|
retour plan du cours, accueil, panorama |
|
Il s'est avéré que, du point de vue pédagogique, il était indispensable de mêler connaissances et réflexion, ce qui rend toujours le cours beaucoup moins structuré mais plus efficace. J'ai du aussi placer ce chapitre en seconde partie étant donné les difficultés qu'il soulevait. |
|
Thème : Corps humain et
santé |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Plan |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
Source: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1. La nutrition des organes lors de l'effort |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Tout le corps participe à l'effort physique. Lors de l'exercice physique ce sont principalement les fonctions de nutrition (donc les échanges de matière et d'énergie) qui sont sollicitées mais les fonctions de relation (communication entre organes, et communication entre l'organisme et son milieu, notamment par les organes des sens) interviennent aussi. |
|
La vie est caractérisée par trois grandes fonctions; nutrition, relation et reproduction (voir partie I). |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
1.1 - L'adaptation de la distribution sanguine lors de l'effort |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Même si de nombreux organes travaillent lors de l'effort, certains sont plus actifs que d'autres. Ils doivent donc maintenir une nutrition importante malgré l'effort. Pour évaluer le travail de chaque organe étudions la distribution sanguine chez un homme au repos et lors d'un effort physique intense. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Objectif méthodologique : analyse d'un document (tableau) |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
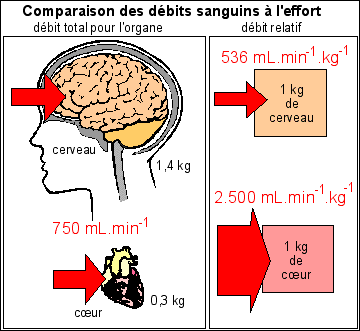
Le débit sanguin au niveau de chaque organe (débit total du sang perfusant l'organe) reflète la distribution de la masse sanguine. Pour pouvoir être comparés utilement entre organes, les débits doivent être corrigés par les débits relatifs (rapportés à la masse de l'organe). |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Explication de la notion de débit relatif (à la masse) |
|
Un débit
est un volume (ou une masse) de fluide qui passe à
travers un organe par unité de temps. Il s'écrit
donc en mL.min-1 par exemple. Pour pouvoir comparer l'irrigation de chaque cellule appartenant à deux organes différents, on divise par la masse de l'organe et l'on compare donc l'irrigation de deux masses identiques d'organes. |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Ici les débits relatifs sont donnés en mL.kg-1.min-1, ce qui veut dire que l'on compare les débits d'1 kg de chaque organe: le cerveau pesant 1,4 kg, son débit relatif est de 750/1,4 mL.kg-1.min-1 = 536 mL.kg-1.min-1; ce qui revient à dire qu' 1kg de cerveau reçoit un débite de 536 mL.min-1. On peut faire de même avec le cœur qui reçoit un débit de 750 mL.kg-1.min-1 à l'effort, mais comme sa masse est de 0,3 kg (300g), son débit relatif, c'est-à-dire correspondant à 1 kg de cœur, est de 750/0,3 mL.kg-1.min-1 = 2.500 mL.kg-1.min-1; ce qui revient à dire que 1 kg de cœur reçoit un débit de 2500 mL.min-1. On peut alors comparer les débits relatifs entre 1 kg de cerveau et 1 kg de cœur à l'effort et l'on trouve une différence de 536 à 2500 mL.min-1, ce qui est bien plus parlant physiologiquement et surtout bien plus éclairant que l'égalité des débits à l'effort (750 mL.min-1) trouvée sur la deuxième ligne du tableau. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Analyse du tableau |
|
Description |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Au repos, les organes recevant le débit le plus faible ne sont pas les reins mais l'abdomen (29 mL.kg-1.min-1 ) et aussi les muscles (41 mL.kg-1.min-1). Le débit maximal passe en fait par les reins (3667 mL.kg-1.min-1); cerveau, cœur et peau ont des débits importants mais intermédiaires entre ceux des reins et des muscles. |
|
À l'effort, l'abdomen voit son débit diminuer (divisé par 2) ainsi que les reins, dont le débit reste cependant très élevé (il est divisé par 2 entre 3667 et 2000 mL.kg-1.min-1). Le débit au niveau du cerveau reste stable. Le débit au niveau du cœur devient maximal (2500 mL.kg-1.min-1, ce qui correspond à un facteur multiplicatif de 3). De très faible, le débit au niveau des muscles devient moyen avec un facteur de multiplication de plus de 10. Le débit au niveau de la peau est aussi multiplié par environ 4x, ce qui fait qu'il est supérieur au débit au niveau du cerveau ou des muscles. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Interprétation - Explication |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
Le débit sanguin est la masse de sang qui irrigue
les cellules d'un organe par unité de temps. Plus le
débit est important plus les cellules peuvent avoir de
nutriments à leur disposition et évacuer leurs
déchets. On établit donc une relation directe
entre activité et débit sanguin. Le débit
étant directement proportionnel à l'activité
(le travail) d'un organe. Les variations de débit sont
considérées comme étant commandées
par l'organe lui-même qui adapte
la vasoconstriction artériolaire et la fermeture des
sphincters à son activité métabolique. |
|
Le rein représente un organe à part car le sang qui perfuse à travers est filtré, ce qui veut dire que ce sang ne sert pas uniquement et directement à la nutrition de l'organe, comme c'est le cas pour les autres débits. Un débit de 1,1 L.min-1 pour les reins signifie que tout le volume de sang de l'homme (~6 L) passe à travers les reins en moins de 6 min. Cependant tout le sang qui perfuse le rein ne passe pas le filtre proprement dit (en donnant l'urine primitive) car seul le plasma (sang débarrassé de ses cellules) passe à hauteur de 180 L par jour, soit 30 fois le volume sanguin tout de même. La majorité de l'eau est réabsorbée (voir cours général pour des précisions). Le rein est l'organe de filtration du sang qui travaille moins à l'effort qu'au repos. |
|
Le débit superficiel au niveau de la peau est élevé même au repos et devient supérieur à celui des muscles lors de l'effort, car la peau effectue un travail spécifique: l'évacuation de l'énergie thermique métabolique (chaleur produite par les réactions chimiques et la contraction musculaire). C'est le réseau artériolaire et capillaire superficiel qui permet l'échange thermique entre le sang et le milieu extérieur. On notera que la transpiration, qui nécessite la consommation d'une énergie de vaporisation (ou chaleur latente de vaporisation), permet aussi grandement de diminuer la température sanguine superficielle (ou pulmonaire) et donc d'évacuer de la chaleur. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
En résumé : |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
1.2 - La nutrition des cellules musculaires est une respiration ou une fermentation |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Les substances organiques
sont des substances carbonées que l'on trouve dans
l'organisme vivant (voir
cours précédent de biochimie) |
Les glucides sont absorbés par l'intestin soit directement sous forme de sucres simples (oses) comme le glucose (à 6 atomes de carbone), soit indirectement sous forme de sucres complexes (osides), comme l'amidon, composé de très nombreuses molécules de glucose, qui seront modifiés en glucose par les cellules intestinales. | La mesure sanguine de la concentration en glucose est apelée glycémie. Elle oscille habituellement entre 0,8 g.L-1 et 1,2 g.L-1, mais localement elle peut atteindre 9,6 g.L-1 (sa moyenne est à 1 g.L-1). | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
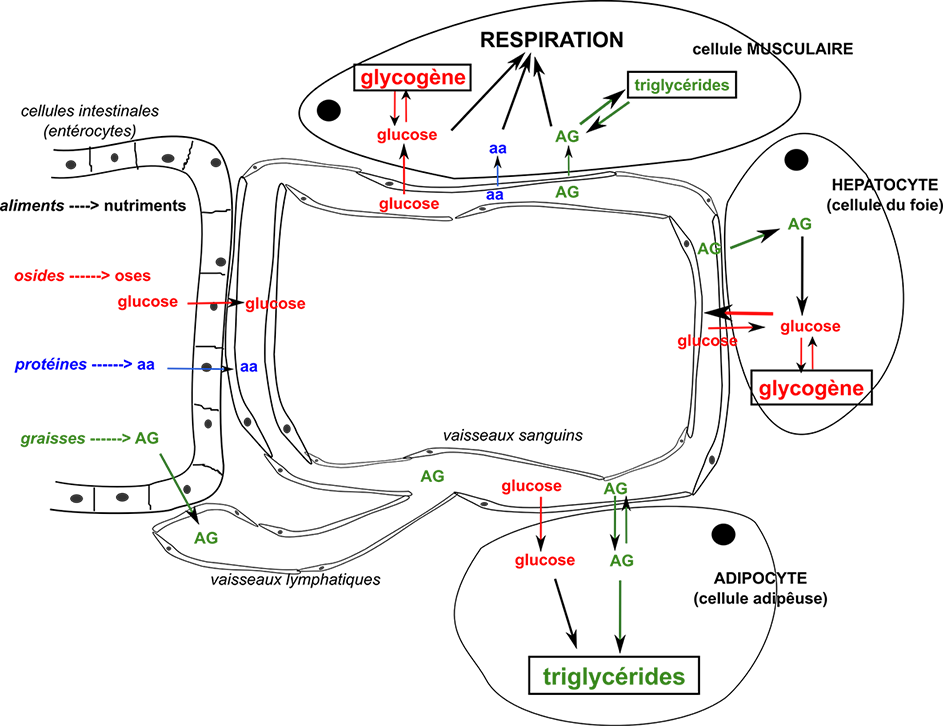 |
Le glucose est (pratiquement)
le seul sucre circulant
dans le sang. Le glucose est stocké
(dans le foie et les muscles) sous forme de glycogène
mais est surtout transformé
en acide gras (AG) par le tissu adipeux qui le stocke alors
sous forme de triglycérides. Le glucose est le seul
nutriment énergétique des cellules nerveuses.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Le glucose (C6H12O6 )
est oxydé en dioxyde de carbone (CO2)
puis il faut du dioxygène (O2) qui est réduit
en eau (H2O), pour fournir beaucoup d'énergie à
la cellule (sous forme de molécules riches en
énergie). |
|
L'équation bilan de la respiration cellulaire est :
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
En absence de dioxygène dans la cellule musculaire (ou en sa présence mais sans que celui-ci n'intervienne), le glucose est fermenté en acide lactique, ce qui n'apporte que très peu d'énergie à la cellule. L'acide lactique doit ensuite être pris par la circulation sanguine pour être dégradé ou retransformé en glucose dans le foie. Remarque : |
|
L'équation bilan de la fermentation lactique (qui est aussi une oxydoréduction) est :
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
2. L'adaptation de l'organisme à l'effort |
||||||||||||||||||||||||||||||||||||||
| |
|
2.1 Mesurer l'effort : quelques paramètres et grandeurs de l'effort et leurs variations lors de l'effort |
||||||||||||||||||||||||||||||||||||
|
Un de mes anciens lycées utilise depuis 2007 le logiciel LoggerPro et les capteurs de la firme Vernier (commercialisé en France par Calibration); nous disposions de 14 PC portables (la région ayant refusé l'équipement Mac) mais je travaille à mon domicile sur Mac... Étant donné la qualité de
ce matériel, même sur PC, je laisse ici quelques
anciennes "Fiches découvertes" sur le logiciel et les
capteurs: Vous pouvez télécharger une version démo EN ANGLAIS (http://perso.orange.fr / calibration/logger.html) du logiciel pour voir les fiches TP |
|
Le terme de paramètre semble avoir
été employé pour la première fois
par Leibniz (dans le
développement d'une famille de courbes). C'est aussi
Leibniz qui, dans un manuscrit de 1684, utilise le mot fonction
dans le sens où on le comprend actuellement (ou
presque): « toute quantité qui varierait d'un
point à un autre d'une courbe » (on lui
doit aussi les mots de variable et constante).
(d'après M. Kline, Mathematical Thought from Ancient to
Modern Times, New York, Oxford University Press, 1972, p.
