|
retour panorama, plan du cours de 1èreS |
|
|
||||||||||||||||||||||||||||||||||||||
|
Bases
de la classe
de seconde
indispensables
: |
|
Cette page est une mise à niveau de 1èreS de l'ancienne page de seconde « Comment les cellules se multiplient-elles ?» avec des éléments des anciennes pages de 1èreS sur les divisions de croissance végétale. |
||||||||||||||||||||||||||||||||||||||
|
Plan |
|
|
||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||
|
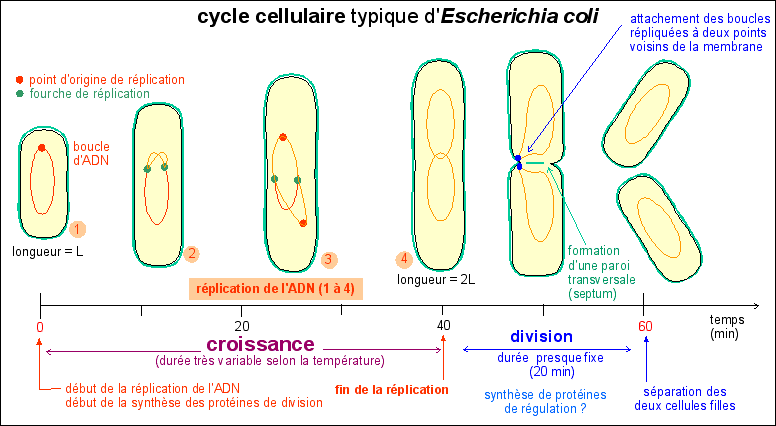
1 - Le cycle cellulaire d'un procaryote : Escherichia coli |
|
Le programme est centré sur la division des cellules eucaryotes mais un petit aperçu de la division des Prtocaryotes est plus qu'utile. |
||||||||||||||||||||||||||||||||||||||
|
Ancien
cours de
seconde: |
|
Le cycle cellulaire comprend les événements situés entre deux divisions. Il commence donc, pour une cellule, lors de sa séparation d'avec sa cellule sœur; et finit lors de sa séparation en deux cellules filles. |
|
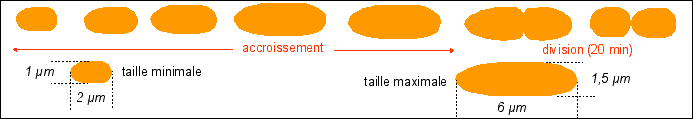
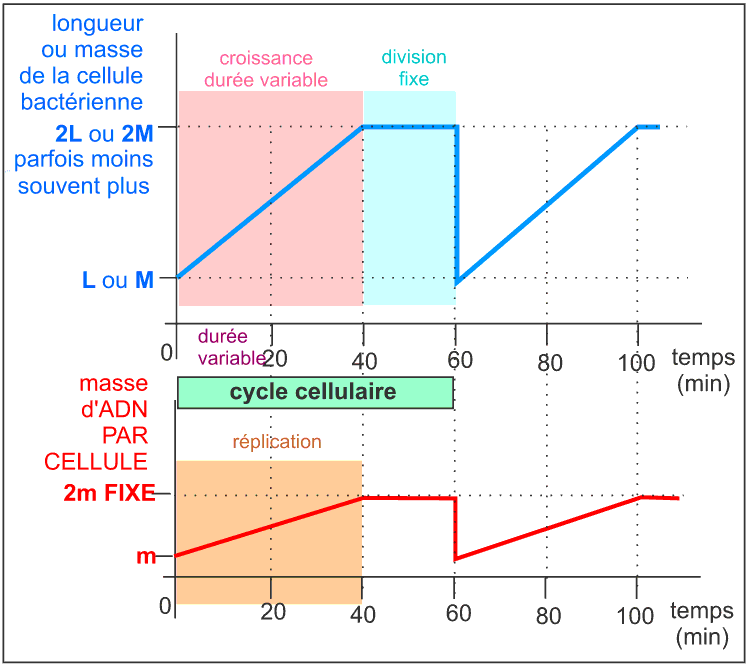
Le cycle cellulaire commence par un allongement de la cellule, sans augmentation de diamètre, qui atteint ainsi 2 fois sa longueur initiale : c'est la croissance cellulaire. Comme il n'y a pas d'augmentation de diamètre on peut penser que la masse initiale double pendant cette croissance étant donné que la bactérie peut être assimilée à un cylindre de diamètre constant. Pendant cette croissance l'ADN est dupliqué et de nombreuses protéines dites protéines de division sont synthétisées. Si l'on bloque la réplication de l'ADN on empêche la division et la cellule s'allonge démesurément en formant un long filament. La division cellulaire commence presque toujours, quelque soient les conditions de culture de la bactérie et donc quelque soit la durée du cycle cellulaire, 20 minutes avant la fin de la réplication. Mais plusieurs réplications sucessives peuvent intervenir avant la division. Ainsi E. coli pourrait contenir en moyenne environ 6 molécules d'ADN (et non pas une seule). |
|
La division a donc une durée presque fixe de 20 min alors que la phase de croissance a une durée variable selon les conditions de nutrition. La division d'E. coli se fait par scissiparité : étranglement (formation d'un septum) puis séparation (scission). Ce mode de division s'oppose au bourgeonnement (excroissance d'une sphère de petite dimension) ou aux modalités de la cytodiérèse des cellules eucaryotes qui accompagnent la mitose, qui désigne plus précisément les phénomènes nucléaires).
Le
cycle
cellulaire d'E.
coli est
donc la
succession
d'un
allongement et
d'une
séparation
d'une
structure
unique,
phénomène
dont la
durée
est sous la
dépendance
des conditions
du milieu et
de
mécanismes
moléculaires
internes comme
la
réplication
de l'ADN. |
||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||
|
Les documents cinématographiques pour les bactéries sont peu nombreux car difficiles à obtenir du fait de la petite taille des cellules (on ne peut pas prendre en film en microscopie électronique puisque l'objet observé est mort, fixé, coloré (OsO4) et observé sous vide !!!!!). Quelques petits films pris en microscopie optique avec ou sans fluorescence sont disponibles sur internet. |
|
Les films de Shapiro sont superbes : par exemple : http://shapiro.bsd.uchicago.edu/ singlecell_microcol_08_88.mov ; mais ils manquent d'explications (technique, temps séparant deux images...). |
|
Voici
un extrait
(avec
l'aimable
autorisation
de Jim
Shapiro)
présentant
la
première
division qui
dure -je crois
pouvoir lire-
1h |
|
|
||||||||||||||||||||||||||||||||||
|
|
|
Beaucoup mieux, la croissance d'une microcolonie de Bacillus subtilis (dont les certaines cellules sporulent (forment des petites spores sphériques reproductives) et d'autres meurent...) dans le dernier article de l'équipe Jan-Willem Veening, Eric J. Stewart, Thomas W. Berngruber, François Taddei, Oscar P. Kuipers et Leendert W. Hamoen (Bet-hedging and epigenetic inheritance in bacterial cell development, PNAS 18 mars 2008, vol 105, n°11, 4393-4398 : http://www.pnas.org/ content/105/ 11/4393 ). |
||||||||||||||||||||||||||||||||||||||
|
On peut aussi voir le site commercial de cells alive (http://www.cellsalive. com/ qtmovs/ ecoli_mov.htm) mais c'est à la limite de l'animation informatique. |
|
Je conseille par exemple le film n°1 qui correspond à une durée de 35h (247 images). Les 122 premières images correspondent à une image toutes les 5 minutes. Ensuite une image toutes les 12 minutes. Dans le cadre du bas les images sont prises en fluorescence (la souche modifiée par la protéine GFP est PspoIIA-gfp). |
|
|
|
film
n°1 (3,3
Mo): http://www.pnas.org/
content/suppl/
2008/03/10/
0700463105.DC1/
00463Movie1.mpg |
