|
K.R. Porter, 1984, The Journal of
the Cell Biology, vol 99, n°1
|
||||||
|
Traduction personnelle mot-à-mot non garantie |
|
|
||||
|
|
||||||
|
Dès 1820 et pendant plusieurs décennies, les premiers observateurs de la matrice cellulaire ont travaillé principalement avec le microscope optique et un matériel très simple. Ce n'était pas rare pour le chercheur d'écraser tout simplement une cellule entre deux lames et de regarder les résultats. La seule motivation pour ces études, pour autant que je puisse en juger, était la satisfaction de la curiosité, car il n'y avait pas de subventions. Ces observations sont précieuses pour nous aujourd'hui, en particulier celles qui s'accompagnent d'interprétations imaginatives. Les études sur la matrice cellulaire peuvent se scinder approximativement en trois périodes, initiale, intermédiaire et récente. La première période, qui fut essentiellement descriptive, est la plus longue et correspond aux années 1820 à 1910. C'était le temps où microscopes optiques se sont grandement améliorés, lorsque les fixateurs chimiques ont été découverts, lorsque les microtomes et les procédures de coloration ont été introduits, et enfin quand la communauté des biologistes a pris conscience de la notion de cellule, de comment celle-ci se divisait et s'assemblait en tissus. La pathologie cellulaire fût également lancée au cours de cette période. La période intermédiaire a commencé autour de 1910 et a continué jusqu'en 1940. L'accent a été mis dans cette période sur l'étude des cellules vivantes et sur des expériences conçues pour répondre à des questions fondamentales concernant les propriétés de la matrice cellulaire et des bases structurelles de l'organisation intracellulaire. La période récente a vu l'introduction de la microscopie à contraste de phase, ce qui a grandement facilité l'observation des cellules vivantes. Elle a été aussi, bien sûr, été dominée par la méthode du fractionnement cellulaire et ma microscopie électronique. Cela fut une période d'activité intense, où l'on obtint une grande quantité d'informations sur les cellules et les tissus. La biologie cellulaire est maintenant considérée comme une science distincte, mais la matrice cytoplasmique n'est pas encore perçue par tous les biologistes cellulaires comme quelque chose d'intéressant à étudier. Je voudrais commencer ma discussion avec la période intermédiaire, puis revenir sur la première période de son arrière-plan, pour finir par la période récente et la plus familière. La place ne me permettant pas de mentionner tous les détails que je voudrais. |
|
As early as 1820 and
for several decades thereafter the early observers of
the cytomatrix worked mostly with the light microscope
and very simple equipment . It was not uncommon for the
investigator simply to crush a cell between two slides
and watch the results . The sole motivation for
these studies, as far as I can judge, was the
satisfaction of curiosity, for there were no grants .
Such observations are valuable to us today, especially
those accompanied by imaginative interpre-tations. |
||||
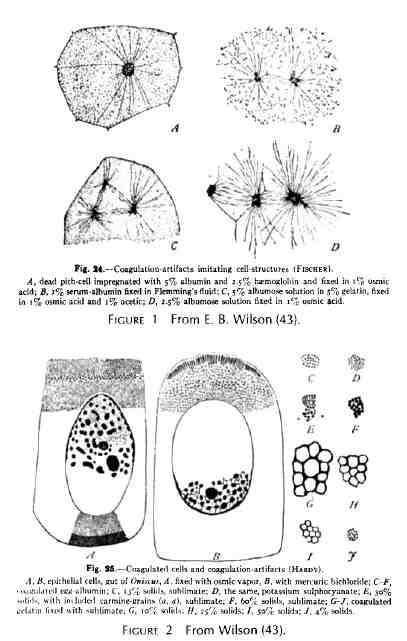
FIGURE 1 - De E, B. Wilson (43) FIGURE 2 De Wilson (43)
|
|
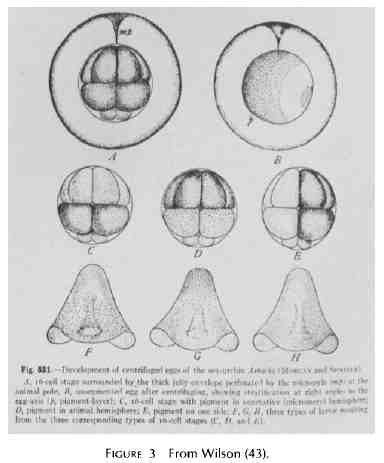
Figure 2 - De Wilson (43)
|
||||
|
La période intermédiaire Du point de vue d'un biologiste, la période moyenne est peut-être la plus intéressante. A. Fischer et W. B. Hardy, travaillant séparément, ont surpris la communauté des microscopistes optiques en publiant des preuves montrant que les élégants dessins de leurs prédécesseurs étaient pleins d'artefacts, tous produits par la coagulation pendant la fixation et/ou la déshydratation (15, 17). Des cytologistes antérieurs, tels que Flemming, Berthold et Bütschli, avaient déjà mis en garde leurs contemporains du risque de confusion entre les coagulations et les structures natives. Fischer et Hardy montrèrent des photos de dessins d'asters et de fuseaux trouvés dans les cellules mortes de moelle imprégnées avec de l'albumine (Fig. 1). Pour Hardy, la structure alvéolaire de la matrice cellulaire, le spongioplasme, n'était