339-340.; cité dans les notes de Paraboles et
catastrophes, Réné Thom) Pour essayer de classer les paramètres nous verrons d'abord ceux de que l'on peut mesurer de l'extérieur et ensuite les paramètres plus complexes soit parce qu'on ne peut pas les mesurer de l'extérieur soit parcequ'ils sont des fonctions d'autres paramètres. |
|
Remarque: |
||||||||||||||||||||||||||||||||||
|
2.1.1 Les paramètres ventilatoires et respiratoires |
|
Remarque: |
||||||||||||||||||||||||||||||||||||
|
fréquence ventilatoire |
|
Un cycle ventilatoire comprend une inspiration (active) et une expiration (passive). Le volume courant est le volume d'air renouvelé dans les poumons à chaque cycle ventilatoire. Lors d'exercices physiques un volume supplémentaire peut être mobilisé (venant du volume résiduel et de l'espace mort). |
|
|||||||||||||||||||||||||||||||||||
|
pulmonaire total (6 L) |
|
Les volumes pulmonaires de la colonne de gauche sont estimés AU REPOS. Il est essentiel de noter qu'au cours d'un exercice physique l'espace mort diminue pour atteindre moins de 1/20ème du volume courant et devient donc négligeable. AU REPOS, à chaque inspiration, 0,35 L (volume renouvelé) d'air "frais" se mélangent aux 1,65 L (1,5 L de réserve expiratoire + 0,15 L de l'espace mort) et donc il n'y a un renouvellement que du cinquième de l'air pulmonaire. Lors d'un exercice le renouvellement atteint près de 0,5 L sur 1,5 L soit 1/3. |
||||||||||||||||||||||||||||||||||||
|
Fiche TP (nécessite LoggerPro) volumes ventilatoires (vide).cmbl |
|
Le rythme ventilatoire, ou plutôt la fréquence ventilatoire, nombre de cycles ventilatoires par unité de temps, augmente bien sûr avec l'activité physique. (Rythme est un mot très général pour caractériser des phénomènes périodiques ou même des mouvements temporels très divers). Le paramètre le plus utilisé pour comparer la ventilation chez différents organismes est apellé "volume" de ventilation qui est égal au produit de la fréquence ventilatoire avec le volume courant (c'est donc en fait le débit ventilatoire encore apellé "ventilation"). Le débit ventilatoire (DV ou ventilation (noté V° voir ci-contre-->), exprimé en L.min-1 ou L/min) est égal au volume courant (VC) multiplié par le fréquence ventilatoire (FV) : |
|
Exercices et étude de documents (Belin p 121 et p160-163, Bordas p 154-155)
Le débit est
représenté ici par V° mais il devrait
être noté avec un V majuscule surmonté
d'un point : |
||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||
|
|
|
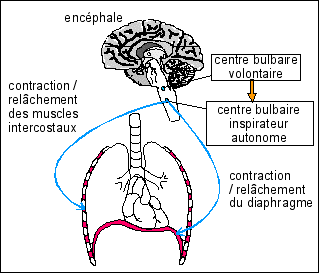
La ventilation est un phénomène automatique
dont l'origine est nerveuse. La fréquence ventilatoire
est sous la dépendance d'un centre bulbaire
respiratoire relié par des nerfs aux principaux muscles
de l'inspiration: les muscles intercostaux et
le diaphragme. Schématiquement, on pense
que la seule phase active ventilatoire est
l'inspiration qui est involontaire ;
l'expiration étant strictement passive
(décontraction de tous les muscles). C'est donc un
tonus
ventilatoire (une stimulation
nerveuse permanente, mais rythmique, des muscles intercostaux
et du diaphragme). * Dans une boucle nerveuse, voir plus-bas, « efférent » désigne les voies allant du centre nerveux aux organes périphériques effecteurs et « afférent » désigne les voies venant des récepteurs périphériques et gagnant les centres nerveux. |
||||||||||||||||||||||||||||||||||||
|
En 2005 le "Prix du Ministère" de La Recherche a été attribué à Bernard Sapoval, Maddalena Felici , Marcel Filoche , Benjamin Mauroy et Ewald Rudolf Weibel pour leurs travaux sur la "Physique de la respiration chez les mammifères : efficacité ou robustesse" (http://www.larecherche.fr/ html / LPLR/ laureats/ 2005.jsp# ministere: je conseille de lire la présentation faite lors de la remise du prix: http://www.larecherche.fr / html /2008/pdf/ sapoval.pdf) On peut aussi voir la conférence de Bernard Sapoval (Laboratoire de physique de la matière condensée, Ecole Polytechnique: http://pmc.polytechnique.fr/ groupes /irregularite/activites/ hydropoumon/ hydropoumon.htm ) : Physique de la respiration accessible sur le site de l'ENS, diffusion des savoirs (http://www.diffusion.ens.fr / index.php?res=conf&idconf=230), et d'autre part, pour une approche plus biologique orientée vers le développement, je conseille les travaux de Vincent Fleury et notamment son article dans Génétiquement indéterminé, 2007, Quæ (Morphogenèse des structures arborisées et conditions physico-chimiques d'une croissance biologique auto-organisée, pp 111-140). |
|
Propos de biophysiciens Nos connaissances sur la dynamique de la ventilation sont loin d'être définitives et satisfaisantes. Depuis la fin du XXème siècle les physiciens et les mathématiciens, associés aux médecins et physiologistes, s'efforcent de modéliser les voies aériennes. Il y a en effet de quoi être émerveillé devant l'extraordinaire surface pulmonaire (environ 100-140 m2) et le nombre d'acini (environ 30.000) qui sont considérés comme autant de petites pompes placées à l'extrémité de ramifications dichotomiques (voir respiration sur le site associé). Le nombre d'alvéoles étant estimé à 300 millions. On considère qu'à partir du niveau de l'acinus (ordre supérieur à 16, c'est-à-dire ordre à partir duquel on voit des alvéoles le long des conduits) l'air n'est plus mis en mouvement par les cycles ventilatoires (les échanges de gaz entre le sang et l'air alvéolaire sont modélisés par les lois de la diffusion; ils sont améliorés chez l'adulte lors d'un exercice (car les acini sont plus grands que l'efficacité maximale calculée l'exige); par contre chez le nourisson où lesacini sont petits, il n'existe aucune amélioration possible... le nourisson qui fait de l'exercice (vocal !) se cyanose...). Actuellement on ne sait modéliser que le flux d'air dans un réseau de second ordre (1 branche principale, 2 branches de 1er ordre, 4 branches de 2ème ordre) alors que l'arbre bronchique s'étend sur 24 ordres (en comptant la trachée d'ordre 0). Les résultats de la mécanique des fluides nous indiquent que le système est extrêmement sensible à la géométrie (angle entre les branches, longueur et diamètre des branches...; ce point permet de mettre en évidence qu'étant donné cette sensibilité très forte et le caractère fragile du système optimal, il est indispensable que l'organisme ait un système de contrôle de la dynamique de la géométrie pulmonaire -par exemple du diamètre des bronchioles , par la quantité de liquide ou par contraction ...). Un autre résultat original est que l'inspiration et l'expiration ne sont pas des régimes homogènes symétriques. Les notions de phases passive/active sont certainement à remettre en cause. Pour ce qui est du contrôle, voir la troisième partie de cette page. |
||||||||||||||||||||||||||||||||||||
|
|
|
|
||||||||||||||||||||||||||||||||||||
| |
On mesure la concentration en dioxygène et en dioxyde de carbone de l'air expiré (en % et en ppm) et en en déduit la consommation cumulée de dioxygène et le rejet cumulé de dioxyde de carbone (en L.h-1 ou mL.mn-1). |
|||||||||||||||||||||||||||||||||||||
|
Fiche TP (nécessite LoggerPro) consommation dioxygène dioxyde carbone effort (vide).cmbl |
|
L'intensité respiratoire (IR) est le
volume de dioxygène consommé par unité de
temps et de masse (en L.min-1kg-1). |
|
Après un effort l'organisme continue de consommer plus de dioxygène qu'au repos. Cette surconsommation est appelée "dette en oxygène" et correspond probablement à l'inertie du système métabolique et à la reconstitution des réserves énergétiques de l'organisme. |
|
V°O2max est un excellent indicateur de la capacité d'effort d'un sportif J'ai jusqu'à maintenant parlé façon erronée du VO2 max comme étant le volume d'O2 consommé (en L) lors d'un effort physique d'intensité maximale. Mais ce n'est pas un volume mais un débit - noté avec un V majuscule surmonté d'un point ( V°O2max varie selon l'entraînement et les individus. (Belin p 126-127, Bordas p 119, 131) |
||||||||||||||||||||||||||||||||
| |
|
Remarque: L'IR n'est pas un bon paramètre pour mesurer les caractéristiques de l'effort ENTRE INDIVIDUS car il varie trop en fonction des caractéristiques individuelles, notamment l'état physiologique. Mais il est pertinent pour faire des comparaisons chez un même individu (... mais dans ce cas pourquoi diviser par la masse si ce n'est pour comparer avec d'autres ?). C'est un paramètre qui est préférentiellement utilisé pour des bilans nutritionnels et qui est alors couplé avec le quotient respiratoire (QR = rapport du rejet de dioxyde de carbone et de la consommation de dioxygène - voir cours de terminale spécialité). (Bordas p190 doc2) |
|
|||||||||||||||||||||||||||||||||||
| |
|
|
|
|
||||||||||||||||||||||||||||||||||
|
pressions partielles sanguines (différence artério-veineuse) |
|
|
|
Étude de documents: (Belin p 142-143 et p 147, Bordas p 128-129 et 130-131)
ATTENTION Au niveau des poumons, au contraire des autres organes, le sang se recharge en dioxygène. Comme on est très proche du cœur on garde la même terminologie pour désigner les vaisseaux irriguant les poumons: les artères pulmonaires, venant du cœur apportent un sang faiblement chargé en dioxygène (bleu) alors que les veines pulmonaires ramènent au cœur un sang chargé en dioxygène (rouge). La couleur du sang dépend de sa charge en dioxygène par changement direct de la couleur de l'hémoglobine. Lors de la fixation du dioxygène sur l'atome de Fe contenu dans une chaîne de l'hémoglobine (la protéine qui transporte le dioxygène), la couleur devient rouge |
||||||||||||||||||||||||||||||||||
|
débit respiratoire |
|
|
|
Étude de documents: (Belin p 147, Bordas p 123) |
||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||
|
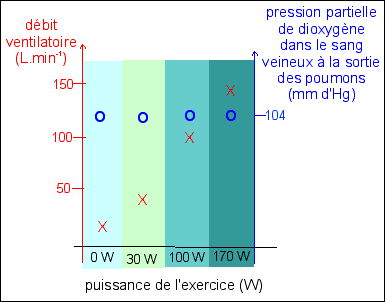
1er exemple :
On ne connaît pas l'âge, le sexe, le type d'éffort, la durée de l'effort, les conditions individuelles physiologiques... d'après Bordas p 131 (ou p 125 de la nouvelle édition, CORRIGÉ) |
|
Description Comme la variation de la puissance de l'exercice n'est
pas régulière, on ne peut pas vraiment tracer
une courbe - dans ce graphique - qui exprime une relation
entre débit ventilatoire et puissance de l'exercice. Il
y a peut-être une fonction mais elle n'est pas
représentable sur ce graphique (voir exemple
2). On peut
cependant dire que le débit ventilatoire augmente avec
la puissance de l'effort. |
||||||||||||||||||||||||||||||||||||
| |
|
Explication Nous verrons dans le dernier chapitre comment exprimer l'idée, qui est ici patente : la nutrition en dioxygène des cellules est STABLE, malgré les variations de l'effort.et du débit ventilatoire. |
||||||||||||||||||||||||||||||||||||
|
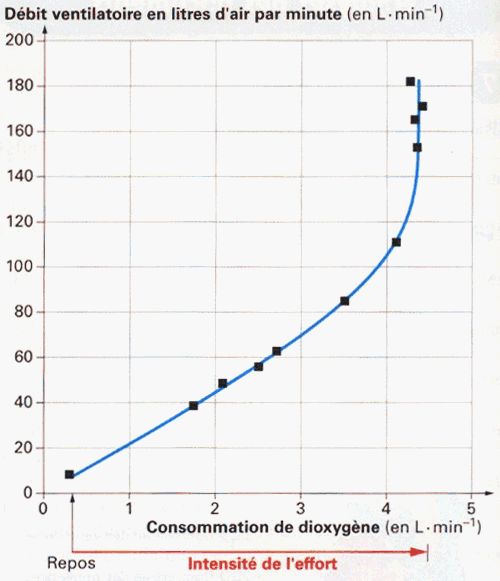
Certains manuels scolaires fournissent une courbe du débit ventilatoire en fonction de la puissance de l'effort (ou/et de la consommation instantanée en dioxygène). |
|
Deux éléments: - l'axe des abscisses superpose deux
paramètres. On peut donc penser qu'ils sont
reliés. C'est vrai pour un exercice de
faible intensité (la consommation de
dioxygène par unité de temps (ou
débit respiratoire = R°) augmente
proportionnellement avec l'intensité de
l'effort : il y a une relation de
linéarité : R° = c . W
(où c = constante, et W l'intensité
de l'effort (puissance) en Watts). Mais, pour des
exercices plus intense (vers 300 W), la VO°2max
est atteinte et le R° ne peut plus augmenter. - On observe que pour des valeurs
élevées du débit ventilatoire
(V°) (pour lesquelles l'intensité de
l'effort ne peut plus être
augmentée), le V° continue d'augmenter
sans lien avec l'effort - ou plutôt sans
lien avec la consommation de dioxygène qui
est alors maximale -, on a donc une déconnexion
du système ventilatoire par rapport au
système respiratoire en lien avec l'effort.