||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||
| |
|
Le cycle cellulaire - d'une durée de 60 min - d'une souche d'Escherichia coli à croissance lente |
|
|
||||||||||||||||||||||||||||||||||||
|
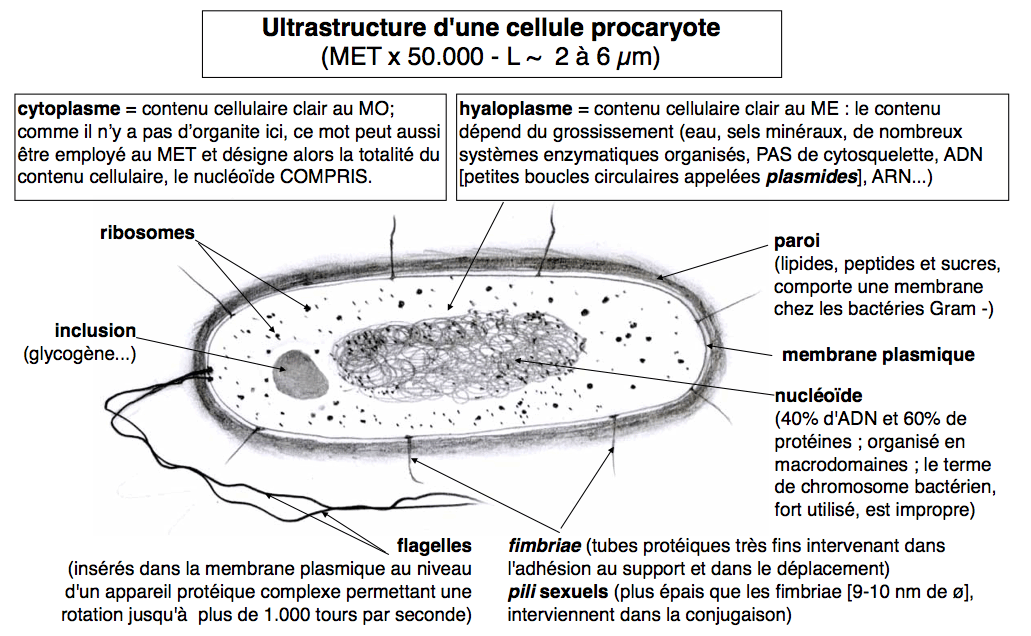
Pour une image au MET voir par exemple le site de Markus Drechsler (E. coli au MET en coloration négative à l'uranylacétate). Données moléculaires très récentes sur l'organisation du nucléoïde. |
|
La structure du nucléoïde bactérien observé en ME - sous forme d'une zone plus claire, moins opaque aux électrons et souvent attachée à la membrane cellulaire (voir cours de seconde) - reste mal connue. |
|
Il contiendrait environ 40% d'ADN, un peu d'ARN et près de 60% de protéines (mais ce n'est pas le cas de toutes les bactéries, loin de là, un grand nombre contiennent un nucléoïde d'ADN presque pur). La forme "théorique" ou plutôt le modèle, qui cadre avec les résultats expérimentaux, serait une boucle unique entortillée et empaquetée grâce à des protéines de type non-histones (différentes des protéines de la chromatine et du chromosome eucaryote) (voir chromosome). La taille de l'unique molécule d'ADN en boucle serait d'environ 1400µm chez Escherichia coli. |
|
De petits ADN circulaires ou plasmides sont le plus souvent présents dans le cytoplasme. Ils ne sont pas indispensables à la division cellulaire mais contiennent des gènes pouvant être exprimés. Ils sont répliqués de façon non synchrone avec l'ADN du nucléoïde. |
||||||||||||||||||||||||||||||||||
|
Les mécanismes moléculaires de la réplication seront traités avec la molécule d'ADN dans la partie biochimique du cours. |
|
Pour simplifier, par commodité, et comme c'est l'usage, nous représenterons ABUSIVEMENT le nucléoïde comme une unique boucle d'ADN de petite taille non enroulée. |
||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||
|
Remarque
: |
|
En effet cette description de la multiplication cellulaire bactérienne ne prend pas en compte le niveau d'organisation supérieur à la cellule (la colonie). De nombreuses bactéries gardent en commun leur revêtement de sucres (glycocalix), une paroi (streptocoques en chaînettes...). On observe une véritable différenciation entre les cellules au centre de la colonie (certaines sont mortes) et celles qui sont sur les bords... Certains chercheurs comme Shapiro n'hésitent pas à affirmer que les bactéries sont des organismes pluricellulaires. L'individu étant la colonie. Les phénomènes de multiplication des cellules sont alors comparables aux phénomènes de croissance tissulaire lors du développement d'un pluricellulaire. |
(Pour avoir une vue des travaux de James Shapiro, voir par exemple : http://shapiro.bsd. uchicago.edu/ Shapiro98AnnRevMicro.pdf ). |
|||||||||||||||||||||||||||||||||||||
|
|
|
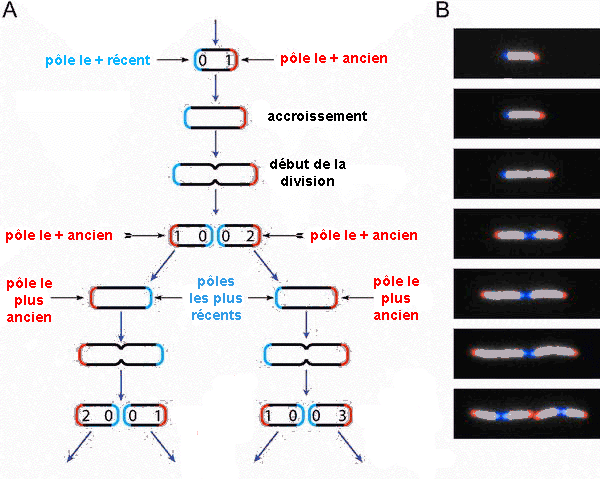
Certains résulats récents d'une équipê fraçaise viennent conforter cette vision grâce à la mise en évidence du vieillissement cellulaire des bactéries qui gardent en mémoire l'âge de leur cytioplasme. |
||||||||||||||||||||||||||||||||||||||
|
Les expériences de Eric Stewart, Richard Madden, Gregory Paul et François Taddei sont rapportées dans un article de PLOS de février 2005 (article complet en anglais disponible à l'adresse http://biology.plosjournals. org/perlserv/? request=get-toc&issn= 1545-7885&volume=3&issue=2). Je vous encourage à lire les explications en français qui sont à l'adresse http://www.forumlabo.com /2006/ actus/actus/ INSERM/0305bacterie.htm. Une des 96 vidéos (microscopie en fluorescence; 1 image toutes les 4 min de 0 à 160 min et 1 image toutes les 2 minutes ensuite) de la croissance d'une microcolonie qui atteint 505 cellules en 305 min: http://biology.plosjournals.org/ archive / 1545-7885/3/2/ supinfo/10.1371_ journal.pbio. 0030045.sv001.wmv  Une microcolonie avec reconnaissance et coloriage automatique des cellules http://biology.plosjournals. org/ perlserv/ ?request= slideshow & type=figure &doi= 10.1371 / journal.pbio. 0030058&id=21204 |
|
Ils
ont suivi au
microscope
(pendant 6h
avec 94
vidéos
(voir
ci-contre) et
sur plus de
35.000
cellules) et
analysé
automatiquement
les images
numérisées
de la
croissance
d'une culture
de la
bactérie
Escherichia
coli . Ce
suivi visuel
de la
lignée
bactérienne
a
été
rendu possible
par la
caractéristique
morphologique
de la
bactérie
qui est en
forme de
bâtonnet
et qui se
divise par
allongement,
ce qui permet
de
considérer
un pôle
ancien et un
pôle
nouveau, au
niveau du
sillon de
division (voir
l'image: http://biology.plosjournals.org/
perlserv/
?request=
slideshow&type=
figure&doi=10.1371
/ journal.
pbio.0030045&id=22668).
Les limites de
cette
méthode
sont la
formation de
plusieurs
couches de
cellules ou
bien sûr
la sortie du
champ de la
caméra.