rien d'autre qu'une conséquence de la fixation (Fig. 2). Il se peut que ces attaques sur les découvertes que nous avons maintenant être fondées aient induit dans l'esprit des cytologistes et des physiologistes suffisamment de scepticisme pour les encourager une plus grande attention dans leurs études de cellules vivantes. En tout cas, il a suivi une période de vif intérêt dans l'étude de protoplasme vivant. Robert Chambers a été une figure dominante dans cette tendance. Je doute qu'il n'ait jamais fixé lui-même une cellule ou réalisé une section. Il préférait effectuer de la microchirurgie sur les cellules et puis leur injecter des corps étrangers, en particulier des huiles et observer les conséquences. Au début de sa carrière, il est allé à Cambridge, en Angleterre, pour travailler sur des cellules en culture avec Honor. B. Fell. Dans une expérience, ils ont pu montrer que la l'insertion d'une microaiguille dans le noyau provoque son éclatement, puis la lyse de la cellule entière. Si, toutefois, la cellule avait deux noyaux et qu'ils n'en détruisent qu'un seul, l'autre survivait, comme l'ensemble de la cellule (8). L'utilisation d'aiguilles lors de différentes étapes de la vie cellulaire devint très populaire. G. W. Scarth, un botaniste de l'Université McGill, a observé que le noyau de Spirogyra peut être déplacé de plusieurs micromètres, puis lorsqu'il est relâché, il reviendrait rapidement à sa position d'origine. Certaines propriétés viscoélastiques de la matrice ont donc été supposées (37). G. L. Kite était un autre passionné de micromanipulation. Quand il a placé une aiguille entre le pronucléus mâle et le pronucléus femelle d'un œuf de Toxopneustes récemment fécondé, il a découvert qu'il pouvait pousser un des noyaux, mais que celui-ci continuait à glisser loin de l'aiguille et à avancer vers son compagnon (7). Encore une fois, un mécanisme de la matrice semblait jouer. Chambers a fait un examen assez systématique de ce qui pouvait et ne pouvait pas être manipulé parmi les composants structuraux cellulaires. Il a constaté des différences frappantes dans la capacité des structures à résister à ses interventions. Il a trouvé, par exemple, que « les brins fibreux, les vésicules et les mitochondries en forme de bâtonnet peuvent être déplacés et perturbés sans perte apparente de leur intégrité. Alors que d'autres structures, telles que l'aster, lorsqu'elles sont piquées, disparaissent rapidement, et que "l'état physique du protoplasme ressemble à celui d'un système colloïdal de type sol-gel réversible"(7). Il a aussi découvert que la transformation du gel en sol, avec disparition de certains éléments structuraux, peut être induite par différents procédés expérimentaux, en particulier ceux qui impliquent l'utilisation de la pression hydrostatique ou une soudaine agitation mécanique. Nous reviendrons sur ce sujet ultérieurement. Il est également important de se rappeler certains travaux de Chambers en collaboration avec M. J. Kopac impliquant l'injection de gouttelettes d'huile dans les œufs d'oursin (7, 24). Ordinairement, ces corps graisseux conservent une forme sphérique. Cependant, quand ils sont placés à l'intérieur "de l'aster radialement gélifié d'un ovule fécondé, ils adoptent une forme ovoïde". De toute évidence, l'organisation structurelle de l'aster est suffisamment rigide pour déformer la goutte d'huile. Kopac (24) a écrit que les gouttes d'huile insérées soigneusement dans les ovocytes immatures d'Asterias restent sphériques et bien délimitées tant que l'huile ne provoque pas de dommage sensible au cytoplasme, mais « qu'une réaction marquée se produit, quand, après que l'huile soit introduite, la cytolyse est induite. Dans les 30 s après la cytolyse de la cellule une membrane de protéines adsorbées apparaît à l'interphase huile-cytoplasme et qui se plisse (c'est l'effet Devaux) quand une partie de l'huile est retirée". A ce propos, Chambers (7) écrit :« Comme le plissement de l'effet Devaux de la goutte d'huile avec la méthode de rétraction de l'huile n'est pas perceptible dans le cytoplasme vivant, on peut en