(Pour x constant, y varie sans relation avec x ;
ici V° et W n'ont plus de lien entre eux). |
||||||||||||||||||||||||||||||||||||
|
Physiologie du sport et de l'exercice, Wilmore et Costill, De Boeck, 2006, p 222s |
|
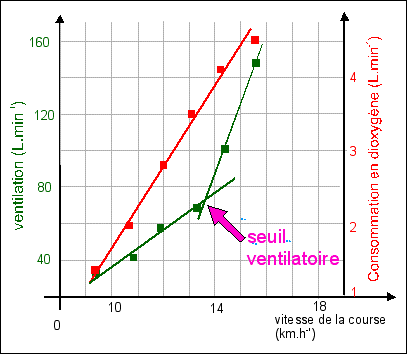
graphique 1
|
|
Les physiologistes utilisent aussi : - l'équivalent respiratoire en dioxygène = rapport entre le volume d'air ventilé (VV) et le volume de dioxygène consommé (VO2), les deux volumes étant estimés pour une durée donnée (au repos ou lors d'un exercice). - l'équivalent respiratoire en dioxyde de carbone = rapport entre le volume d'air ventilé (VV) et le volume de dioxyde de carbone produit (VCO2), les deux volumes étant estimés habituellement pour une durée d'une minute. On note qu'il existe un seuil ventilatoire lorsque l'on réalise un effort d'intensité croissante: seuil d'effort à partir duquel l'augmentation de la ventilation est supérieure à celle de la consommation en dioxygène (graphique 1). Dans le graphique 2 où les deux équivalents respiratoires sont indiqués pour chaque intensité d'effort, on retrouve le seuil ventilatoire qui reste proportionnel au CO2 produit (la courbe de régression est une droite de faible pente sans seuil) mais pas à l'O2 consommé (puisqu'on doit envisager deux droites de régression séparées par un seuil), ce qui tend à faire penser que c'est bien pour éliminer le CO2 produit que la ventilation augmente. Comme on sait que la production de CO2 augmente de façon importante lors de la fermentation lactique (on parle de seuil lactique à partir duquel la lactémie (concentration sanguine d'acide lactique) s'élève au-dessus des valeurs de repos), il est tentant d'identifier seuil ventilatoire et seuil lactique. Cependant cette identification reste délicate car chez des sujets présentant une pathologie les rendant incapables d'augmenter l'acide lactique sanguin, le seuil ventilatoire persiste. Il existe donc probablement d'autres facteurs que le pH (dont est responsable pour une bonne part l'acide lactique et pour une encore plus grande part le CO2 sanguin dissous, voir ci-dessous) et la pCO2 sanguine pour expliquer cette augmentation brusque de la ventilation à partir d'un certain niveau d'effort. |
||||||||||||||||||||||||||||||||||
| |
|
Le contrôle de la ventilation est très sensible à la pCO2 du sang mais très peu à la pO2 |
||||||||||||||||||||||||||||||||||||
|
Physiologie animale, Adaptation et milieux de vie, Schmidt-Nielsen, Dunod, 1998, pp33s
page spéciale sur les carbonates sur le site associé |
|
Il est très facile de mettre en évidence la
sensibilité des Mammifères au CO2 de
l'air pulmonaire en ajoutant du CO2 dans l'air
inspiré. Pour une concentration de 2,5% de CO2
(par rapport au 0,04% habituels)
la ventilation est presque doublée. A plus fortes
concentrations le CO2 a un effet narcotique. Le
dioxygène a également un effet sur la
ventilation mais il est beaucoup plus faible (pour une
variation de 21% à 18,5%, l'effet est quasiment
indécelable). |
|
On pense donc que le centre ventilatoire bulbaire est
directement sensible à la pCO2 du sang (et
du liquide céphalo-rachidien) et non à la pO2. |
||||||||||||||||||||||||||||||||||
| |
|
On explique ainsi les inspirations profondes que les nageurs en apnée pratiquent avant la plongée, non pas par une charge supplémentaire de dioxygène dans le sang, mais par une élimination supérieure du CO2 sanguin, ce qui a pour effet de diminuer le réflexe ventilatoire (dont le centre est au niveau du bulbe) et augmenter ainsi la durée de l'apnée. En effet la pCO2 reste alors suffisamment faible pendant la plongée pour ne pas déclencher le réflexe d'inspiration. Il va sans dire que cette pratique est fort dangereuse, car une trop forte baisse de la pO2 sanguine peut provoquer une perte de connaissance conduisant ainsi à la noyade. |
||||||||||||||||||||||||||||||||||||
|
2.1.2 - Les paramètres cardiaques |
|
|
|
|
||||||||||||||||||||||||||||||||||
|
fréquence cardiaque |
|
Le rythme cardiaque ou plutôt la fréquence cardiaque (nombre de battements cardiaques par unité de temps) augmente juste après le début de l'effort mais reste à un niveau élevé après la fin de l'effort pendant quelques minutes (c'est la récupération). |
|
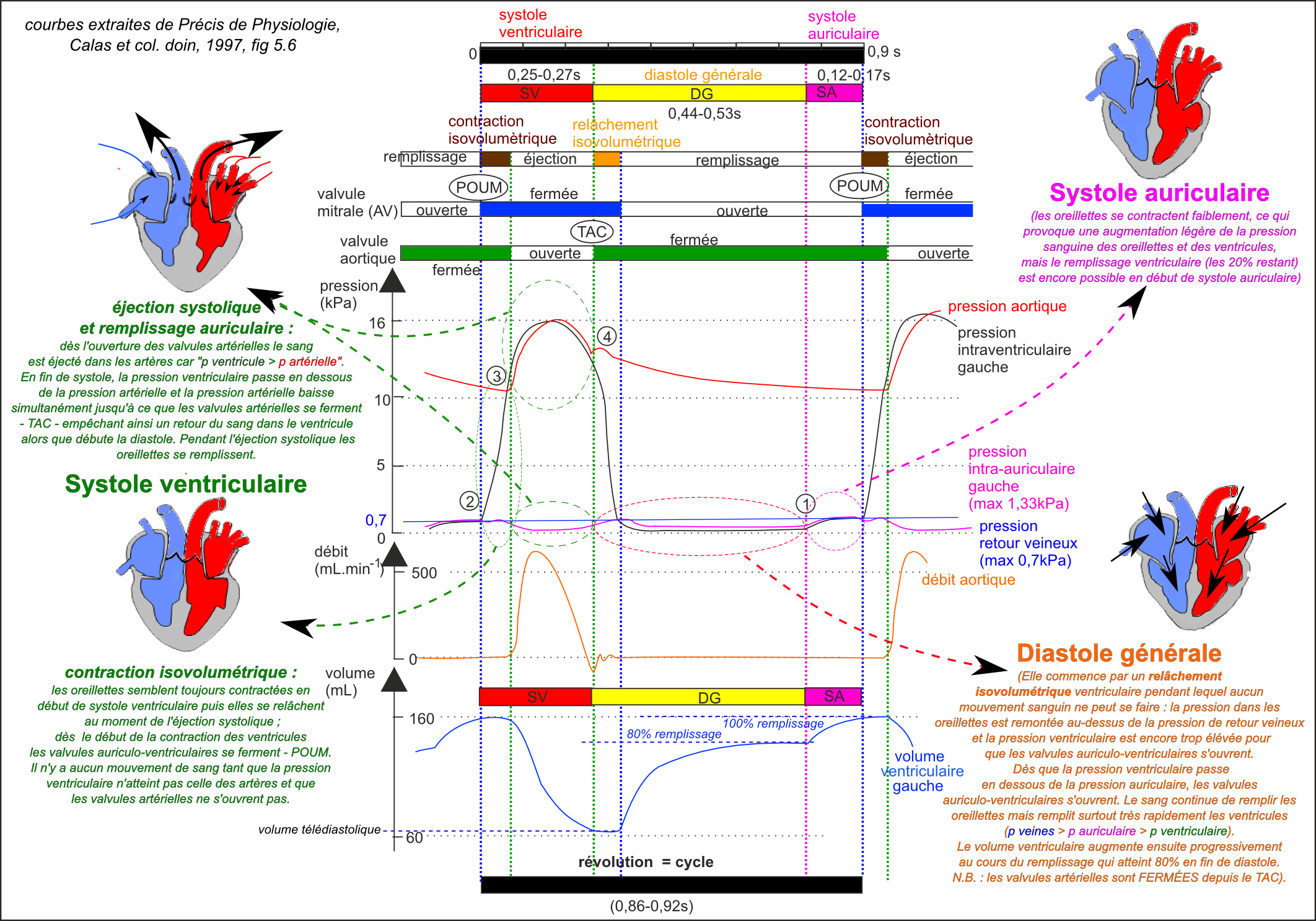
Exercices et études de documents: la révolution cardiaque (Belin p 140, Bordas p 126) et son contrôle nerveux (Belin p158 -159, Bordas p 152-153; et exercice n° 1 p 167) |
|
|
||||||||||||||||||||||||||||||||
|
|
|
Le cœur isolé de ses nerfs bat à un rythme autonome élevé (± 89 battements.min-1). L'origine de l'autonomie est musculaire, interne aux cellules cardiaques (myocytes) qui, dès les premiers mois de la vie embryonnaire, sont capables de se contracter rythmiquement. Certaines des cellules musculaires cardiaques restent à l'état embryonnaire et forment le tissu nodal à l'origine de la contraction de l'ensemble du cœur. Les autres cellules cardiaques associées à des cellules conjonctives perdent leur rythme autonome, mais se contractent rythmiquement sous l'impulsion électrique donnée par le tissu nodal. Remarque : Le rythme cardiaque est aussi sensible à certaines substances chimiques circulantes (médiateurs endocrines ou hormones) plus ou moins identiques à celles libérées par les terminaisons nerveuses au niveau du tissu nodal (neuromédiateurs). |
|
|||||||||||||||||||||||||||||||||||
|
Un super moyen
mnémotechnique pour retenir le rôle du
système nerveux végétatif sur la
fréquence cardiaque : |
|
En plus de l'autonomie musculaire il existe une
autonomie nerveuse : le bulbe stimule - et inhibe
à la fois - en permanence et rythmiquement le
cœur : c'est un tonus* nerveux, comme
pour les muscles ventilatoires (voir
ci-dessus).. (Belin p 121, Bordas p 122). |
|
Remarques: ** Dans une boucle nerveuse, voir plus-bas, « efférent » désigne les voies allant du centre nerveux aux organes périphériques effecteurs et « afférent » désigne les voies venant des récepteurs périphériques et gagnant les centres nerveux. |
||||||||||||||||||||||||||||||||||
|
Exercice
d'application: |
|
|
|
Le VES est assez peu adaptable lors de l'effort mais par contre l'entraînement peut augmenter considérablement sa valeur. La fréquence cardiaque est par contre un paramètre TRÈS ADAPTABLE. La fréquence cardiaque maximale autorisée lors des test d'effort (mesure du V°O2max) est de 200 min-1 (ou parfois 220-âge (en années) min-1). |
||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||
|
* on aimerait savoir sur
quel types de sujets ces mesures ont été
faites, avec quel type d'effort et avec quelle incertitude
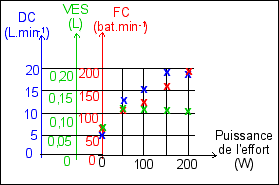
ces résultats sont donnés Le paramètre "puissance de l'effort", exprimé en watts (W) ne correspond pas à une variation continue (courbe) mais à une série de 5 mesures: - au repos (puissance de l'effort nulle: ce qui bien évidemment ne correspond pas à un effort global de l'organisme nul mais uniquement à un effort supplémentaire nul par rapport au repos). - pour des puissances de 50W, 100W, 150W et 200W qui sont donc des puissances IMPOSÉES. Il y a donc une variation DISCONTINUE de la puissance de l'effort. IL NE FAUT DONC PAS TRACER DE COURBE (encore moins des segments de droite) ENTRE CES POINTS EXPÉRIMENTAUX. -------> |
|
Rien ne permet de dire que la variation de débit cardiaque, de fréquence cardiaque ou de volume d'éjection systolique, est une variation régulière en fonction de l'augmentation de la puissance de l'effort. C'est-à-dire que la pente de la courbe (qu'il ne faut pas tracer) réunissant les points de mesures pour l'une des trois grandeurs mesurées (DC, VES ou FC), n'est pas significative. Elle ne permet que de visualiser des écarts de valeurs entre deux points de mesure. Ce que l'on compare ce sont les valeurs relatives des 3 grandeurs pour chaque puissance d'effort; ce ne sont pas les variations, en considérant de façon fausse que le temps est représenté en abscisses. Entre 0 et 50W par exemple la variation de la fréquence cardiaque n'est pas "moins forte" que celle du débit cardiaque. Ce ne sont pas les mêmes unités et les variations ne sont donc pas comparables, numériquement. Si l'on avait une évolution en fonction du temps on pourrait penser que l'on mesure la vitesse d'un phénomène et donc que les pentes seraient comparables. |
||||||||||||||||||||||||||||||||||||
| |
|
Le DC est très nettement augmenté lors d'un effort physique. Il augmente d'autant plus que l'effort physique est important mais pas au delà d'un certain seuil, probablement situé entre 150 et 200W pour ce sujet. Le DC est un paramètre qui augmente lors de l'effort mais qui semble limité pour des efforts importants. Il est donc à la fois adaptable et limité. En fait il est relié aux deux paramètres précédents par une relation simple (DC = FC x VES). Ce paramètre visualise bien l'adaptation de l'organisme à l'effort pour de faibles valeurs de la puissance de l'effort (<150W) mais aussi, pour des efforts plus importants, la contrainte physiologique du VES qui mesure en fait l'efficacité du remplissage du cœur. |
|
Le VES est augmenté lors de l'effort mais d'une valeur qui semble quasiment indépendante la puissance de l'effort (elle aurait même tendance à diminuer par rapport à cette valeur pour l'effort le plus important). Le VES est le paramètre le plus stable à l'effort, si ce n'était la différence entre la valeur au repos et celle lors des efforts entre 50 et 200w, toutes voisines. Il est donc peu adaptable. Son augmentation fait appel à des paramètres comme la pression de retour veineux et certains paramètres de la contraction (durée et amplitude). |
|
La FC augmente avec la puissance de l'effort. La FC est le paramètre dont l'augmentation au cours de l'effort semble être la plus "régulière" (même si ce terme est faux - voir ci-dessus). On aimerait trouver une fonction physiologique qui soit associée à cette grandeur et qui soit maintenue stable au cours de l'effort. Mais c'est au contraire l'effort, imposé, qui oblige l'organisme à adapter sa FC à la puissance de l'effort. La FC est donc au contraire une grandeur qui visualise la réponse de l'organisme à la demande accrue de nutriments. La FC est donc hautement adaptable à l'effort. Les mécanismes d'adaptation rapide peuvent s'expliquer classiquement par une commande nerveuse (S et PS par exemple). Les variations de l'activité nerveuse pouvant être d'origine volontaire (préparation et réalisation consciente de l'effort) ou involontaire (adaptation du système nerveux à des informations en provenance de récepteurs internes à l'organisme mesurent des paramètres nutritionnels - comme la teneur en dioxygène ou en glucose du sang - ce que l'on appelle un réflexe de régulation). |
||||||||||||||||||||||||||||||||
| |
|
Finalement, que nous apprennent ces
graphes ? |
||||||||||||||||||||||||||||||||||||
|
2.1.3 - Les paramètres vasculaires |
|
|
|
Vasculaire vient du latin vascula = petit vase et désigne tout ce qui a trait aux vaisseaux, aussi bien sanguins (veineux et artériels) que lymphatiques. On utilise aussi le préfixe "vaso" pour désigner les vaisseaux. |
||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||
| |
|
La résistance périphérique
est la force qui s'oppose à l'écoulement du sang
dans le réseau artériolaire et capillaire. Elle
dépend de la constriction des artérioles et de
la fermeture des sphincters (muscles circulaires pouvant
fermer un conduit en se contractant). |
|
 Structure transversale simplifiée des principaux vaisseaux |
||||||||||||||||||||||||||||||||||
| |
La vasoconstriction ou la vasodilatation sont sous contrôle nerveux et hormonal (par des substances chimiques ou médiateurs). Si le système nerveux exerce un CONTRÔLE CENTRAL, des médiateurs peuvent être libérés par les tissus irrigués lors de leur fonctionnement : c'est un CONTRÔLE LOCAL. Chaque muscle, par exemple, pouvant lui-même adapter son irrigation à son activité. |
|
Remarque : |
|
Une hormone (ou médiateur
endocrine) est une substance chimique
sécrétée par des cellules
spécialisées (dites endocrines),
transportée par le sang, et qui agit sur
des cellules cibles particulières (pourvues
de récepteurs à cette hormone). (Belin p 145 et 147; Bordas exercice n°2 p 143; sphincters à ajouter au schéma p 115). |
|||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||
|
Pour des données
classiques |
|
La physique de la géométrie de l'arborisation artério-veineuse est aussi fort étudiée, comme celle de l'arbre bronchique. D'une part la mise en place lors du développement (croissance dendritique; voir par exemple les travaux de Vincent Fleury (article dans Génétiquement indéterminé, 2007, Quæ (Morphogenèse des structures arborisées et conditions physico-chimiques d'une croissance biologique auto-organisée, pp 111-140)) et d'autre part la physique de l'écoulement du fluide sanguin dans cet incroyable réseau (27 ordres de grandeur entre l'aorte et les plus fins capillaires).
il reste 2500 mL dans le
cœur... |
||||||||||||||||||||||||||||||||||||
| |
Le terme pression artérielle désigne couramment la pression moyenne (au-delà de la pression atmosphérique) qui règne dans les grosses artères et MESURÉE à l'aide d'un sphygmomanomètre (ou brassard gonflable associé à un manomètre et un stéthoscope). On mesure bien évidemment soit la pression au bras en position couchée, soit la pression au bras en position debout ou assise, à hauteur du cœur, de façon à s'affranchir de l'énergie gravitaire. On donne deux valeurs: la valeur la plus élevée est la pression systolique (pas) et la pression la moins élevée: la pression diastolique (pad). |
|
On parle d'hypertension lorsque la pression
artérielle se situe ne permanence au-dessus* des
valeurs normales.