Mais
on peut aussi
imaginer
d'autres
phénomènes
de
déplacement
de
matière
au sein de la
cellule qui
rendraient ces
observations,
purement
morphologiques
et externes
à
partir d'une
forme
figée,
fausses. |
||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
|
|
|
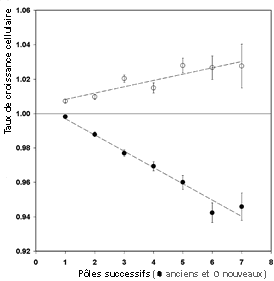
Ces résultats montrent que, pour l'espèce étudiée et dans les conditions expérimentales ci-dessus, - chaque bactérie est donc bien pour partie ancienne et pour partie nouvelle (hypothèse de continuité) - le temps de génération d'une bactérie n'est pas le bon critère pour mesurer son âge ((sa durée de vie) mais c'est le nombre moyen de divisions qui a donné naissance au cytoplasme, donc, en fait, l'âge du cytoplasme. Il s'agit donc plus d'un vieillissement du cytoplasme que de l'âge de l'individu qui pourrait être mesuré par le temps de génération. On peut donc penser que ces résultats apportent des arguments en faveur de l'hypothèse de Shapiro selon laquelle les bactéries sont des organismes pluricellulaires. L'individu le plus abouti étant la colonie. Comme dans un pluricellulaire, à un instant donné il possède des cellules anciennes et des celllules de plus en plus récentes (Pour avoir une vue des travaux de James Shapiro, voir par exemple : http://shapiro.bsd.uchicago.edu/ Shapiro98AnnRevMicro.pdf ). |
||||||||||||||||||||||||||||||||||||||
|
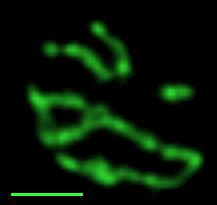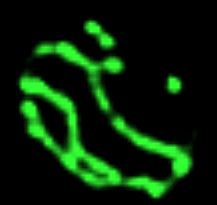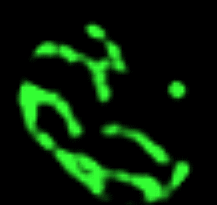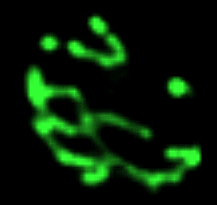
Les bactéries communiquent grâce à des nanotubes, Anne Debroise, La Recherche, 452, mai 2011, 20 Intercellular nanotubes mediate bacterial communication, Dubey and Ben-Yehuda, Cell, 144, 590, 2011 |
|
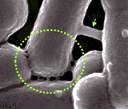 De grands (flèche verte, environ 130 nm de Ø) et petits (cercle en pointillés, environ 30 nm de Ø) nanotubes assurant une communication cytoplasmique entre des cellules de Bacillus subtilis (le diamètre du cercle est d'environ 0,8 µm) Image extraite de l'original sans autorisation mais avec une résolution très faible |
|
On
connaissait
déjà
les "pili" ou
"poils
sexuels" qui
étaient
des ponts
cytoplasmiques
établis
entre
bactéries
CONJUGUANTES.
Ces
dernières
s'échangeaient
alors du
matériel
génétique
dans le sens
donneuse ->
receveuse. Ces
nanotubes ont
été
observés
aussi bien
entre les
cellules d'une
même
espèce
bactérienne
(B.
subtilis)
qu'entre
cellules
d'espèce
différente
(B.
subtilis et
Staphylococcus
aureus). Ce mode de communication doit encore être exploré (généralité, sens du transfert, mode d'apparition des ponts, mécanisme moléculaire du transport des substances, devenir des molécules transférées...). |
||||||||||||||||||||||||||||||||||||
|
2 - Le cycle cellulaire d'une cellule eucaryote Le cycle cellulaire des eucaryotes à surtout été étudié chez la levure de bière. Les données obtenues chez les cellules en culture et les cellules embryonnaires sont les plus nombreuses pour les animaux. Selon l'organisme, l'âge de la cellule, et le type cellulaire, on a des différences fondamentales. Seuls quelques points communs ou particulièrement marquants seront présentés ici. |
|
|
||||||||||||||||||||||||||||||||||||||
| |
|
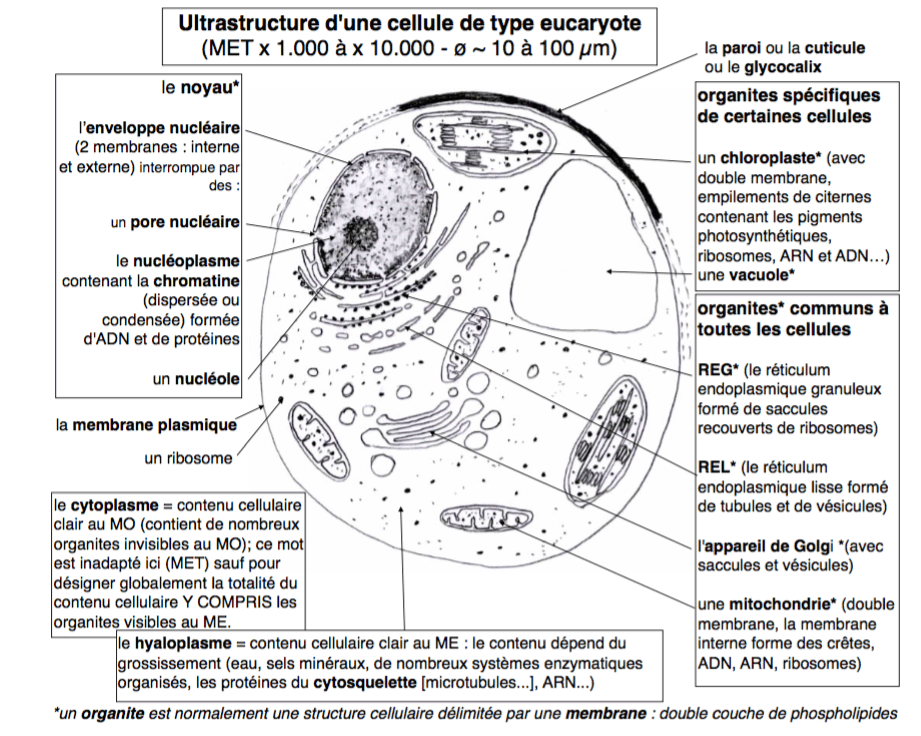
2.1 - Dans une cellule eucaryote, la plupart des structures sont en permanence l'objet de phénomènes dynamiques de fusion-séparation |
||||||||||||||||||||||||||||||||||||||
|
Les
représentations
en 3D
dynamiques
sont TOUTES
(à ma
connaissance)
très
imparfaites
étant
donné
qu'elles
supposent des
organites -
même
lorsqu'ils
sont
déformables
- comme
flottant dans
un cytoplasme
plus ou moins
liquide. Le
cytoplasme est
STRUCTURÉ
sans
pratiquement
aucune eau
libre (voir
image et page
sur l'eau dans
la cellule).
Je
préfère
encore les
dessins
statiques,
même de
piètre
qualité: |
|
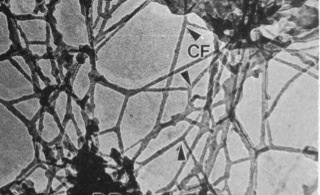
La microscopie électronique AVEC INCLUSION dans la résine, en focalisant l'attention sur les membranes, a favorisé une vision statique de la cellule basée sur une compartimentation membranaire. De plus, comme les traitements utilisés supprimaient un grand nombre de protéines cellulaires, on est passé à côté de la structure du cytoplasme (que l'on a appelé hyaloplasme, en pensant qu'il était plus ou moins vide de structure, voir Penman, 1995). Cette vision est fausse. Grâce
aux techniques
de microscopie
électronique
sans inclusion
dans la
résine
on a pu voir
la cytomatrice
dans
toute sa
richesse
(nombreux
filaments et
autres amas
denses que
l'on trouve
aussi dans le
nucléoplasme)
et surtout
visualiser les
lamina
protéiques
qui entourent
la cellule et
le noyau (face
interne) et
qui la limite
bien davantage
que les
membranes
fluides. Pour simplifier, on aurait donc une lamina externe, un cytoplasme très structuré où les organites dynamiques sont sans arrêt en train de se former et de se séparer, et une structure centrale, le noyau, au sein d'une cavité du réticulum (REG), stable, mais dont le nucléoplasme serait aussi fortement structuré et limité par une lamina nucléaire collée à l'enveloppe du réticulum qui le limite. 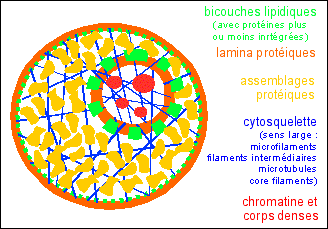 Les bicouches lipidiques des organites dynamiques (mitochondries, RE, Golgi ... ) n'ont pas été représentées. |
|
La
structuration
du cytoplasme
(matrice) me
semble
être
d'une
indiscutable
évidence.
Il est
structuré
comme un gel
qui entretient
la formation
de filaments
qui donnent la
direction des
mouvements
intracellulaires,
qui supportent
l'anisométrie
de la forme
cellulaire et
les variations
nécessaires
de
viscosité.