conclure que les protéines de la cellule vivante ne s'accumulent pas sur des surfaces introduites expérimentalement lorsque le protoplasme est intact. Ceci suggère que les protéines ne sont pas librement diffusibles et adsorbables dans le protoplasme, et que, par conséquent, ces protéines peuvent être liées ensemble pour former une sorte de phase continue", et il poursuit en disant que : " le meilleur argument en faveur de l'existence d'une couche différenciée à la surface du protoplasme est probablement le fait qu'une solution colorée qui ne peut pas entrer de sans la volonté humaine, est capable, lors de micro-injections, de se répandre à l'intérieur de la cellule, mais sans en ressortir. "Apparemment, le colorant est confiné dans une phase riche en eau. Les discussions sur l'existence de l'organisation des cellules - et notamment des cellules-oeufs - étaient monnaie courante, en particulier dans la littérature émanant du Laboratoire de biologie marine de Woods Hole. Les noms de Conklin, Lillie, et Boveri sont intimement associés au travail effectué principalement dans le but de déterminer si les substances appelées déterminants (organ-forming) dans l'ooplasme étaient distribuées de façon non aléatoire et immobilisées. Ils trouvèrent que l'on pouvait sédimenter par centrifugation ces substances intracellulaires, comme le jaune et le pigment des œufs d'Arbacia, sans que les étapes de la segmentation ne soient modifiées. En d'autres termes, une partie de l'ooplasme qui ne se déplace pas avec le pigment a participé à la formation des composants des asters et rayons et contribué à la composition des micromères et des autres cellules du stade 16 cellules, comme si l'œuf n'avait jamais été centrifugé (11, 40) (Fig. 3). Comme E. B. Wilson (43) le souligne, « il y a une évidente difficulté à concevoir une organisation prédéterminée de l'œuf reliée à une substance liquide ou semi-liquide, comme cela semble souvent être fait avec le hyaloplasme. Lillie et Conklin ont aussi plaidé en faveur d'un état relativement ferme des substances agrégées dans le hyaloplasme, même si les inclusions cytoplasmiques devaient pouvoir encore se déplacer en son sein. Ainsi des "mouvements de flux", que ce soit dans l'œuf normal ou résultant de la centrifugation, sont considérés comme n'étant que des mouvements granulaires au sein de réseau semi-solide de l'ooplasme. Conklin a produit une remarquable preuve de ce qu'un tel réseau stable de protoplasme plus visqueux (hyaloplasme) existe dans la substance claire de l'œuf de Crepidula, et qu'il constitue la base des activités vraiment localisées . Cette substance est considérée par Conklin comme étant élastique et contractile, et donc capable de produire les soi-disant mouvements fluides du protoplasme vivant et et de son contenu granulaire. Il souligne le fait, également noté par les observateurs précédents, que si la centrifugation est réalisée suffisamment précocement et n'est pas trop longue les composants séparés des œufs ont tendance à revenir plus ou moins complètement à leur position d'origine ; et ceci serait aussi dû à l'action du réseau contractile qui est resté inchangé dans sa polarité originelle." |
|
Middle Period From the viewpoint of a biologist, the middle period is perhaps the most interesting . A . Fischer and W . B. Hardy, working separately, had just startled the community of light microscopists by publishing evidence claiming to show that the elegant drawings made by their predecessors were full of artifacts, all products of coagulation during fixation and/or dehydration (15, 17) . Earlier cytologists, such as Flemming, Berthold, and Butschli, had already cautioned their contem- poraries of the danger of confusing coagulations with genuine structures . Fischer and Hardy provided in their drawings images of asters and spindles found in dead pith cells impreg- nated with albumin (Fig. 1) . For Hardy, the alveolar structure of the cytomatrix, the spongioplasm, was nothing more than a consequence of fixation (Fig. 2) . It may be that these attacks on findings that we now know were meaningful