* il n'y a pas de maladie de l'hypotension (valeur située en permanence en dessous des valeurs normales): l'hypotension n'est pas un état stable à la différence de l'hypertension. |
|||||||||||||||||||||||||||||||||||
|
Analyse de documents : Analyse: Interprétation : |
|
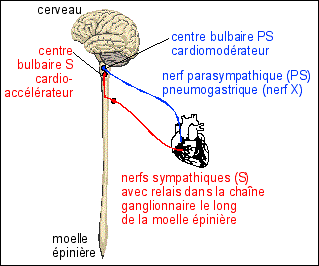
Mise en évidence de la relation
entre la pa, la FC et les systèmes S et PS chez
le Chien. |
||||||||||||||||||||||||||||||||||||
|
Mise en évidence de la relation complexe entre la pa et les paramètres cardiaques Bordas p 221 doc 2 Pour voir l'allure d'un
enregistrement chez le Rat voici l'action de l'Ach (un
neuromédiateur libéré par le PS) sur la
pa chez le rat (image externe)
|
|
En attendant de trouver de vrais enregistrements - pour le tracé du PS je pense (?)
que le petit pic a été inversé avec le
grand pic, dans les périodes de repos, mais pas pour la
période de stimulation, je l'ai corrigé... * Statistiquement (in Schmidt-Nielson, p 101) la FC varie selon la loi FC = 241. M-0,25, où M est la masse de l'animal en kg (25 min-1 chez un éléphant, 600 min-1 chez la musaraigne et 1200 min-1 chez des Colibris).
**Remarque : |
|
Courbe du Bordas 2011 très
MODIFIÉE |
||||||||||||||||||||||||||||||||||
|
Analyse: |
|
Courbe du Bordas MODIFIÉE |
||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||
|
Calculer une pression moyenne n'a de sens que par rapport au débit. On considère que la la pression artérielle moyenne (pam) est égale à la pression constante qui permettrait de garder un débit constant égal au débit mesuré pour un état donné du système vasculaire. |
|
Si on assimile le réseau vasculaire à un tube de diamètre variable (de résistance à l'écoulement RT) on peut utiliser la loi de Darcy. = pam - parv = DC x RT où parv est la pression de retour
veineux (qui règne dans l'oreillette droite), DC est le
débit cardiaque et RT la résistance
périphérique totale [réseau
artériel (10% artères + 60% artérioles -
très modulable) + capillaire (15%) + veineux(15%)].
On notera aussi que dans un tel système c'est la pompe qui crée la pression: la pression s'annule lorsque le cœur s'arrête. Dans la réalité il existe d'autres pompes (voir ancien cours). |
|
On calcule la pam par un calcul mathématique complexe mais on utilise la formule approchée suivante: On notera donc que cette valeur n'est pas la moyenne arithmétique des deux valeurs extrêmes (120/80 mmd'Hg = 93 mmd'Hg et non 100 mm d'Hg) mais qu'elle tient compte de la durée de la diastole (2/3) qui est supérieure à celle de la systole (1/3). Remarques : |
||||||||||||||||||||||||||||||||||
| |
|
|
|
|
||||||||||||||||||||||||||||||||||
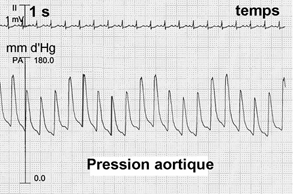
| |
La pression artérielle (définition ci-dessus) augmente fortement à l'exercice. Si une valeur de 200 mmd'Hg est courante on observe fréquemment chez des athlètes très entraînés mais non hypertendus des valeurs de 240 à 250 mm d'Hg à l'exercice. |
|
Il faut bien comprendre que les artères
n'étant pas contractiles (voir histologie ci-dessus)
mais élastiques l'augmentation ou la baisse de
la pression artérielle au niveau du point de mesure est
TOUJOURS le résultat d'un changement physiologique en
un point ÉLOIGNÉ DE CE POINT DE MESURE et qui
agit sur un autre système que le système
artériel, et qui est donc INDIRECT. L'augmentation de la Pam (voir sigles ci-dessus) est essentiellement due à celle de la Pas. L'augmentation de la Pas s'explique principalement par l'augmentation du débit cardiaque alors que le Pad change peu à l'effort. Une augmentation de plus de 15 mm d'Hg de la Pad est considérée comme pathologique et constitue un critère obligeant à stopper toute épreuve d'effort à visée diagnostique. |
|||||||||||||||||||||||||||||||||||
|
Wilmore et Costill, 2006, Physiologie du sport et de l'exercice, De Boeck Université |
|
On notera que les exercices sollicitant les membres supérieurs plutôt que les membres inférieurs font nettement plus augmenter la pression artérielle. On pense que les membres supérieurs possèdent une masse musculaire moindre et un volume circulant plus faible et donc une résistance à l'écoulement du sang plus forte que les membres inférieurs. Du fait de l'irrigation cardiaque (artères coronaires) et comme la consommation myocardique (du tissu musculaire du cœur) de dioxygène est proportionnelle au débit sanguin du myocarde (qui varie lui-même de façon directement proportionnelle à la fréquence cardiaque et à la pression artérielle systolique) tout exercice physique réalisé avec les bras augmente fortement la travail du myocarde. |
|
On peut noter une légère baisse de la pas lors d'exercices sous-maximaux lorsque l'effort est prolongé. Elle serait due à une vasodilatation des artérioles musculaires diminuant la résistance périphérique totale. Lors d'exercices de force (haltérophilie ...) on peut atteindre des pressions artérielles de 480/350 mm d'Hg. Les athlètes réalisent alors souvent la manœuvre de Valsalva qui consiste en une expiration volontaire, bouche, nez et glotte fermés. Cette manœuvre a pour effet d'augmenter la pression intrathoracique et donc la pression dans le reste du corps puisque la pression intrathoracique ainsi augmentée s'ajoute à la valeur habituelle de la pression à laquelle sont soumis les vaisseaux au niveau du cœur. |
||||||||||||||||||||||||||||||||||
 graphique
2
graphique
2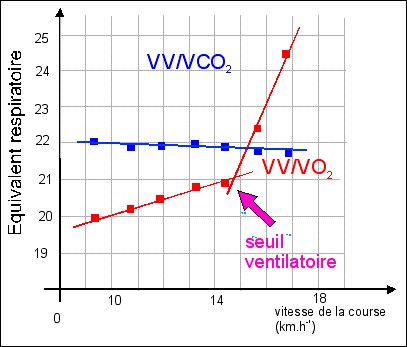


|
Dans un premier temps il m'avait semblé qu'il serait préférable, pour cette partie, de ne pas faire de cours à proprement parler car, en classe de seconde, il ne s'agit pas (et c'est le choix du programme et non pas le mien) d'apprendre des données anatomiques ou physiologiques, même regroupées de façon cohérente mais il s'agit, ce qui est bien plus difficile, de savoir utiliser des concepts biologiques pour comprendre des résultats expérimentaux. Il est parfois difficile d'éviter l'écueil de chercher à comprendre avant d'avoir appris à manipuler toutes les grandeurs utilisées dans les expériences.... Pour atteindre cet objectif, des TP ou TD (analyse de documents) seraient suffisants ; le "cours" se limitant à regrouper tous les paramètres et grandeurs vus lors des TP-TD dans un grand tableau. Après deux années d'expérience ce projet me semble irréalisable et je suis revenu à un cours plus classique. TP1 - Mesure de la ventilation pulmonaire au repos et lors d'un effort (les espaces ventilatoires) TP2 - Mesure de la consommation de dioxygène au repos et lors d'un effort TP2bis (dém) - Estimation du V°O2max TD4 - Métabolisme et bilan énergétique (métabolisme, métabolisme de base, apport énergétique d'un repas, obésité) TD5 - Fréquence cardiaque, pression artérielle et boucle réflexe TP4 - Dissection d'un cœur de mouton TP5 - Dissection d'un muscle strié squelettique (de dinde) - observation de vaisseaux sanguins sur une coupe histologique Fiche méthode - Observation microscopique Fiche méthode - Ne pas avoir peur des mathématiques en SVT |
||||||
| |
volume, concentration, fréquence, énergie, pression, débit |
|||||
|
|
|
|
||||
| |
|
|||||
|
espace contenu L.cm-3 |
|
Volume courant (0,5 L) : volume d'air échangé par les poumons à chaque cycle ventilatoire (0,5L). |
||||
| |
|
|||||
| |
Volume de dioxygène dans l'enceinte (TP2) |
|
L'air contient environ 1/5 de dioxygène pour 4/5 d'azote (N2). Il y a donc environ 18% de dioxygène dans l'air inspiré. |
|||
| |
Volume d'éjection systolique (VES) : volume de sang éjecté à chaque systole (systole = contraction cardiaque) |
|
Le VES est habituellement compris entre 60 et 150 mL, mais lors d'un effort, il peut ateindre des valeurs supérieures à 200 mL mais celles-ci, difficiles à mesurer, sont discutées par les spécialistes (PSE p 195). |
|||
| |
grandeur associée |
|
Quotient respiratoire = c'est le rapport du volume de dioxyde de carbone rejetté sur le volume de dioxygène consommé. Il permet essentiellement d'estimer la qualité des nutriments consommés (glucides/lipides/protides). |
|||
|
|
||||||
| |
|
|
|
|
|
|
|
rapport entre deux grandeurs mesurant une quantité d'un élément parmi d'autres éléments de même type %, g.L-1, mol.L-1 |
|
Concentration du dioxygène dans l'air (18%,
rapport de deux volumes) |
|
Concentration de glucose dans le sang (glycémie): rapport d'un nombre de moles ou la masse dans un litre (moyenne = 1 g.L-1 = 5,6.10-3 mol.L-1) Concentration de lipides dans le sang (lipidémie): rapport d'un nombre de moles dans un litre (trop variable pour donner un chiffre moyen). |
|
Concentration en dioxygène du sang (rapport de deux volumes: en mL par mL de sang). |
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
||
|
En travaux |
||||||
| |
|
2.2 - Comprendre l'effort |
||||||||||||||||||||||||||||||||
|
2.2.1 - Un modèle mécaniciste remis au goût du jour: la boucle réflexe |
|
|
||||||||||||||||||||||||||||||||
| |
|
2.2.1.1 - Le mécanicisme : une analogie mécanique |
|
|
||||||||||||||||||||||||||||||
|
On qualifie de mécanisme ou de mécanicisme une pensée philosophique qui explique la Nature par les lois des mouvements des corps matériels.  L'homme machine ; pour une réflexion ouverte voir Histoire de la notion de vie par André Pichot (texte personnalisé) |
|
Cette philosophie a eu un développement
très important au XVIIème
siécle mais de nos jours on peut encore dire que le
mécanicisme fait partie de nos modes de pensée
intuitifs : qui n'a pas entendu On pourrait dire que cette philosophie s'est
diluée dans le matérialisme athée. « Et
véritablement l'on peut fort bien comparer les nerfs
de la machine que je vous décrits, aux tuyaux des
machines de ces fontaines ; ses muscles et ses tendons,
aux autres divers engins & ressorts qui servent à
les mouvoir ; ses esprits animaux à l'eau qui
les remuë, dont le cœur est la source, & les
concavitez du cerveau sont les regars... » |
|
 |
||||||||||||||||||||||||||||||
| |
|
2.2.1.2 - Un réflexe est un mouvement involontaire en réponse à un stimulus |
||||||||||||||||||||||||||||||||
|
Le réflexe est un mouvement (au sens du mot grec métabolè = mouvement) qui peut être mécanique mais aussi chimique et donc métabolique : une contraction, un déplacement, une sécrétion.... |
|
Un réflexe est involontaire (il n'est pas sous commande nerveuse pour sa mise en route, même s'il passe par des centres : moelle ou bulbe) mais il peut bien sûr être contrôlé par des voies nerveuses supérieures cérébrales. |
|
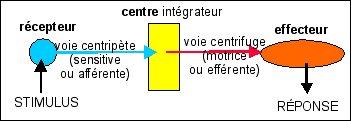 Schéma théorique d'un arc réflexe ou boucle réflexe |
||||||||||||||||||||||||||||||
|
Les expériences historiques de section-stimulation de nerfs permettant d'une façon fruste de déterminer la nature sensitive ou motrice d'un nerf dans le cadre d'un schéma réflexe simple quasi-théorique (récepteur (R) - nerf sensitif - centre - nerf moteur - effecteur (E)).