C'est un gel
vivant,
à la
fois dynamique
et capable de
répondre
par des
changements
structuraux
à de
nombreux
stimuli tout
en
préservant
la
capacité
à
retrouver la
forme
habituelle. |
||||||||||||||||||||||||||||||||||||
|
Quelques compléments une page avec quelques dessins simples en 3D: la cellule |
|
Le cytosquelette
a longtemps
désigné
les seuls
microtubules. Le terme cytosquelette, étant donné les progrès de la microscopie électronique, doit désormais aussi prendre en compte le réseau réticulé de protéines et autres structures observées à haute résolution dans la cytomatrice. |
|
Début 2011 a été publié dans Science un article (Berning, et al. Science 335 (6068)) décrivant les résultats obtenus à partir de la technique de la STED (Stimulated Emission Depletion) permettant l'acquisition d'une image OPTIQUE (en fluorescence - c'est ce que l'on apelle la nanoscopie avec fluorescence en champ lointain) avec une résolution pouvant atteindre 0,1 nm, soit bien moins que les 250 nm habituellement considérés comme la limite physique de l'observation d'objets à la lumière.  Les vidéos de croissance de neurites (expansions cellulaires des neurones) sont visibles sur Youtube: http://www.youtube.com/watch?v=ZOcfsf2M7jk&feature=youtu.be |
||||||||||||||||||||||||||||||||||||
|
|
|
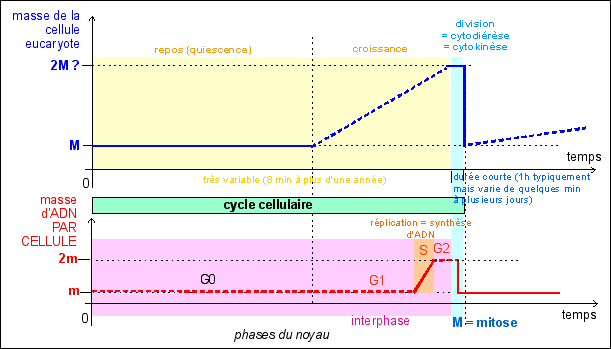
2.2 - Pendant la phase de repos ou de croissance cellulaire, le noyau est au repos en interphase, mais il réplique son ADN avant la division |
||||||||||||||||||||||||||||||||||||||
|
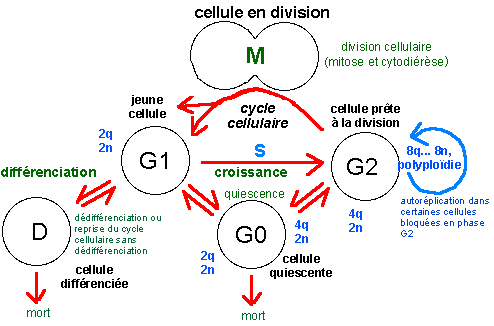
La cellule eucaryote ne doit pas obligatoirement croître ou se diviser comme la cellule procaryote, elle peut être au repos. Lors
de la phase de
repos (ou quiescence)
de la cellule,
le noyau est
aussi au
repos, mais
dans un
état
particulier (phase
G0 de
l'interphase).
Il
serait sans
doute
judicieux de
ne plus
appeler cet
état
interphase. Lorsque la cellule est en croissance, l'interphase du noyau correspond à trois phases successives: G1, S puis G2. Les
phases G (en
anglais G pour
"gap" qui
signifie
"fossé")
sont des phases
d'attente
et la phase S
la phase
de
synthèse
(S pour "synthesis"
=synthèse...
de l'ADN bien
sûr). La durée du cycle cellulaire peut être tellement allongée du fait de l'apparition d'une phase G0 que la notion même de cycle perd son sens. Chez les pluricellulaires qui renouvellent leurs tissus et organes, certains cycles cellulaires durent typiquement 24h (épithéliums, cellules immunitaires...), alors que d'autres durent toute la vie ou au moins des dizaines d'années (cellules du rein, du foie...). |
|
|
||||||||||||||||||||||||||||||||||||||
|
|
La réplication de l'ADN précède toute division (sauf dans le cas de la deuxième division de la méiose, voir cours de terminale). Nous verrons le détail des phénomènes moléculaires de la réplication de l'ADN dans le prochain chapitre. Pendant toute l'interphase, le métabolisme cellulaire est actif et de nombreuses molécules sont synthétisées. |
|
|
|||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||
|
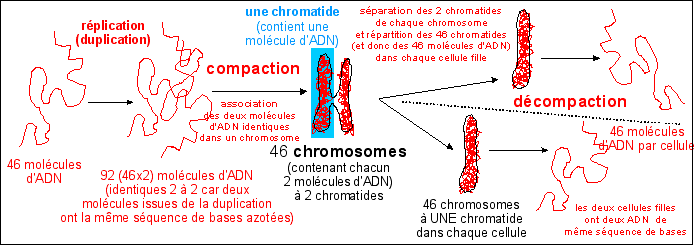
Lors de l'interphase, le noyau contient une substance qui peut être colorée très facilement par des colorants basiques : la chromatine. La chromatine contient environ 50% d'ADN et 50% de protéines. |
|
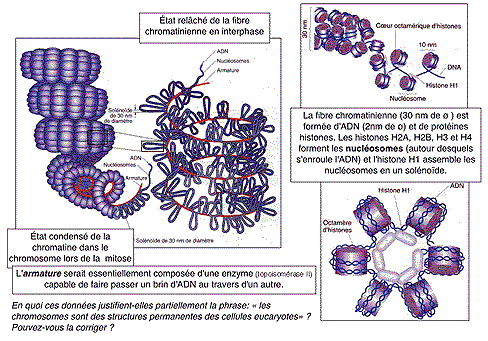
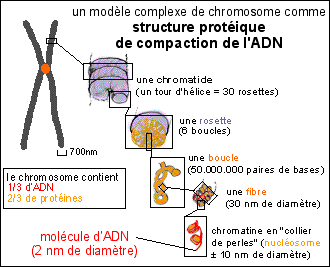
La chromatine forme habituellement un enchevêtrement de fibres (en "réseau") mais peut aussi prendre un aspect plus condensé (les zones denses sont appelées "chromocentres"). On parle de fibre chromatinienne qui aurait un diamètre d'environ 30 nm. La fibre chromatinienne est un solénoïde de 30 nm de diamètre qui comporte des protéines histones. Un octamère d'histones (composé de 8 protéines de type H2A, H2B, H3 et H4) intervenant dans la structure des nucléosomes autour duquel s'enroule l'ADN alors que les grosses molécules d'histones de type H1 structureraient un assemblage de 6 nucléosomes en un solénoïde pour former la fibre. (voir document TP)
|
|
On peut la mettre en évidence par coloration de Feulgen ou par le test de Brachet Dans
les coupes
ultrafines
observées
au microscope
électronique
(MET) la
technique est
bien
différente
de la
coloration
réalisée
pour le MO.
Mais on garde
le terme de
chromatine
pour
désigner
la substance osmiophile
(qui fixe le
tétroxyde
d'Osmium, le
colorant
opaque aux
électrons
utilisé)
MAIS ce n'est
pas
forcément
la même
structure que
la chromatine
vue au MO. |
||||||||||||||||||||||||||||||||||||
|
Remarque: |
|
Les
protéines
de la
chromatine
sont de 2
types : L'association protéines-ADN est dynamique avec une durée d'association de l'ordre de quelques minutes (l'association se forme et se défait sans cesse). (HCC, p 42) Le ou les nucléoles sont des inclusions très facilement colorables par les colorants basiques, la plupart du temps sphériques et d'aspect homogène. Ils sont constitués par des zones particulières (organisateur nucléolaire) de certains chromosomes. Ces nucléoles, en nombre défini pour une espèce, contiendraient de multiples exemplaires des gènes codant pour des ARN ribosomiaux. Il existe aussi une certaine quantité d'ADN extrachromosomique chez certaines espèces. |
|
|
||||||||||||||||||||||||||||||||||||
|
TP9 - Mitose et chromosomes |
|
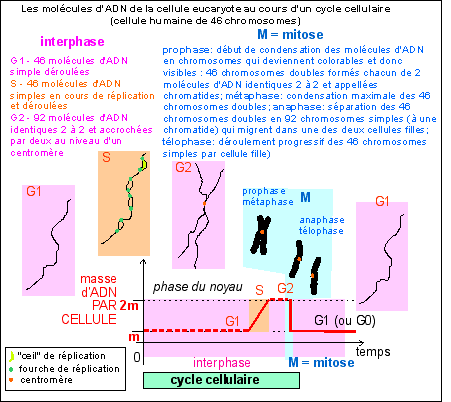
2.3 - Pendant la phase de division cellulaire (cytodiérèse), le noyau subit une mitose |
||||||||||||||||||||||||||||||||||||||
|
|
La
division
cellulaire
comprend : |
|
Les organites sacculaires (en forme de sacs) et/ou tubulaires (en forme de tubes) comme les mitochondries (et les chloroplastes dans les cellules qui en possèdent), l'appareil de Golgi et le réticulum endoplasmique sont DYNAMIQUES et donc, lors de la cytodiérèse, ils n'ont aucune difficulté à se séparer pour se répartir dans les deux cellules filles. La cytodiérèse est habituellement commandée par un anneau de filaments d'actine (une protéine contractile) qui se permet m'étranglement puis sa séparation du cytoplasme en deux cellules filles. |
|
|
||||||||||||||||||||||||||||||||||
|
2.3.1 - La mitose commence par la condensation de la fibre chromatinienne en chromosomes |
|
|
||||||||||||||||||||||||||||||||||||||
|
L'étymologie
du mot chromosome
est
suffisamment
précise
pour que l'on
réserve
le terme aux bâtonnets
colorables
apparaissant lors
de la mitose chez
les eucaryotes.
