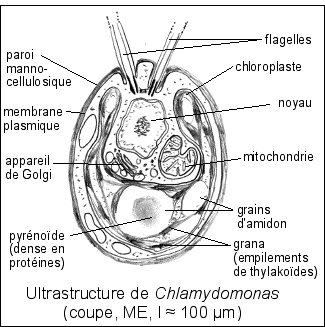
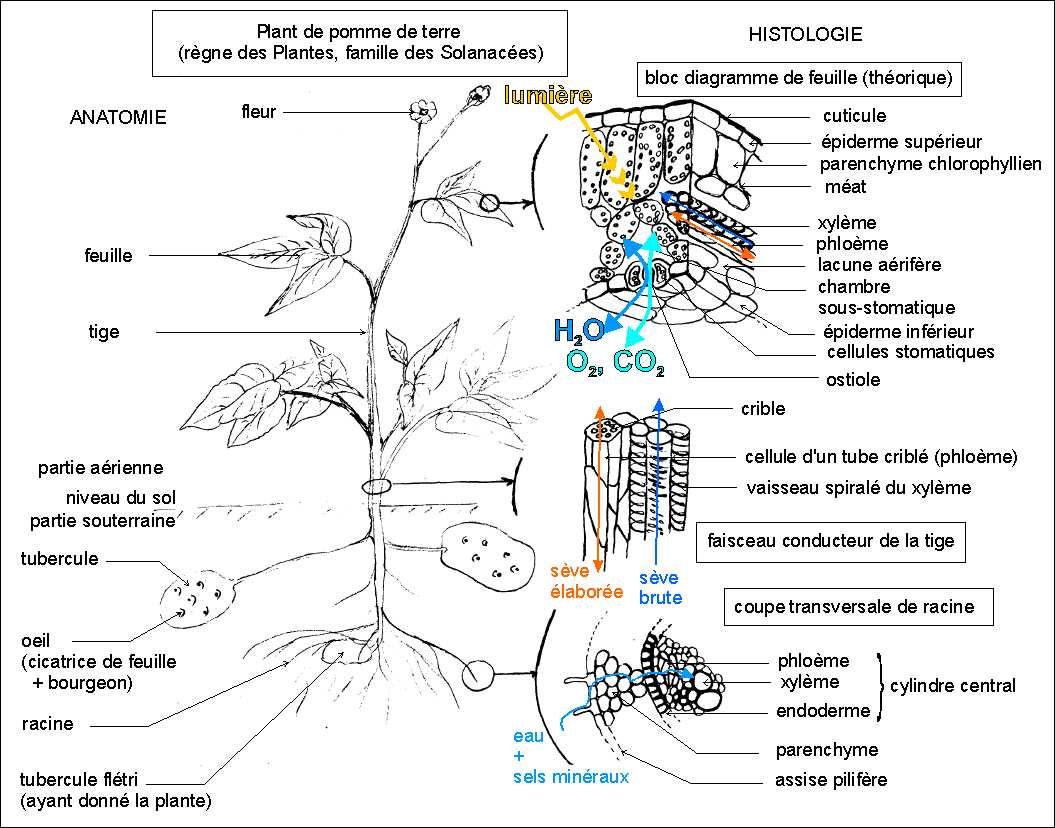


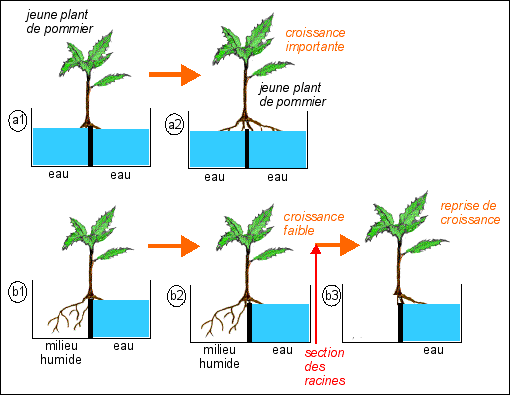



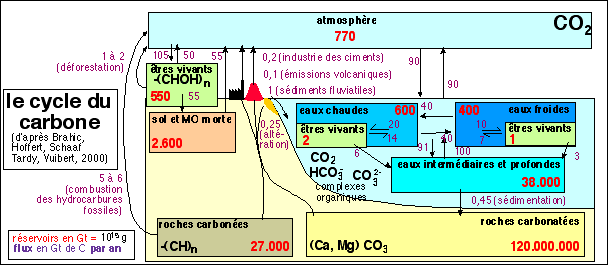
{kind=link}
{kind=link}