|
|
Il faut impérativement que le nerf ne comporte qu'un seul type de fibres (motrices ou sensitives) et qu'elles soient reliées au même organe (ce qui n'est pas généralement le cas, les nerfs comportant de nombreuses fibres issues d'organes différents; plus rarement des fibres sensitives avoisinant des fibres motrices). La propagation du message nerveux est à sens unique, du fait des synapses qui ne transmettent l'information que dans un seul sens (voir chapitre suivant). Le bout central (BC) d'un nerf est en liaison avec le centre (en gris) et le bout périphérique (BP) avec un récepteur (pour un nerf sensitif) ou un effecteur (pour un nerf moteur). |
|
On peut déduire deux composantes du message nerveux par ces expériences:
si l'on stimule le BC d'un nerf sensitif sectionné, comme le BP d'un nerf moteur sectionné, on doit obtenir une réponse de l'effecteur en liaison avec ce nerf. |
||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||
| |
|
2.2.1.3 - Le baroréflexe : un modèle mécanique de boucle réflexe de régulation de la pression artérielle par la fréquence cardiaque seule On a vu ci-dessus que le cœur était l'organe essentiel du soutien nutritif des muscles lors de l'effort. On a vu d'autre part que le rythme autonome du cœur (FC) était modulé par le système nerveux végétatif (S et PS). Enfin, on a vu que la fréquence cardiaque était le paramètre le plus adaptable lors de l'effort. Dans cette partie, qui cherche à comprendre les variations de la pression artérielle (pa), nous allons donc supposer que le cœur est le SEUL organe impliqué et que seule sa fréquence est modifiable par le système nerveux. C'est un modèle SIMPLISTE, voire faux, mais qui nous servira de point de départ. |
||||||||||||||||||||||||||||||||
|
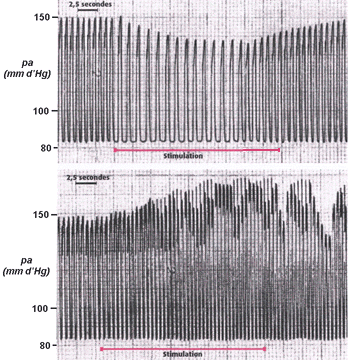
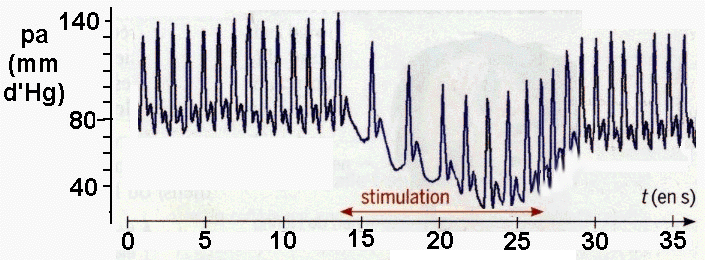
Mise en évidence du baroréflexe chez des patients attachés en position allongée à qui l'on fait effectuer un changement brusque d'inclinaison (horizontal -> vertical) - le temps a été ajouté au document et reste approximatif. Fiche
élève d'aide à l'analyse
Hypotension provisoire |
|
Le premier graphique représente un
enregistrement du BARORÉFLEXE chez un patient sain
(changement rapide de position : couché,
incliné, couché). La pression artérielle
(pa) est représentée EN HAUT, la
fréquence cardiaque (FC) en bas. Le second
enregistrement est fait chez un patient âgé
malade (atteint d'un
dysfonctionnement, par exemple au niveau des sinus - voir plus
bas). |
|
Analyse: Interprétation : |
||||||||||||||||||||||||||||||
|
Les voies sensitives du baroréflexe ont été explorées notamment par Carl Ludwig et Elias Cyon ne 1866 puis par Heinrich Ewald Hering, un vétérinaire autrichien dans les années 1920.
|
|
Le message nerveux (ou influx nerveux) est
composé de signaux "électriques"
d'intensité constante (ou PA = potentiels
d'action) mais de fréquence variable.
|
|
Analyse: |
||||||||||||||||||||||||||||||
|
|
|
Les barorécepteurs carotidiens et
sinusaux envoient aux centres bulbaires un message
sensitif codé en modulation de
fréquence : plus la pa
carotidienne ou aortique est elevée plus la
fréquence des potentiels d'actions sur les
nerfs de Hering et de Cyon est elevée. |
||||||||||||||||||||||||||||||||
|
On a donc un premier niveau de compréhension directement issu du mécanicisme. Le schéma théorique d'un réflexe peut donc aisément s'appliquer ici :
|
|
|
||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||
|
2.2.2 - Complexification du modèle mécanique en modèle cybernétique |
|
|
||||||||||||||||||||||||||||||||
| |
|
2.2.2.1 - La vie sensitive comme réponse à des stimuli |
|
|
||||||||||||||||||||||||||||||
|
Cette étape peut trouver un repère historique dans ce que certains appellent "la plus retentissante des anciennes découvertes" par laquelle François Magendie (1822) attribua aux racines dorsales des nerfs rachidiens la fonction de conduire les messages de la sensibilité et aux racines ventrales celle de conduire ceux de la motricité.  François Magendie
|
|
« Depuis longtemps, je désirais faire une
expérience dans laquelle je couperais sur un animal
les racines postérieures des nerfs qui naissent de la
moelle épinière. (…) je crus d'abord le
membre correspondant aux nerfs coupés
entièrement paralysé ; il était
insensible aux piqûres et aux pressions les plus
fortes ; il me paraissait immobile, mais (…) à
ma grande surprise, je le vis se mouvoir d'une
manière très apparente, bien que la
sensibilité y fût toujours tout à fait
éteinte. (…) Il se présentait
naturellement à l'esprit de couper les racines
antérieures en laissant intactes les
postérieures. (…) Comme dans les
expériences précédentes, je ne fis la
section que d'un seul côté, afin d'avoir un
terme de comparaison. (…) le membre était
complètement immobile et flasque tandis qu'il
conservait une sensibilité non équivoque.
» On peut trouver sur Google Books son livre et le télécharger (préférez le format ePub moins lourd (1,4 / 9,6Mo) et qui permet une recherche dans le texte alors que le pdf ne contient que des images); cependant il manque plein de pages au fichier ePub (1/2): (Leçons sur les) phénomènes physiques de la vie, Volume 2 Par F. Magendie,Societe Belge de Librairie (Bruselas) On trouve aussi sur ce site un accès à 2 publications antérieures (téléchargeable en .doc): * Magendie, F. (1822a). Expériences sur les fonctions des racines des nerfs rachidiens. Journal de Physiologie Expérimentale et Pathologique, 2, 276-279. * Magendie, F. (1822b). Expériences sur les fonctions des racines des nerfs qui naissent de la moelle épinière. Journal de Physiologie Expérimentale et Pathologique, 2, 366-371. |
|
|
||||||||||||||||||||||||||||||
|
Dans cette vision mécaniciste qui se développe au cours du XIXème siécle le fonctionnement des organes serait une réponse aux stimuli de l'environnement. |
|
La compréhension de toute activité comportementale mettant en jeu le système nerveux était une généralisation de ce qui avait été historiquement mis en évidence au sujet des réflexes médullaires (voir ci-dessous) : toute activité résultait d'un stimulus déclenchant associé à la notion de besoin physique ou intellectuel (nourriture, boisson, connaissance, sexe...). |
||||||||||||||||||||||||||||||||
| |
|
2.2.2.2 - De la vie sensitive comme programme de comportements |
||||||||||||||||||||||||||||||||
|
|
|
Le point de vue qui va ensuite se développer au XXème siècle reprend les réflexes et les intégre dans des systèmes complexes qui présentent le fonctionnement des organes comme un programme déterminé de comportements. L'activité d'un organisme résulterait d'abord d'un programme central organisateur qui définit une séquence déterminée d'étapes; lors de l'action, les informations sensitives servent à adapter, plus ou moins selon les types de mouvement, l'action à l'environnement. |
|
Cette conception est formalisée à l'aide d'outils issus de la théorie des systèmes (servomécanisme, boucles ouvertes, rétroactions, comparateurs d'erreurs...). Cette théorie implique de rechercher le lieu de la mémoire des comportements programmés. Pour certains, cette mémoire repose sur le "câblage neuronal", qui serait déterminé génétiquement. |
||||||||||||||||||||||||||||||
| |
|
On considère Norbert Wiener , auteur de Cybernetics or Control and Communication in the Animal and the Machine (1948) comme le père de la cybernétique. Dans son ouvrage, Wiener fait la synthèse de toute une série de recherches poursuivies dans le domaine des mathématiques pures (théorie de la prédiction statistique), dans le domaine de la technologie (machines à calculer, télécommunications), dans ceux de la biologie et de la psychologie, et jette les bases d'une science nouvelle, à support mathématique, destinée à couvrir tous les phénomènes qui, d'une manière ou d'une autre, mettent en jeu des mécanismes de traitement de l'information. |
|
|
||||||||||||||||||||||||||||||
| |
|
La cybernétique (du
grec "kubernêsis" = gouverner)
ou science des échanges d'information a
été, depuis le milieu jusqu'à
la fin du XXème siècle,
une clef de compréhension du fonctionnement
de l'organisme. La biocybernétique
s'est intéressée à la logique
des systèmes biologiques en se focalisant
d'une part sur la recherche d'un programme
(cet aspect a été
absorbé par la biologie moléculaire
avec sa notion de programme
génétique) et d'autre part sur
la notion d'automate abstrait (plus
ou moins intégrée au sein d'une biologie
des systèmes ou systémique). |
||||||||||||||||||||||||||||||||
| |
|
2.2.2.3 - Modèle cybernétique du baroréflexe avec le cœur comme seul effecteur |
||||||||||||||||||||||||||||||||
|
Le modèle présenté ci-contre est celui qui est présenté avec plus ou moins de bonheur dans les manuels scolaires : c'est la version cybernétique du baroréflexe étendu à l'effort. |
|
La pam est considérée comme une grandeur
à régler, c'est-à-dire
à maintenir dans des limites fixes (physiologiques). Dans cette vision mécaniciste on se contente de
dire que, lors de l'effort, le point de consigne est
modifié (voir ci-après). |
||||||||||||||||||||||||||||||||
|
2.2.3 - Le cœur n'est pas le seul effecteur : modèle cybernétique amélioré |
||||||||||||||||||||||||||||||||||
| |
|
2.2.3.1 - Intervention des autres organes que le cœur dans la pression artérielle |
||||||||||||||||||||||||||||||||
|
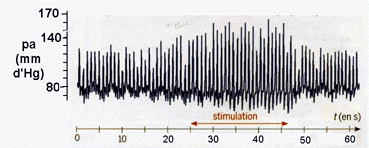
Le premier niveau de complexification consiste d'abord à tenir compte de façon plus détaillée de la résistance totale du réseau vasculaire. Modifications des divers paramètres lors d'un baroréflexe chez des patients SAINS attachés en position allongée à qui l'on fait effectuer un changement brusque d'inclinaison (horizontal -> vertical) - le temps a été ajouté au document et reste approximatif. pam = pression
artérielle moyenne,
probablement mesurée au niveau d'un membre, |
|
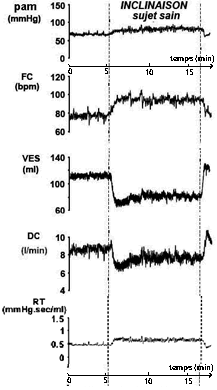 |
|
Analyse: |
||||||||||||||||||||||||||||||
| |
||||||||||||||||||||||||||||||||||
| |
|
2.2.3.2 - Modèle cybernétique du baroréflexe avec plusieurs effecteurs |
||||||||||||||||||||||||||||||||
|
Schéma cybernétique du
fonctionnement du baroréflexe (une boucle
réflexe) avec le cœur et les muscles lisses
des vaisseaux
|
|
|
||||||||||||||||||||||||||||||||
|
La boucle baroréflexe est la boucle la plus importante qui intervient lors d'un changement rapide de position, d'une hémorragie, de situations de stress... À quoi sert-elle ? |
|
En affirmant qu'elle "régule" la pa -
ce qui sous-entend que l'on considère que la pam
est une grandeur qu'il faut maintenir dans des limites
précises- on se place dans une vision
cybernétique developpée ici (on suppose un programme
de contrôle). On notera cependant que la pam
après la mise en jeu du baroréflexe est PLUS
ÉLEVÉE qu'avant la stimulation. Il est clair que l'on n'est pas là dans une démarche de compréhension de la physiologie de l'effort. |
||||||||||||||||||||||||||||||||
| |
|
2.2.3.3 - Les limites des modèles cybernétiques |
||||||||||||||||||||||||||||||||
| |
|
Une analogie très parlante de la vision cybernétique est celle de la distribution d'électricité par l'EDF qui maintient à tout instant une différence de potentiel entre le phase et le neutre de votre prise de courant de 210-200 V quel que soit le nombre d'appareils branchés sur le réseau. Dans cette vision mécaniciste, l'organisme maintient toujours une valeur fixe de la pression artérielle pour approvisionner correctement les organes en dioxygène par exemple. Il est évident qu'en cas de demande excessive, il n'est plus possible de servir tout le monde et certains consommateurs sont prioritaires... |
|
En laissant dans le flou les systèmes de contrôle (occupés par l'homme dans la réalité), cette analogie est peut-être valable pour expliquer la constance de débit encéphalique - dans la mesure où l'on a peut-être une constance du débit SANS AUCUNE ADAPTATION si ce n'est mécanique gravitaire -, mais elle échoue pour tous les autres paramètres.... |
||||||||||||||||||||||||||||||
| |
|
Pour arriver à un niveau supérieur de compréhension on est obligé de prendre en compte de nombreux autres mécanismes, notamment le système hormonal et nerveux de contrôle de la vasoconstriction et de la distribution sanguine LOCALE. Mais très rapidement le complexe devient indigeste.