Seulement, il
existe de
nombreuses
personnes qui
persistent
à
employer le
mot chromosome
pour
désigner
toute
molécule
d'ADN d'une
cellule, tant
procaryote
qu'eucaryote.
Il faut
éviter
cette
confusion. Les
membres du GTD
qui ont
énoncé
le programme
de
1èreS
n'ont pas fait
mieux en
affirmant:
«Les
chromosomes
sont des
structures
constantes des
cellules
eucaryotes qui
sont dans des
états
de
condensation
variables au
cours du cycle
cellulaire.»
Certes le mot
n'est pas
élargi
aux
procaryotes...
explicitement,
mais il est
évident
que le mauvais
usage ne peut
que se
répandre. Du
point de vue
chimique,
c'est la chromatine
(ADN +
protéines)
extraite du
noyau sous
forme d'une
fibre plus en
moins
enroulée
qui est
permanente. On
pourrait donc
dire que «
la fibre
chromatinienne
est
une structure
permanente du
noyau des
cellules
eucaryotes».
En
tout cas il
est hors de
question
d'appeler
"chromosome"
la
molécule
d'ADN des
procaryotes,
qui n'est que
parfois
associé
à des
protéines,
qui d'ailleurs
ne sont pas
des histones. |
|
La mitose commence avec l'apparition des chromosomes (du grec chromo = couleur et soma = corps) qui sont des bâtonnets plus ou moins allongés, facilement colorables et qui sont déplacés grâce à un réseau assez complexe de microtubules qui forment le fuseau mitotique. Les chromosomes sont des organites (au sens large, car ils n'ont pas de membrane limitante) de quelques µm de long pour 0,7 µm de diamètre. 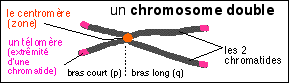 |
||||||||||||||||||||||||||||||||||||||
|
Lors de la mitose (division du noyau, voir ci-dessous) toutes les observations portent à croire que la chromatine se condense en chromosomes.
|
|
Les chromosomes sont aussi facilement colorables que la chromatine avec des colorants basiques. Les chromosomes contiennent plus de protéines que d'ADN (1/3 d'ADN et 2/3 de protéines). Les protéines se répartiraient pour moitié en protéines histones et pour moitié en non-histones. |
|
Image
provisoire
venant du TP
9 - Mitose et
chromosomes
(extrait de
Analyse
génétique
moderne, fig
2-22) |
||||||||||||||||||||||||||||||||||||
|
Dans
un film
du CERIMES (http://www.cerimes.fr/la-banque-dimages/
mitose-debut-de-prophase
-a-telophase.html),
il est
particulièrement
clair que ce
sont des
territoires
nucléaires
qui
donnent
naissance aux
chromosomes. Références
de l'article:
Three-Dimensional
Maps of All
Chromosomes in
Human Male
Fibroblast
Nuclei and
Prometaphase
Rosettes |
|
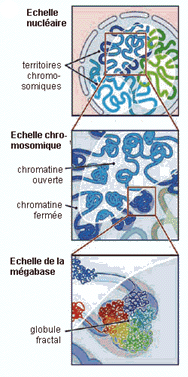
Pour des données plus récentes, on peut voir par exemple un article en anglais accessible sur le net librement (licence Creative Commons Attribution) est à l'adresse http:// biology.plosjournals. org/archive/ 1545-7885/3/5/pdf /10.1371_ journal.pbio. 0030157-S.pdf dont voici une figure (Fig. 1-D - réf: 10.1371/journal.pbio.0030157.g001) particulièrement accrocheuse: 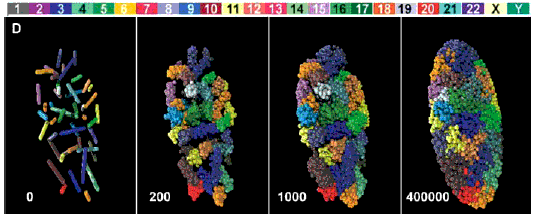 Le positionnement des 46 chromosomes (dans un état de décondensation croissant de la gauche vers la droite) d'un fibroblaste humain en reconstitution 3D simulant les résultats obtenus par microscopie (voir l'article pour des précisions) J'aime beaucoup la légende ajoutée par La Recherche (brève du n°388, juillet-août 2005, 15): Paysage nucléaire, qui rappelle le paysage épigénétique de Waddington (voir page annexe). |
||||||||||||||||||||||||||||||||||||||
|
Remarque: |
|
Remarque
; Les boucles rattachées à une armature seraient dans un état plus ou moins condensé selon l'état de la chromatine. La question de savoir si cette fibre correspond à une unique et gigantesque molécule d'ADN - comme cela est le plus souvent affirmé - ou bien si l'ADN se trouve sous forme de petites boucles enchâssées au niveau de l'armature de la chromatide, est finalement plus une question de dynamique que de structure, étant donné que l'on pense que l'armature est justement principalement composée du système enzymatique (topoisomérase II) capable de couper et recoller extrêmement rapidement la molécule d'ADN. Des électronographies de structures de chromosomes "éclatées" sont données pour "preuve" de la présence d'une unique molécule. |
|
|
||||||||||||||||||||||||||||||||||||
|
|
|
 coloration bleu de méthylène, cellules de racine d'oignon http://www.microscopy -uk. org.uk/ mag/artnov 04macro/ jronionroot.html |
||||||||||||||||||||||||||||||||||||||
|
Caryotype
|
|
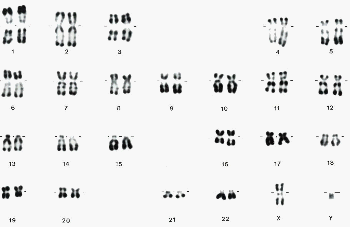
La technique qui consiste à colorer les chromosomes puis à prendre un photo lorsqu'ils sont étalés (en vue polaire en métaphase de mitose, la cellule étant éclatée) pour les dénombrer est appelée technique de caryotypage. On dit que l'on réalise un caryotype. (image ci-dessous)La coloration la plus ancienne pour les chromosomes , réalisée depuis les années 1970-1972, est celle qui permet de mettre en évidence des bandes (méthode du banding en anglais). |
|
Des bandes claires (non colorées) alternent avec des bandes sombres (colorées). On obtient jusqu'à 800 bandes pour l'ensemble du caryotype haploïde (n chromosomes). |
|
Toutes ces colorations n'ont aucune explication satisfaisante: leur origine est UN MYSTÈRE... en absence de traitement, la coloration est pratiquement uniforme, mais étant donné que les traitements sont très variés, on ne connaît aucune cause commune certaine. La coloration est reproductible et peut permettre de repérer des translocations (déplacement de fragments d'un chromosome à un autre). Elle est très homogène au sein d'une espèce : il y a peu de variations individuelles. |