|
||||||||||||||||||||||||||||||||
 Un réseau d'expression génétique au cours du développemnt... |
|
Il y a eu un très fort développement des modèles mathématiques appliqués à la biologie en ce début du XXIème siècle. Mais il me semble que les réticences sont fortes et que les partenariats sont encore le fruit d'initiatives relativement individuelles. Il faudra certainement encore du temps pour que ces modèles auxquels participent les physiologistes (parfois), des médecins (souvent), des mathématiciens (souvent) et des physiciens (rarement), soient enseignés (voir par exemple les travaux du groupe des Conférences et cours d'été Marie Curie intitulés "Modeling of human physiological systems with medical applications" (Modélisation des systèmes physiologiques humains à applications biomédicales) supportées par l'Union Européenne (2007-2010): http://www.uni-graz.at/ mc_training_schools /index.html; pour les travaux des années précédentes voir par exemple l'initiative commune de l'American Institute of Mathematics (AIM) et du Research Group on Heart-Brain Control Systems (RGHBCS) : Short-term Cardiovascular-Respiratory Control Mechanisms (Mécanismes de contrôle cardio-vasculaire à court terme) http://www.uni-graz.at/biomath/Collaboration_group/AIM/aim_index.html). |
||||||||||||||||||||||||||||||||
|
2.2.4 - Retour sur l'effort : vers des modèles continus où seules les fonctions sont stables et non pas les paramètres |
||||||||||||||||||||||||||||||||||
| |
|
2.2.4.1 Des efforts |
|
|
||||||||||||||||||||||||||||||
|
On distingue deux types d'efforts physiques (les limites n'en sont bien évidemment pas figées) qui mettent en jeu des fonctions différentes: |
|
|
|
|
|
L'échauffement, ou exercice préparatoire, en augmentant la température de fonctionnement des muscles principalement: cœur et muscles squelettiques, permet d'augmenter le V°O2 max et la contractibilité des fibres cardiaques (Hachette, n°7 p 271) |
||||||||||||||||||||||||||||
|
La pression artérielle (moyenne, diastolique et systolique) augmente à l'effort
|
|
À l'effort la résistance périphérique a tendance à diminuer avec le V°O2 alors que la pression artérielle augmente avec le V°O2
|
||||||||||||||||||||||||||||||||
|
Remarque : L'entraînement peut modifier durablement les fonctions mais aussi les structures
"Physique de la respiration chez les mammifères : efficacité ou robustesse" (http://www.larecherche.fr/ html/ LPLR/ laureats/ 2005.jsp# ministere présentation faite par Bernard Sapoval lors de la remise des prix "La Recherche" 2005: http://www.larecherche.fr /html /2008/pdf/ sapoval.pdf) |
|
Un exercice physique régulier et progressif (entraînement) est susceptible de modifier la structure des muscles cardiaques et squelettiques (Belin p 172-173). Cette modification, possible chez l'adulte bien après le développement, est qualifiée de plasticité pour les cellules et organes. Dans le cas des muscles on considère qu'il n'y a pas de renouvellement cellulaire c'est-à-dire pas de divisions. On explique donc la plasticité par le remaniement moléculaire permanent des cellules adapté à leur activité. Par exemple la cellule musculaire d'un muscle squelettique fortement sollicité voit aussi bien sa synthèse de protéines contractiles augmenter (ce qui change sa taille) que son équipement enzymatique changer. |
|
Certains paramètres, comme le V°O2 max ou la débit cardiaque présentent une nette modification à la suite de l'entraînement. Nous seulement quant à leur valeur au repos, mais aussi quant à leur variation en fonction de la puissance de l'effort. On pense que la cellule fait une utilisation différente des gènes disponibles en fonction de son activité et des contraintes de son environnement. On sait que certaines protéines contractiles musculaires sont présentes sous des formes voisines (isoformes) correspondant à des gènes voisins (isogènes). Le changement métabolique est donc interprété comme un changement de rendement de chaque gène (ce qui va dans le sens de la présentation du développement que fait R. Chandebois: voir page sur quelques éléments de sa conception). |
|
Mais l'entraînement ne peut modifier l'acquis héréditaire (au sens large) et développemental (mis en place lors du développement) * Pour l'appareil ventilatoire, il existe des contraintes
physiques (géométriques) déterminantes.
Tous les humains n'ont pas la même
géométrie pulmonaire et donc la même
efficacité (de toutes petites
différences géométriques conduisent
à de grandes différences physiologiques en terme
d'efficacité). Un
déséquilibre acquis (par
exemple du fait d'une tabagie précoce - avant 12 ans,
âge de la maturité pulmonaire - empêchant
un développement symétrique et/ou complet de
l'arborescence pulmonaire) peut avoir pour
conséquence un handicap définitif. * Pour l'appareil circulatoire, il est possible qu'il existe une plasticité plus grande que pour l'appareil ventilatoire. |
||||||||||||||||||||||||||||
|
Source: |
|
2.2.4.2 - Modèle cybernétique simplifié appliqué à l'effort (avec le cœur comme seul effecteur et le sympathique seul détaillé) |
||||||||||||||||||||||||||||||||
|
|
|
Or ce n'est pas ce qui est observé, car la pa augmente. |
||||||||||||||||||||||||||||||||
|
Explication de l'augmentation de la pa
malgré la diminution de l'activité sympathique (appréciation
semiquantitative): |
|
À l'exercice la pression
artérielle moyenne (pam) est augmentée. Pour
que le système puisse garder une efficacité,
il est indispensable, dans le modèle
cybernétique, que la courbe soit
déplacée vers des pressions plus
élevées. Ce qui nécessite donc que le
point de consigne (situé au niveau du point
d'inflexion) se déplace aussi afin de rester au
voisinage d'une valeur moyenne de la pam lors de l'effort
pour que l'organisme puisse être sensible à une
variation de cette pam. Ce formalisme est clairement déstabilisant et semble peu naturel. C'est une des critiques majeures que l'on peut adresser au système cybernétique puisqu'il revient à postuler un changement d'un élément clé du programme (la notion de programme est celle qui veut se rapprocher le plus de l'idée de fonction) alors que justement ce ne sont pas les fonctions qui changent à l'effort, mais les paramètres. |
||||||||||||||||||||||||||||||||
| |
|
2.2.4.3 - Des modèles continus |
||||||||||||||||||||||||||||||||
|
Continu versus discontinu |
|
Les deux illustrations présentées ci-contre permettent de saisir intuitivement la différence entre les modèles discontinus (à partir de réseaux) et les modèles continus (à partir de fonctions, d'attracteurs et de champs). (in D. Ingber, Int. J. Dev. Biol. 50: 255-266 (2006)) |
|
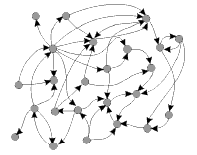 Un réseau génétique stylisé |
|
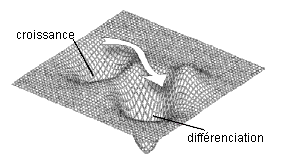 Un paysage avec 2 attracteurs |
||||||||||||||||||||||||||||
|
Waddington est avec
René Thom un des fondateurs d'une vision continue des
phénomènes du vivant (voir page
sur les modèles thomiens) Pour le vocabulaire et quelques notions sur les modèles continus voir une page sur les modèles thomiens. |
|
Quelques mots de René Thom dans Paraboles et catastrophes (1983, Flammarion, col. Champs) «… en biologie ce sont les structures mathématiques qui ordonneront les phénomènes les plus importants. Par exemple, les phénomènes de régulation exigent un grand nombre de paramètres et, par conséquent, toute description un peu affinée de ces phénomènes nécessitera l'utilisation d'espaces pluridimensionnels. croire que l'on pourra donner une théorie de la régulation tout simplement en manipulant des diagrammes cybernétiques avec des sommets et des flèches est illusoire, selon moi. L'importance de la régulation consistera toujours dans le fait qu'il s'agit d'un phénomène à caractère fondamentalement continu ; » |
|
(suite) « il faudra donc associer des figures
multidimensionnelles à ces situations. Dans ce cas la
théorie des catastrophes pourra
être très utile justement parce qu'elle vise
à interpréter des nuages de points
recueillis expérimentalement, au moyen de
constructions mathématiques qui seraient à
la fois les plus simples et les plus robustes et, ainsi,
qui satisferaient la propriété de
stabilité structurelle tout en résistant
à de petites perturbations dues au milieu ou aux
erreurs expérimentales. Toute la philosophie de
la théorie des catastrophes , son schéma
général, tient justement à ceci: il
s'agit d'une théorie herméneutique
qui s'efforce, face à n'importe quelle
donnée expérimentale, de construire l'objet
mathématique le plus simple qui puisse l'engendrer.»
(...) |
|
Pour expliciter ce que renferme cette notion
de théorie continuiste, je voudrais signaler combien il
est frappant de voir le terme d'homéostase
(du latin "homœo" ? et du grec "homolos" =
"semblable", et du latin "statio" = immobilité,
mais aussi école publique et assemblée)
employé par R. Thom notamment mais aussi par les
spécialistes des sciences cognitives, dans le sens de maintien
de la forme, aussi bien corporelle que
psychique. L'utilisation habituelle de ce terme en
physiologie étant associé à une image de
l'organisme comme système réglé dont
certains paramètres sont maintenus constants. |
||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||
|
|
|
La cybernétique est une vision discontinue du vivant. Nous nous efforcerons, dans ce qui suit, de comprendre l'effort en termes de fonctions, ce qui est bien plus facile qu'en terme de programme de commande. |
|
La tentative proposée ci-dessous, à partir notamment des travaux de René Thom, n'est en rien le reflet d'une recherche, mais une simple présentation de bon sens adaptée au niveau de l'enseignement secondaire. |
||||||||||||||||||||||||||||||
|
L'effort met en œuvre des fonctions en vue d'un travail
|
|
En physiologie, comme en physique, un effort c'est la mise en œuvre de forces en vue d'un travail.. En langage courant l'effort peut mobiliser des forces comme pour un travail intellectuel. Les forces, en biologie sont des fonctions. |
|
Géométriser les fonctions vitales c'est comprendre la vie. Même si cette compréhension repose sur des approximations inhérentes à tout modèle comme l'analyticité des fonctions considérées. C'est un bel horizon qui s'ouvre à la biologie théorique. (Pour des collègues qui liraient ces pages, je renvoie à une page en construction sur la théorie des modèles de René Thom). |
||||||||||||||||||||||||||||||
| |
Le premier principe de dynamique du vivant ou principe d'homéostasie: les fonctions du vivant sont stables (résistantes à de petites pertubations) |
|
|
|||||||||||||||||||||||||||||||
|
«un être vivant est un système homéostatique - ou en homéostase -.» ou «les fonctions du vivant sont STABLES (résistantes à de petites perturbations).» |
|
En physiologie on a l'habitude de considérer que l'organisme est en équilibre dynamique : on dit qu'il fonctionne, ou encore qu'il y a homéostase. Ce terme vient du grec homéo (ou homo = identique) et stase qui signifie position (voir cours de 1èreS). La propriété d'un système en homéostase est l'homéostasie. Un système homéostatique est un système qui est stable face à de petites perturbations. |
|
Il y a une incontestable stabilité de
l'être vivant qui maintient sa forme et se
déplace par exemple en consommant de l'énergie
qu'il doit trouver dans ses aliments. Du point de vue
physiologique, c'est-à-dire du point de vue du
fonctionnement, ce qui est stable c'est la fonction.
L'organisme maintient une stabilité de la fonction. |
|
Par analogie avec le
premier principe de la physique (formulé comme
l'équivalence chaleur-travail ou encore le principe
de conservation de l'énergie - cinétique et
potentielle), l'homéostasie est une
propriété du vivant qui peut être
formulée sous la forme d'un principe. Voir cours
de 1èreS
pour quelques données sur ce principe. |
||||||||||||||||||||||||||||
|
Comparaison des deux types de modèles |
|
Lors de l'effort,
chaque paramètre physiologique
comme la pression artérielle ou le pH
sanguin est
maintenu stable dans des
limites physiologiques grâce à un
contrôle. Lors d'un effort (dynamique), tout
comme au repos, c'est la
fonction de nutrition qui est maintenue stable. système de
contrôle externe au système
: programme génétique, programme
nerveux, programme physiologique.... La stabilité n'est pas due
à un système de contrôle, mais
c'est une
propriété du vivant.
On en reste au niveau de l'observation du
phénomène (la stabilité) et
on la modélise (par une fonction). Le niveau de chaque
paramètre dépend de très
nombreux facteurs qui sont tous sous la
dépendance du fonctionnement de nombreux
systèmes moléculaires. Chaque
molécule est elle-même sous le
contrôle de l'environnement et de sa
synthèse grâce à l'information
génétique (directement si c'est une
protéine, indirectement sinon). Plus on
approche le niveau moléculaire plus le
programme se dilue dans les
propriétés physiques de la
matière. La fonction de nutrition est
l'expression théorique et
mathématique d'une caractéristique
du vivant. Tout être vivant doit se nourrir.