||||||||||||||||||||||||||||||||||
|
(http://www.ac-reims.fr/ datice/svt/d ocsacad/ caryotype/caryo2/ caryonormal.htm) |
|
22 paires d'autosomes (chromosomes qui ont deux paires ayant LES MÊMES BANDES): A (paires 1, 2 et 3), chaque chromosome pouvant être reconnu par les bandes colorées 1 paire de gonosomes (paire 23): ou chromosomes sexuels ; ils ont les mêmes bandes chez la femme (XX) mais ni la même taille ni les mêmes bandes chez l'homme (XY) |
||||||||||||||||||||||||||||||||||||||
|
2.3.2 - La mitose comporte 4 phases basées sur l'aspect et les mouvements des chromosomes |
|
|
||||||||||||||||||||||||||||||||||||||
|
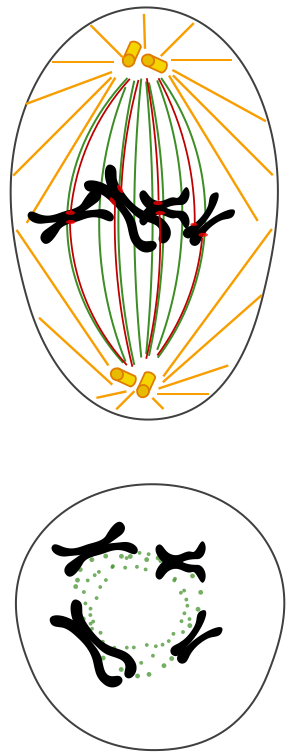
Les 4 phases de la mitose correspondent aux différents états des chromosomes, à leur condensation, leur répartition dans la cellule, et à leurs mouvements. Ces phases se succèdent de façon CONTINUE et il est bien difficile de délimiter chacune précisément. prophase : apparition des chromosomes (doubles, à 2 chromatides identiques) par condensation de la chromatine, disparition de l'enveloppe nucléaire (non visible au MO) qui se désagrège par fragmentation des citernes du réticulum qui la constituent. Formation du fuseau de division (occupant le centre de la cellule) constitué de microtubules (dits "fusoriaux) synthétisés à partir du centre organisateur à chaque pôle de la cellule (qui contient deux centrioles - dédoublés juste avant la mitose - sauf chez les plantes, mais aussi certains autres organismes, notamment unicellulaires). métaphase : disposition des chromosomes dans un tore (anneau creux) situé sur le pourtour du fuseau de division et dans le plan équatorial ; on parle de migration des chromosomes, car ils sont TIRÉS et POUSSÉS par les microtubules kinétochoriens qui, en croissant à partir des pôles, viennent se fixer aux kinétochores; le plan équatorial décrit de façon précise l'orientation des deux cellules filles (il est matérialisé par un anneau ou un disque de filaments d'actine -formant la plaque équatoriale-, voir plus bas); si la cellule est étirée, ce plan peut diviser la cellule longitudinalement ou transversalement ce qui donnera une division en épaisseur ou en longueur, déterminante pour la croissance des cellules végétales à paroi par exemple. anaphase : séparation (ou clivage) des deux chromatides de chaque chromosome et migration de chaque chromosome fils (ou chromatide sœur) vers chaque pôle de la cellule.Chaque chromosome fils est TIRÉ par ses microtubules kinétochoriens qui glissent le long des microtubules du fuseau de division; télophase : regroupement des chromosomes fils, puis disparition des chromosomes (par décondensation de la chromatine), du fuseau de division, et reformation de l'enveloppe nucléaire (non visible au MO).
|
|
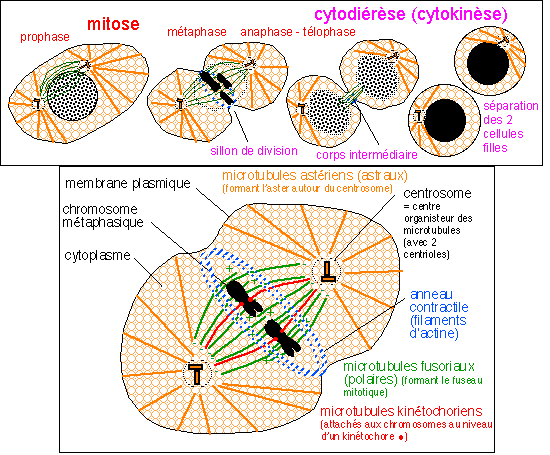 Quelques étapes de la division cellulaire d'une cellule "animale" mettant en évidence le rôle des microtubules et de l'anneau contractile de filaments d'actine intervenant dans la cytodiérèse. Comme organites, seuls les chromosomes sont représentés. |
||||||||||||||||||||||||||||||||||||||
Division
cellulaire
d'une cellule
de type animal
(tous les
organites,
excepté
le noyau - et
les
chromosomes
lorsque le
noyau
disparaît,
ont
été
volontairement
omis) En
prophase
les
chromosomes
apparaissent
par
condensation
de la
chromatine.
Les centrioles
se dupliquent
et migrent aux
deux
pôles de
la cellule.
Les microtubules
astériens
donnent la
forme à
la cellule et
les microtubules
fusoriaux apparaissent
et commencent
à
former le
fuseau.
L'enveloppe
nucléaire
se fragmente.
Dans ce
schéma
les
chrosomomes
sont bien
condensés
(on a
représenté
2n=4) et sont
doubles car
ils
comprennent
deux
chromatides
identiques.
C'est une
prophase
dès que
la chromatine
se condense et
tant que les
chromosomes de
se
déplacent
pas.
|
 La
métaphase
débute
lorsque les
chromosomes
(doubles)
commencent
à se
déplacer.
Plus
exactement ils
sont
tirés
et
poussés
par les microtubules
kinétochoriens
(accrochés
aux
kinétochores)
qui glissent
le long des microtubules
fusoriaux.
Les
chromosomes se
placent ainsi
dans un tore
(anneau creux)
bien visible
sur une vue
polaire
(en bas). La cytodiérèse
n'est pas
encore
commencée
malgré
un allongement
de la cellule
grâce
aux microtubules
astériens
qui croissent.
|
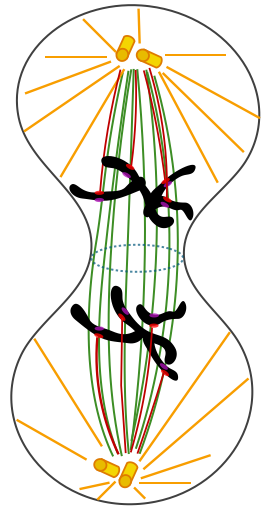 L'anaphase
débute
très
précisément
à la
séparation
des
chromatides de
chaque
chromosome (on
parle de clivage
des
chromosomes
doubles). Les
chromosomes-fils
"migrent" aux
pôles :
en fait ils
sont
tirés
et
poussés
par les microtubules
kinétochoriens
qui glissent
toujours le
long des
microtubules
fusoriaux. La
cytodiérèse
commence
grâce
à la
contraction de
l'anneau
d'actine
présent
dans le plan
équatorial
de la cellule.
|
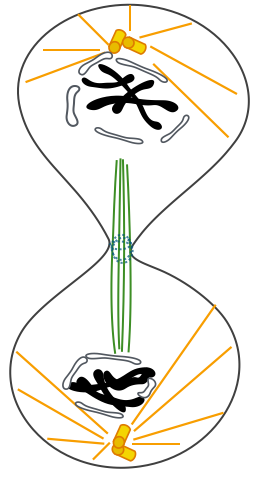 On
parle de télophase
lorsque les
chromosomes
fils se
regroupent aux
deux
pôles.
La fin de la
télophase
est
marquée
par la
disparition
des
chromosomes.
Ici la
télophase
n'est pas
encore
terminée
puisque les
chromosomes ne
sont pas
décondensés.
L'enveloppe
nucléaire
se reforme. La
cytodiérèse
n'est pas
encore
terminée
: les deux
cellules
filles sont
encore
réunies.
La
cytodièrèse
peut
intervenir
lorsque les
noyaux des
cellules
filles sont
déjà
en interphase.
|
|||||||||||||||||||||||||||||||||||||
| |
|
Les phénomènes présentés concernent toujours des divisions normales mais on connaît depuis que l'on a observé des divisions in vivo des anomalies dans la séparation ou dans la répartition des chromatides à l'anaphase, dans la formation des asters (unique non dupliqué ou au contraire au nombre de 3...)... Chez de nombreux Protistes (unicellulaires) notamment, il existe aussi des mitoses atypiques sans fuseau de division, sans disparition de l'enveloppe nucléaire... |
||||||||||||||||||||||||||||||||||||||
|
On ne cesse d'améliorer les techniques de visualisation des cellules en mitose. Un article en libre accès (mais en anglais) d'Inoué et Oldenbourg, 1998 montre quelques avancées en lumière polarisée : une microscopie optique couplée à l'analyse d'image en temps réel à l'aide de ce qu'on appelle un Pol-scope.
|
|
Ce n'est qu'en 2011qu'une équipe américaine (Magidson et col.) a pu suivre le déplacement des chromosomes pendant toute la durée d'une mitose - voir références dans l'article Science in School n°25). |
|
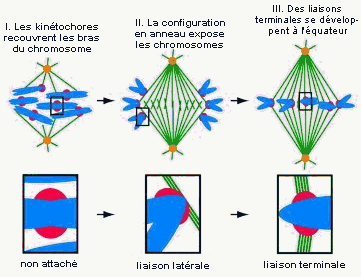
« ... nos résultats prouvent l'instabilité des interactions latérales entre les kinétochores et microtubules lors des débuts de la prométaphase. Ces interactions transitoires entraînent la disposition des chromosomes dans un cercle équatorial sur le pourtour du fuseau. Un modèle informatique a montré que cette répartition toroïdale [selon un tore] des chromosomes expose les kinétochores à une forte densité de microtubules ce qui facilite la formation ultérieure des liaisons amphiteliques [chaque chromatide de chaque chromosome est liée à des microtubules d'un seul pôle]. Ainsi, la formation du fuseau nécessite une étape préalable mal connue de prépositionnement du chromosome qui favorise la formation des liaisons amphiteliques.»
|
|
Quelques
vues d'artiste
assez
fidèles
aux
modèles
moléculaires
actuels sur
WEHI.TV en
anglais (accès
direct
à la
vidéo
en
français)
À
voir sur
Youtube |
|
|||||||||||||||||||||||||||||||||
|
|
Pour
la méiose
c'est au
sein du
laboratoire de
Jan Ellenberg
(EMBL) que
Tomoya (Tomo)
Kitajima avait
suivi en
premier les 10
heures de la
double
division
sexuelle. |
|
-
l'ovocyte est
une cellule
gigantesque et
ce fût
une prouesse
technologique
que d'arriver
à y
trouver et
à y
suivre les
chromosomes
lors de la
méiose; |
|||||||||||||||||||||||||||||||||||||
| |
|
|
||||||||||||||||||||||||||||||||||||||
|
Remarque
: |
|
Aucune voie n'est complètement fermée tant que la cellule est vivante. Il n'en reste pas moins que certaines voies sont nettement plus empruntées que d'autres. Dans la phase de développement embryonnaire la plupart des cellules nouvellement formées se divisent à nouveau, même si parfois la division est inégale dans le sens où seule l'une des deux cellules filles garde son pouvoir mitogène, l'autre se différenciant (on parle de division formative par opposition à une division proliférative, où les deux cellules filles gardent leur pouvoir mitogène). Les divisions chez un organisme adulte sont nettement plus localisées (c'est le renouvellement cellulaire) et souvent dédiées à des cellules spécialisées (ou cellules souches) qui présentent la plupart du temps des divisions formatives. En bleu, les quantités d'ADN par cellule (2q étant la quantité unité dans une cellule diploïde) et nombre de chromosomes (2n étant le nombre de chromosomes dans une cellule diploïde); la phase S (synthèse) du cycle cellulaire est la phase de réplication ou duplication de l'ADN. La lettre G fait référence au mot anglais "gap" désignant un fossé ou état stable de la cellule (d'après la source citée, fig 1, p 498). Au cours du cycle cellulaire, on différencie l'interphase (qui comprend les phases G1, S et G2) et la phase de division cellulaire (M, qui comprend deux phénomènes plus ou moins imbriqués: la mitose, ou division nucléaire, et la cytodiérèse, ou division cytoplasmique) qui correspond habituellement à moins de 10% de la durée du cycle cellulaire. |
|
Pour un organisme pluricellulaire le lignage de chaque cellule de l'individu adulte a pu être reconstitué notamment chez le vers Nématode Cœnorhabditis elegans qui présente entre une dizaine et une vingtaine d'étapes de divisions successives pour un total de 959 cellules somatiques (voir par exemple fig 14.15, p 510 et s. dans Biologie du développement, Gilbert, 1996, De Boeck Université - ce résultat spectaculaire a été rendu possible par le petit nombre de cellules de cet animal, mais surtout par la transparence de sa cuticule et par la disposition spatiale des territoires cytoplasmiques du zygote qui semble fixe....!); on notera qu'au cours du développement NORMAL de C. elegans 131 cellules subissent une mort naturelle ou apoptose (voir page sur le développement pour une explication de cette notion). |
||||||||||||||||||||||||||||||||||||
| |
|
* De protéines motrices semblent pouvoir se déplacer le long des microtubules en transportant de grosses molécules, des particules protéiques ou des organites. * Les cils et flagelles sont des composés complexes de microtubules associés à des protéiques particulières. * Le début de la division cellulaire peut se repérer dans sa phase la plus précoce par le changement d'organisation du réseau des filaments d'actine du cytosquelette, ce qui se fait en fin de phase G2 pour la cellule des plantes, avant que les chromosomes se condensent et marquent le début de la prophase. |
||||||||||||||||||||||||||||||||||||||
|
3 - Des divisions eucaryotes multiples pour quoi faire ? (complément) |
|
|
||||||||||||||||||||||||||||||||||||||
| |
|
Cette partie est passée du programme de seconde au programme de 1èreS avec des... allégements. Il est clair que le programme présente le phénomène sous un angle extrêmement réducteur : le patrimoine génétique. Selon le programme le rôle d'une division est de transmettre un patrimoine. Ce n'est pas admissible. Il fait aussi l'impasse sur la profonde différence entre les procaryotes et les eucaryotes... on ne s'étonnera plus que les élèves parlent de mitose bactérienne. Ce qui compte c'est l'ADN... quel mensonge ! Un deuxième problème plus subtil est l'utilisation un peu désuète des mots "reproduction conforme". La conformité fait référence à la correspondance avec un modèle ("copie conforme"). Ce qualificatif est donc inapproprié pour désigner deux cellules, non pas conformes à un modèle mais bien héritées d'une cellule mère qui se divise après avoir grandi et les divisions sont bien souvent inégales. L'idée de copie ne peut s'appliquer qu'à la duplication de molécules (comme l'ADN). On pourrait élargir la notion en revenant à l'étymologie : con (cum : avec), forme; la conformité désignant ainsi la TRANSMISSION DE LA FORME lors de la division. Cette voie philosophique me plaît bien davantage. Dans ce cas la conformité serait la transmission des caractéristiques de l'espèce, le volet chromosomique n'en étant qu'un des aspects. |
||||||||||||||||||||||||||||||||||||||
|
Cette partie tente de compenser les manques criants du programme d'une part, en précisant quelques phénomènes cytologiques montrant que la division cellulaire est un phénomène global qui touche toute la cellule et ses organites, et, d'autre part, en replaçant la division cellulaire dans le développement de l'organisme. |
||||||||||||||||||||||||||||||||||||||||
| |
|
3.1 - Les étapes des premières divisions de développement d'un zygote d'Oursin |
||||||||||||||||||||||||||||||||||||||
|
Oursin
méditerranéen |
|
Sur une page du laboratoire de la station de Villefranche sur mer, un film (http://biodev.obs-vlfr.fr/ recherche/ lepage/ images/Film%20oursin%20pwp1.mpg) sur le développement de l'oursin méditerranéen Paracentrotus lividus qui vaut largement les films scolaires commerciaux (dommage que l'on ait pas le timing avec le temps séparant les prises de vues; on peut se référer au site de bmedia: http://www.snv.jussieu.fr/ bmedia/ oursinMDC/index.html et particulièrement à la chronologie : http://www.snv.jussieu.fr/ bmedia/ oursinMDC/timingdev.html ).  Première mitose chez l'Ousin (image externe), MET recolorée ?? |
|
Un site "officiel" pour les enseignants : http://www.snv.jussieu.fr/ bmedia/sommaires/dvpt.htm
|
||||||||||||||||||||||||||||||||||||
|
Film
: pour
diverses
images et
films (en
anglais) sur
les
étapes
de la
fécondation
et du
développement
: http://worms.zoology.wisc.edu/
dd2/echino/cleavage/
mitosis/mitosis.html
chez
l'oursin Lytechinus
variegatus
:
|
|
Ces
vues au
microscope
à
contraste de
phase sont
particulièrement
intéressantes:
on peut noter
|
||||||||||||||||||||||||||||||||||||||
|
Observez les deux premières divisions de l'oursin Paracentrotus lividus en microscopie optique; il faut extraire les 9 premières secondes du film http://biodev.obs-vlfr.fr/ recherche/ lepage /images/Film%20oursin %20pwp1.mpg Divisions égales et synchrones chez Asterina gibbosa (étoile de mer); le film en totalité vaut le coup d'œil :http://www.cerimes.fr/ le-catalogue/ asterina-gibbosa-loeuf-la-larve -et-la-metamorphose-dune- etoile-de-mer.html |
|
La
première
division est
identique
à celle
observée
dans le film
précédant
: le plan du
film contient
bien l'axe de
séparation
des deux
cellules
filles (ou le
fuseau de
division).