La nutrition d'un organe est stable
(résistante à de petites
perturbations) parce que l'organe est vivant, il
participe donc de la fonction. Tant que l'on est à un
niveau suffisant pour que la vie s'exprime
(jusqu'au niveau cellulaire), on peut utiliser la
notion de fonction. Au niveau moléculaire, on a
que des propriétés locales et on ne
peut plus s'appuyer sur la fonction (ce qui fait
que l'on a un indéterminisme). L'analogie
reste toujours une voie de connaissance. |
||||||||||||||||||||||||||||||||
| |
|
Par exemple l'approvisionnement en dioxygène d'un muscle doit être maintenu à un niveau qui dépend de l'activité de ce muscle. Comme pour la forme dans l'espace qui est maintenue stable lors du mouvement (nous ne sommes pas des protoplastes mais notre squelette articulé et nos muscles nous permettent à la fois de changer de forme sans changer la forme globale de notre corps - on reconnaît un homme qui court comme homme aussi facilement qu'un homme allongé...), la physiologie métabolique peut être considérée comme une forme qui est maintenue stable dans un espace de grande dimension (qui comprend tous les paramètres physiologiques). On peut dire que la fonction est déployée dans l'espace des régulations. La forme de cette fonction est une géométrie qui reste stable (qui ne change pas malgré l'effort): ce que l'on qualifie d'homéostasie. |
|
Pour l'effort, l'espace de régulation comprend, outre le temps, des paramètres comme le type d'exercice, le sexe, la taille, l'âge, l'apport alimentaire, la capacité vitale pulmonaire, le volume d'éjection systolique, la fréquence cardiaque au repos, le V°O2 max et tant d'autres. Certains de ces paramètres sont quantitatifs (âge, intensité de l'exercice) mais d'autres sont qualitatifs (comme le sexe ou le type d'exercice...) et ont donc une variation discontinue. Il est clair qu'on ne sait pas actuellement représenter une telle forme. Mais dans chaque situation physiologique il serait à mon avis très profitable de commencer à utiliser les modèles mathématiques déjà disponibles (Pour modéliser une fonction que l'on suppose inaccessible dans sa complexité, il faut bien sûr faire des approximations. La plus courante, utilisée par la théorie des catastrophes, consiste à considérer toutes les fonctions comme des potentiels que la vie cherche à minimiser (ce que Thom appelle "la convention de Maxwell"). On obtient alors un modèle d'une sorte d'espace vital composé de gradients vitaux représentés par des fonctions - voir page en construction: cette convention d'approximation ne devrait pas faire peur aux physiciens. Les potentiels sont décrits mathématiquement par des conflits d'attracteurs (ce sont des objets mathématiques qui caractérisent des régimes asymptotiques, c'est-à-dire qui tendent vers une certaine stabilité). |
||||||||||||||||||||||||||||||
| |
|
Il est important de comprendre que l'organisme ne maintient pas stable des paramètres mais bien des fonctions. Ensuite cette stabilité ne se situe pas dans l'espace-temps euclidien (à 4 dimensions) mais dans un espace beaucoup plus complexe qui dépend de nombreux paramètres (et donc à n dimensions, n>4). |
||||||||||||||||||||||||||||||||
|
Dans le cadre du baroréflexe, on peut considérer qu'il intervient dans l'adaptation rapide au changement de position. L'augmentation de la pam est alors simplement la conséquence de la redistribution des forces mécaniques et gravitaires hydrauliques auxquelles l'organisme s'adapte afin de maintenir stable sa fonction de nutrition de ses organes. C'est une vision (qui est mécaniciste au sens d'Aristote mais vitaliste ou fonctionnaliste au sens moderne). |
|
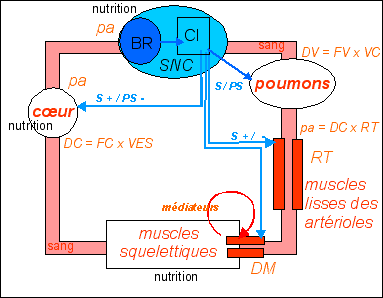 Les organes maintenant la fonction de nutrition stable lors d'un effort physique (DC = débit cardiaque, FC = fréquence cardiaque, VES = volume d'éjection systolique, pa = pression artérielle, BR = barorécepteurs, SNC = système nerveux central, CI = centres intégrateurs, DV = débit ventilatoire, FV = fréquence ventilatoire, VC = volume courant, RT = résistance totale, DM = débit (sanguin) musculaire). |
|
Lors d'un effort physique les fonctions stables sont reliées au travail de nutrition et de relation: en voici quelques-unes:
|
||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||
|
Recherche des fonctions stables au cours de l'effort |
|