L'orientation
est environ
15-20°N-105-110°S. |
|
Si la
seconde
cytodiérèse
semble
intervenir
rapidement,
c'est
probablement
qu'il manque
des images...
L'orientation
du fuseau de
division est
plongeante
obliquement en
arrière
du plan du
film et l'on
ne voit pas
les
phénomènes
fusoriaux. Le
deuxième
fuseau est
donc bien
perpendiculaire
au premier. Le réaccolement intervient ensuite pendant que se prépare une troisième phase de divisions. |
|
Des images sont aussi très parlantes sur le site BIOCLIP, notamment celles de Manuel THERY qui a travaillé sur la physique de la division en orientant celle-ci au moyen d'adhésifs géométriques (chercher Manuel Thery pour voir les vidéos et choisir Randomly oriented cell division). |
||||||||||||||||||||||||||||||||||
| |
|
Mais les divisions suivantes montrent que la multiplication cellulaire est un phénomène complexe qui ne se résume pas à des divisions binaires égales car les divisions ne sont pas indépendantes et la construction de l'embryon est bien plus qu'une somme d'événements tous semblables. |
||||||||||||||||||||||||||||||||||||||
|
Je
recommande un
n°
spécial
de The
International
Journal of
Developmental
Biology,
Vol 50, Nos
2/3, 2006 a
pour titre Morphodynamics. Pour ce qui est de la nature de la matrice de la cellule (cytomatrix en anglais) je recommande un ancien article de K.R. Porter (1984) dont la traduction est disponible ici. ; The Cytomatrix: A Short History of his Study, K.R. Porter, 1984, The Journal of the Cell Biology, vol 99, n°1 |
|
Si l'on observe les divisions suivantes, plusieurs phénomènes peuvent être notés: - il n'y a PLUS de SYNCHRONISATION. - certaines divisions sont INÉGALES . - l'embryon TOURNE plus ou moins dans sa membrane protectrice, ce qui peut être interprété comme l'effet de tensions mécaniques - les fuseaux de deux divisions successives ne sont plus du tout perpendiculaires mais peuvent être plus ou moins parallèles (ce qui amène une croissance de cellules en ligne); c'est ainsi que l'on voit se construire très nettement une couche externe d'assez grandes cellules délimitant une cavité interne (le blastocœle): cette étape est nommée blastulation: l'embryon formé est une blastula. Les divisions successives ne sont donc pas indépendantes, leurs axes sont orientés et elles sont synchronisées ou non, et peuvent être inégales. |
|
On
est donc loin
du
modèle
d'une cellule
(boule) qui
donne deux
cellules
identiques
(boules). Les
divisions
embryonnaires
marquent au
contraire une
construction
ordonnée:
celle de
l'embryon. |
||||||||||||||||||||||||||||||||||||
|
Sur Youtube on peut trouver les films scolaires: commentaire en anglais: http://www.youtube.com/ watch?v=czaO1wYLSDo
Salmon Lab, UNiversity of Carolina, movies |
|
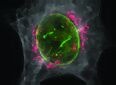
Les films pris au microscope à contraste interférentiel, ont laissé désormais la place aux films présentant un marquage immunologique par plusieurs substances fluorescentes de couleurs différentes qui permettent in suivi IN VIVO des protéines comme la tubuline (en VERT le plus souvent), composant des microtubules, de l'actine (en ROUGE le plus souvent), composant de certains filaments fins de la cellule, ou encore des histones (en BLEU le plus souvent), composant de la chromatine. Cette vision est cependant plus chimique que structurale et il faut se garder d'abandonner les anciennes méthodes. Les nouvelles méthodes doivent compléter les anciennes. |
|
|
||||||||||||||||||||||||||||||||||||
|
voir ancien cours de 1ère S |
|
3.2 - Les étapes de la division cellulaire de croissance d'une cellule de racine de Plante |
||||||||||||||||||||||||||||||||||||||
 Coupe d'une pointe racinaire d'ail (coloration de Feulgen)- Image externe |
|
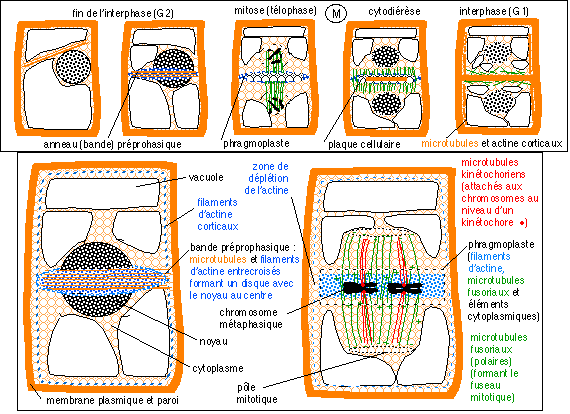
La croissance d'un tissu végétal est réalisée par un accroissement cellulaire (souvent un allongement mais pas toujours; ce qui permet une grande diversité des types de tissus) suivi de la formation d'une cloison (paroi) entre les deux nouvelles cellules. Les phénomènes nucléaires sont ceux d'une mitose avec des caractéristiques particulières, notamment l'absence de centrioles aux deux extrémités du fuseau de division. |
|
Plus la cellule est jeune moins la paroi est épaisse. Lorsque la cellule se différencie se déposent des couches successives de cellulose (glucide complexe formant déja la paroi primaire souple et perméable) qui forment la paroi secondaire. Cette dernière peut s'imprégner de susbtances comme la lignine (polyphénol) qui la rendent rigide et moins perméable. |
|
La grande différence du développement chez les plantes par rapport à celui des animaux réside dans l'absence de déplacement des cellules des plantes qui sont réunies par une paroi sécrétée en permanence et dont la composition change au cours de la vie de la cellule (phénomène qui accompagne la différenciation). La croissance se fait donc dans une direction privilégiée (élongation); l'accroissement en épaisseur des organes étant un phénomène qui intervient dans une phase qui fait suite à la croissance en longueur. |
||||||||||||||||||||||||||||||||||
|
|
|
Division cellulaire d'une cellule de plante (phase M comprenant la mitose et la cytodiérèse). La
fin de
l'interphase
précédant
cette division
et le
début
de
l'interphase
qui la suit,
ont
été
représentées. On retiendra le rôle essentiel des microtubules et des filaments d'actine dans les mouvements déterminant le plan de division, la mitose et la cytodiérèse. Il semblerait cependant que dans le cas de la cellule d'une plante la formation de la plaque cellulaire soit au moins tout aussi importante que la bande préprophasique pour la détermination du plan équatorial de division. |
||||||||||||||||||||||||||||||||||||||
|
|
|
|
||||||||||||||||||||||||||||||||||||||
|
Conclusion |
|
On ne
peut pas
terminer un
chapitre sur
la division
cellulaire
sans parler du
mécanisme
inverse: la fusion
cellulaire.
Deux cellules
peuvent
fusionner en
mettant en
commun leur
cytoplasme et
parfois leur
matériel
génétique. Lorsque l'on expose un phénomène biologique on a toujours tendance à simplifier afin d'en montrer l'universalité. La diversité est alors présentée dans un second temps. Pour la division cellulaire l'unité est mathématique (une séparation : du continu au discontinu, voir page de biologie théorique; c'est d'ailleurs pour moi une voie de recherche des plus prometteuses) mais la diversité prime PARTOUT. Une unité biochimique est tentée avec la molécule d'ADN mais ses limites sont trop grandes. et je regrette fort que le programme bride ainsi l'esprit des élèves. |
||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||