Les fonctions vitales ne sont jamais constantes mais stables. La constance c'est la mort. Mais les fonctions sont maintenues stables (résistantes aux petites perturbations) dans le temps et dans un espace de contrôle de grande dimension. Si on représente un unique paramètre en fonction du temps, on peut parfois avoir une constante (le paramètre garde alors une valeur numérique fixe). exemple 1: mesure de la glycémie arrivant aux muscles (dans le sang artériel) lors de l'effort modéré On mesure ici un paramètre en fonction du temps. Celui-ci semble constant mais comme on sait bien que le muscle consomme du glucose on en déduit que les organes de stockage-destockage (le foie) compensent la consommation par une libération. Donc, la glycémie, qui n'est pas une fonction vitale mais un paramètre, est constante mais pas la fonction qui est en équilibre dynamique. Cette fonction c'est l'approvisionnement en glucose de certaines cellules. Elle est maintenue stable (non pas constante mais adaptée à la demande). exemple 2: mesure de la teneur en dioxygène du sang artériel irriguant les muscles en fonction de la puissance de l'exercice; mesure de la teneur en dioxygène du sang veineux repartant des muscles en fonction de la puissance de l'exercice; évaluation des besoins en dioxygène des muscles en fonction de la puissance de l'exercice (Belin p 136 A2, Bordas p 128 1c); la teneur en O2 du sang artériel est constante (droite horizontale); la teneur en O2 du sang veineux diminue proportionnellement à la puissance de l'exercice (droite de pente négative); la différence artério-veineuse augmente avec la puissance de l'exercice (droite de pente positive); ces trois paramètres dont le dernier n'est qu'une relation algébrique des deux premiers (Ca-Cv), ne sont pas des fonctions; la consommation en O2 n'est pas non plus une fonction vitale mais un paramètre; la fonction vitale approchée ici serait plutôt un fonction de nutrition c'est-à-dire la relation entre l'approvisionnement des cellules en énergie (et notamment en dioxygène mais aussi en nutriments) et l'énergie disponible ou en réserve (et pas uniquement la puissance déployée) ; on aurait bien une fonction qui s'adapte aux besoins, mais qui n'est pas représentable dans un espace à deux dimensions. |
|
Les fonctions sont rarement accessibles. Il est très fréquent que l'on s'intéresse à deux paramètres et que la fonction étudiée soit sous-entendue. Ce serait une grande satisfaction pour un enseignant de voir que ses élèves ont compris ce point délicat. On peut s'en rendre compte par exemple lorsque les paramètres varient à l'intérieur de certaines limites. Par exemple la fréquence cardiaque n'est pas un paramètre contrôlé et maintenu fixe (constant) pendant un effort physique. C'est un paramètre dont l'organisme joue pour réaliser une fonction comme par exemple amener davantage de sang aux muscles. Mais il est maintenu dans certaines limites (un minimum et un maximum) qui sont imposées par les structures ou les mécanismes physiologiques (rythme propre, tonus cardioaccélérateur, tonus cardiomodérateur, accélération parasympathique et modération sympathique). Exemple 3 : si l'on trace (Belin p136 n°12) la fréquence cardiaque en fonction de la consommation de dioxygène d'une jeune femme de 20 ans au cours d'un exercice avant un entraînement et après un entraînement. Le paramètre supplémentaire est ici l'entraînement (et il est qualitatif: soit la jeune femme n'a pas eu d'entraînement, soit elle a eu un entraînement de 10 semaines). La fonction étudiée est en fait de peu d'importance, ce que l'on cherche à voir c'est le rôle du paramètre entraînement sur la relation établie entre les deux premiers paramètres. Or cette relation est établie à cause d'une fonction qui est celle de l'adaptation entre ventilation et circulation, fonction fort complexe. On se contente donc de stipuler que dans l'espace considéré la fonction est régulière et représentée par la relation entre les deux premiers paramètres et l'on suppose que le fait de faire jouer un unique paramètre supplémentaire ne change pas cette relation. Ce qui est une approximation pour le moins osée mais remarquablement vérifiée. On se base sur l'homéostasie qui est une vraie propriété du vivant. (même raisonnement avec la courbe de l'exercice 3 du Bordas p 143: fréquence cardiaque et débit cardiaque en fonction de la puissance de l'exercice, avant et après entraînement). |
||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||
| |
|
|
|
|
|
|
||||||||||||||||||||||||||||
| |
|
|
|
|
|
|
||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||
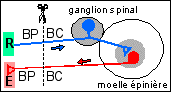
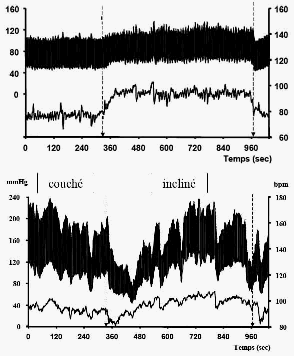

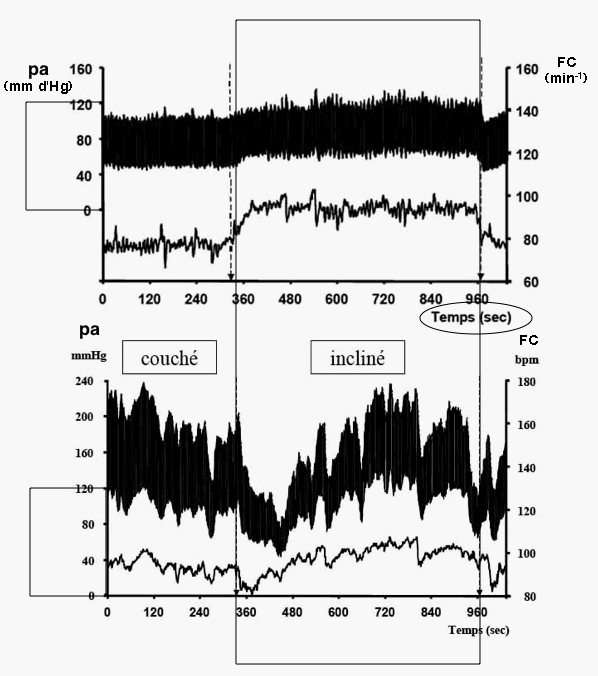
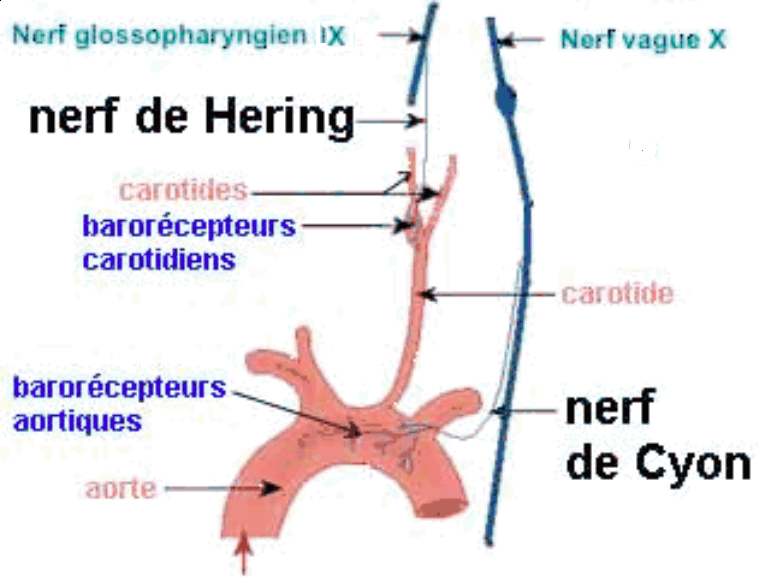
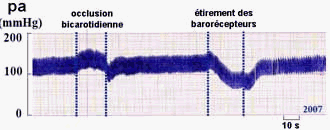
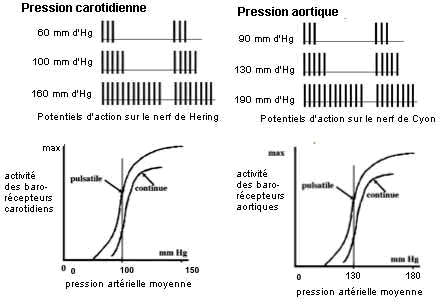
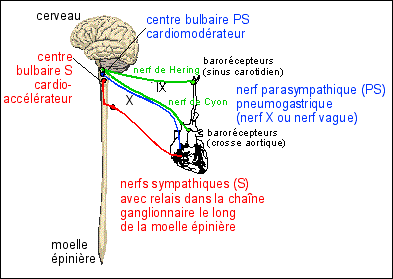

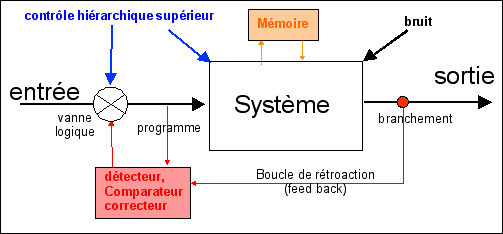

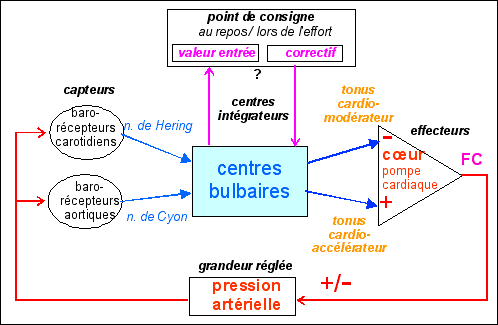
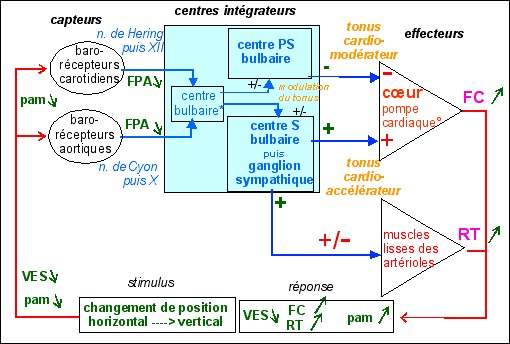

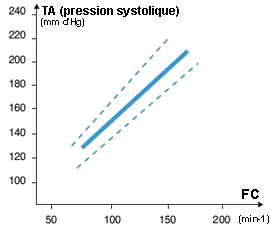
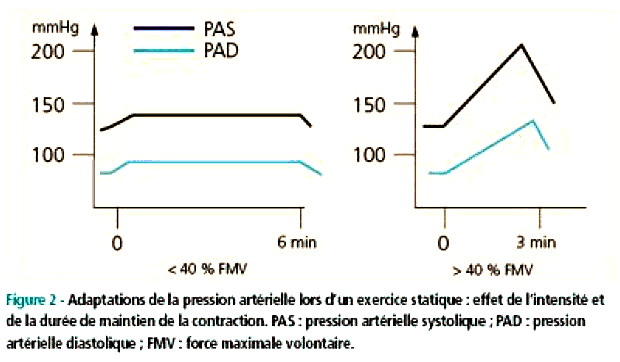
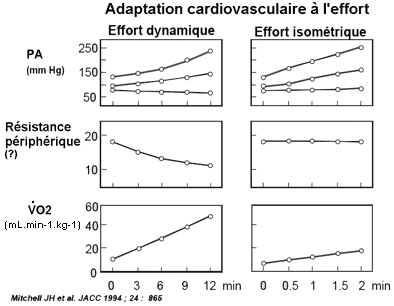
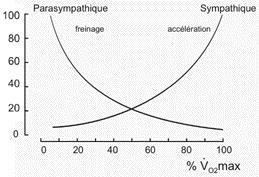
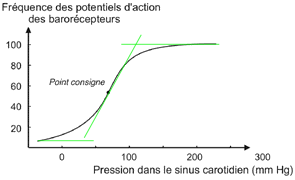
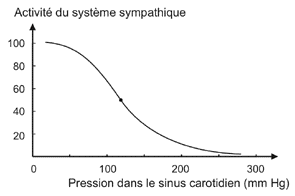
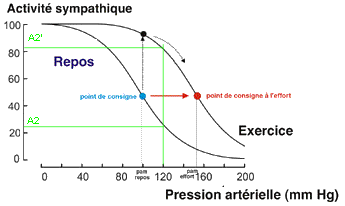

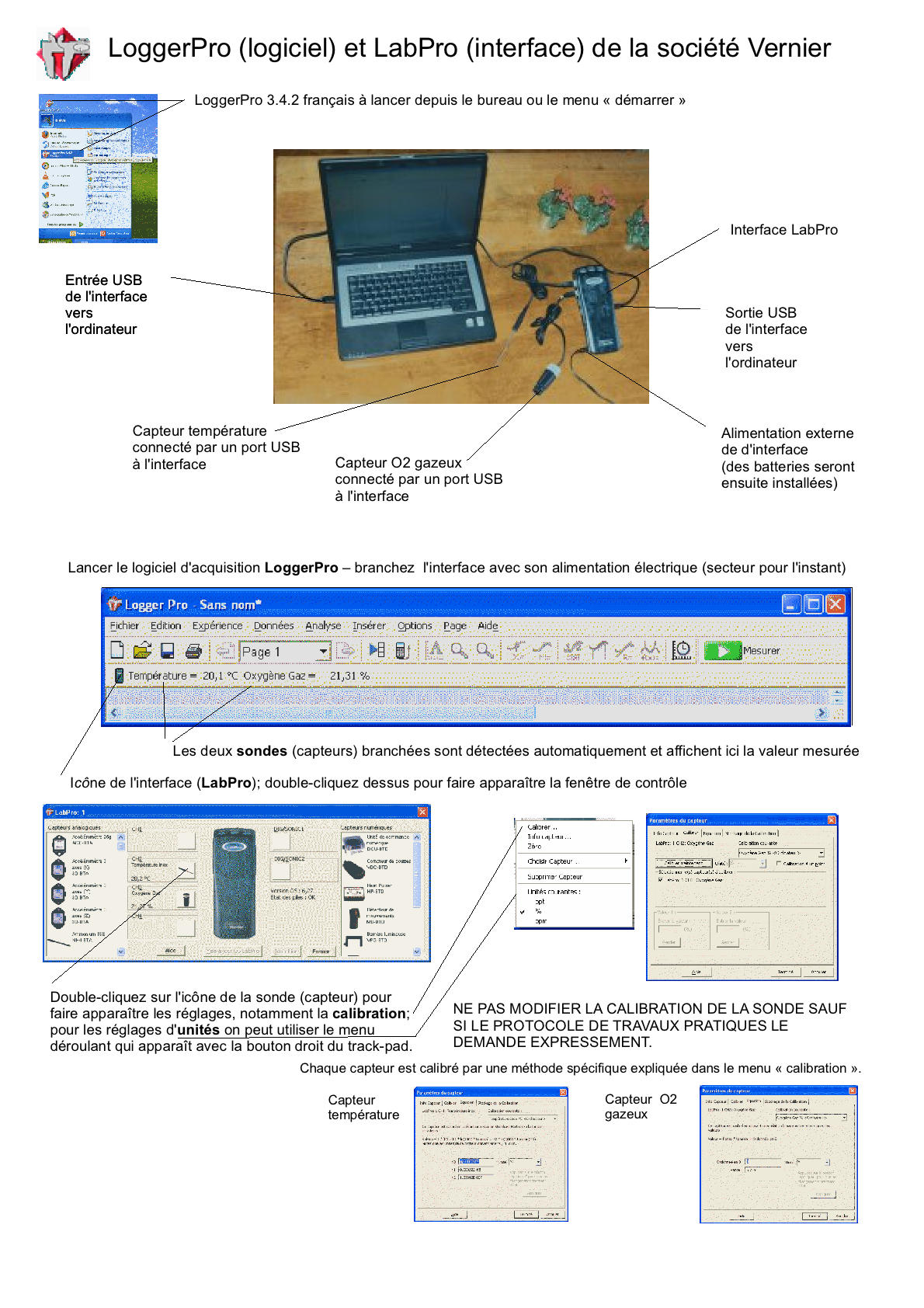{kind=link}
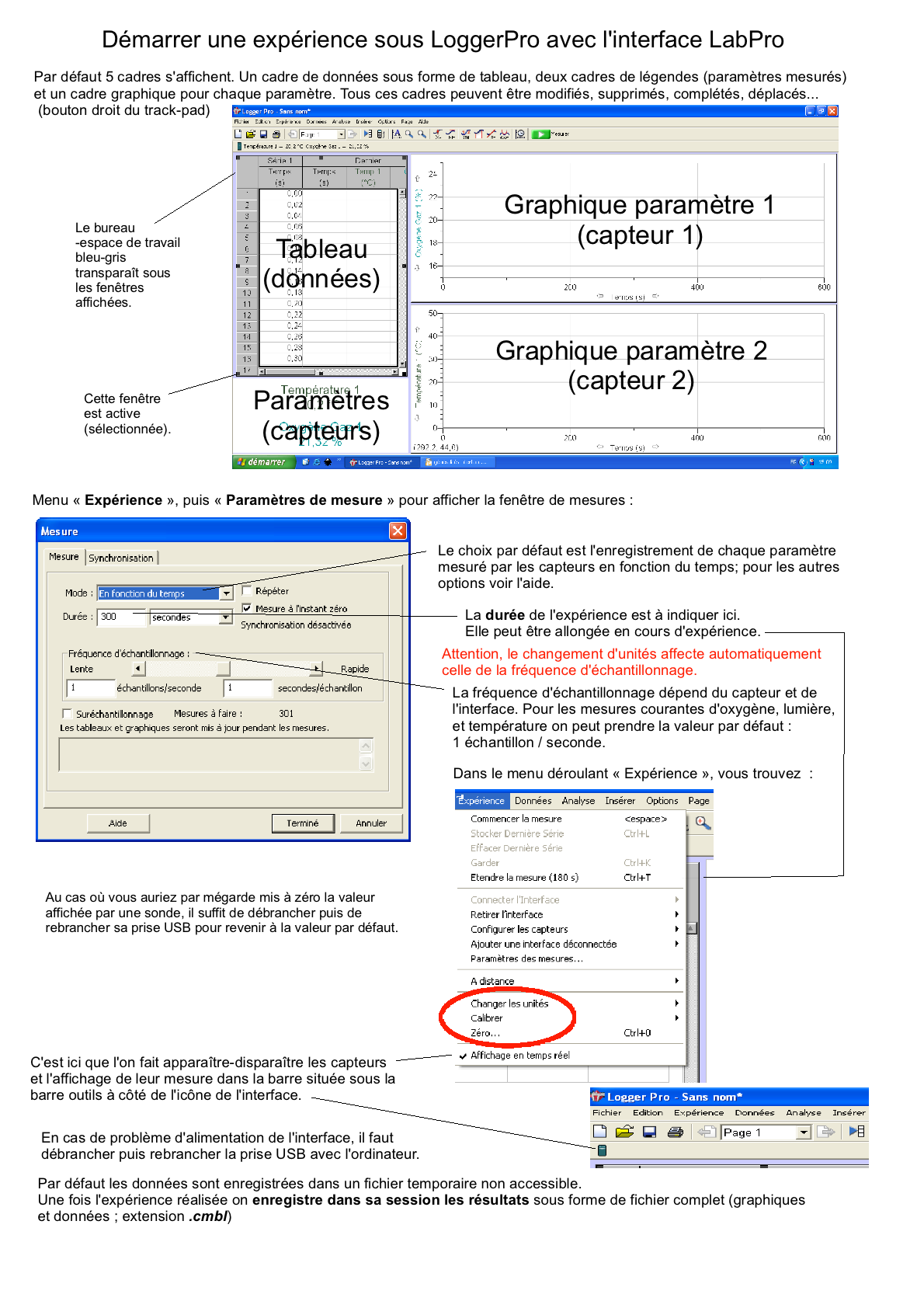{kind=link}
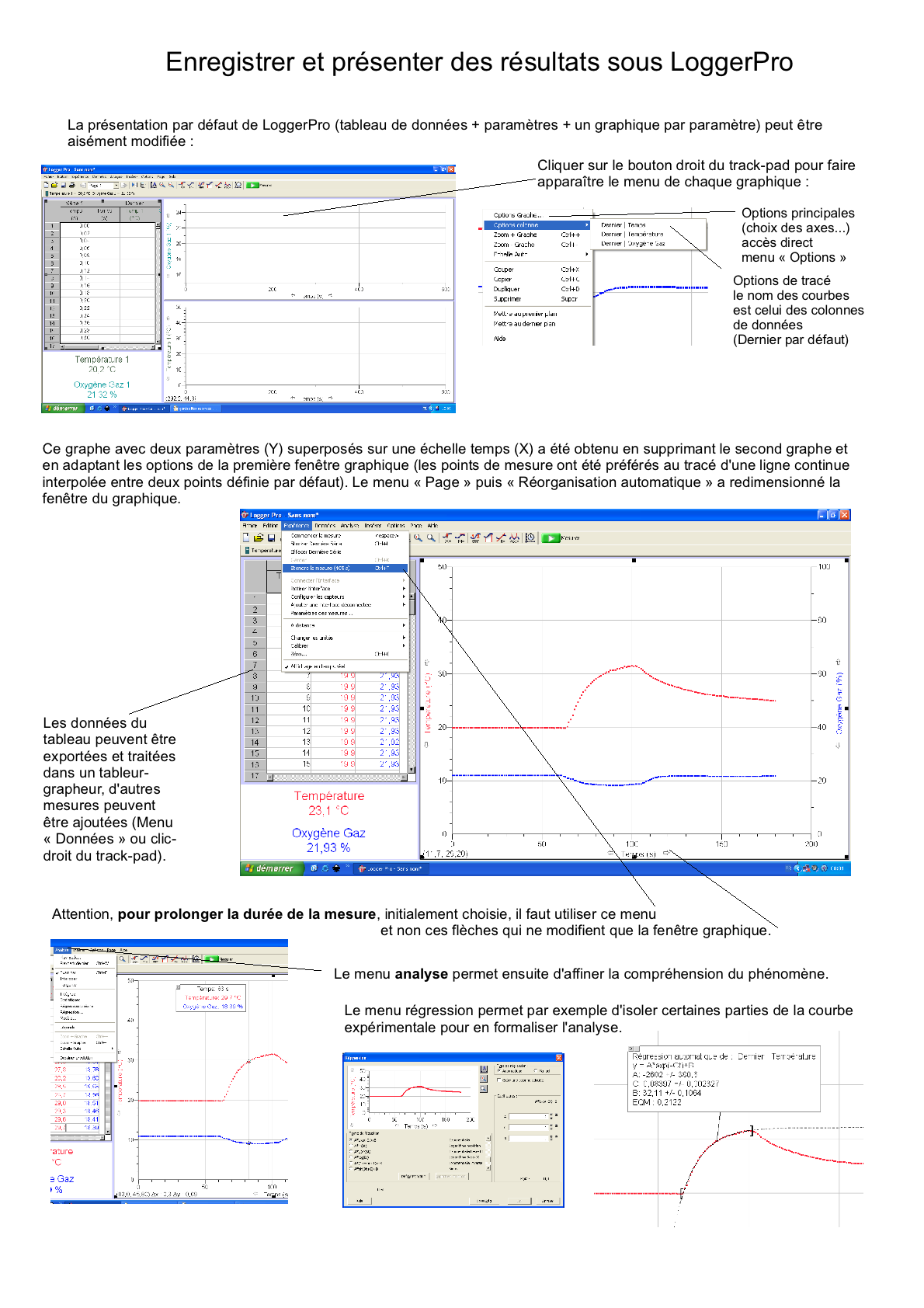{kind=link}
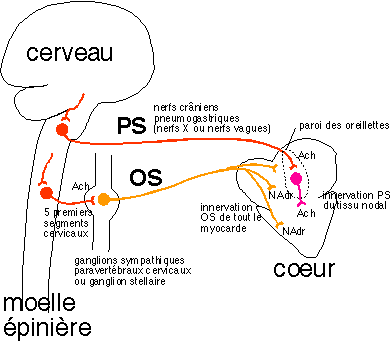{kind=link}
{kind=link}