induced enough skepticism in the minds of cytologists and physiologists to encourage greater emphasis on studies of living cells. In any case, there followed a period of intense interest in the study of living protoplasm . Robert Chambers was a dominant figure in this trend. I doubt that he ever fixed a cell or cut a section. He preferred instead to perform microsurgery on cells and later to inject foreign bodies, particularly oils, into cells and observe the various consequences . Early in his career he went to Cambridge, England, to work on cultured cells with Honor B . Fell . In one experiment, they were able to show that the thrust of a microneedle into the nucleus induced its breakdown and, subsequently, the lysis of the whole cell . If, however, the cell had two nuclei and they destroyed only one, the other sur- vived, as did the whole cell (8) . Needle intervention in the life of cells became quite popular . G . W. Scarth, a botanist at McGill University, observed that the nucleus of Spirogyra could be displaced several micrometers and that when released it would return promptly to its original position . It appeared that some viscoelastic property of the matrix asserted its influence (37) . G . L . Kite was another enthusiast of micro- manipulation . When he interposed a needle between the male and female pronuclei of a recently fertilized Toxopneustes egg he found that he could push one of the nuclei about but that it persisted in slipping off the needle to advance toward its mate (7) . Again, a mechanism of the matrix appeared to be at work . Chambers made a fairly systematic examination of what could and could not be manipulated among the structural components of cells . He found striking differences in the ability of structures to withstand his interventions. He found, for example, that "fibrous strands, vesicles and rod-shaped mitochondria may be moved about and disturbed with no apparent loss of integrity. On the other hand, other structures, such as the aster, when prodded quickly disappear" and that "the physical state of the protoplast resembles that of a reversible sol-gel colloidal system" (7) . He also discovered that the transformation from gel to sol, with the subsequent dis- appearance of structural features, can be induced by various experimental procedures, particularly those that involve the use of hydrostatic pressure or sudden mechanical agitation . I shall return to this topic below. It is important also to recall some work that Chambers did with M . J . Kopac involving the injection of oil droplets into sea urchin eggs (7, 24) . Ordinarily, these fat bodies retain a spherical form . When, however, they are placed within "the radially gelated aster of a fertilized egg, they adopt an ovoid form ." Clearly, the structural organization of the aster is sufficiently rigid to distort the oil drop . Kopac (24) wrote that drops of oil inserted carefully into the immature oocytes ofAsterias remain spherical and sharply defined as long as the oil causes no perceptible injury to the cytoplasm but that "a pronounced reaction occurs, when, after the oil is introduced, cytolysis is induced . Within 30 s after the cell has cytolyzed a membrane of adsorbed protein appears at the oil-cytoplasmic interphase and this is observed to crinkle (the Devaux effect) when some of the oil is re- trieved." Regarding this, Chambers (7) writes : "Since the Devaux crinkling effect of the oil drop with the oil-retraction method is not noticeable in the living cytoplasm, it is con- cluded that the proteins in the living cell do not accumulate on experimentally introduced surfaces while the protoplasm is intact. This suggests that the proteins are not freely diffusible and adsorbable in protoplasm, and that, therefore, these pro- teins may be bound together to form some kind of continuous phase," and goes on to say that "Probably the strongest argument for the existence of a differentiated layer on the surface of protoplasm is the fact that a colored solution which cannot enter from without will, when micro-injected, spread through the interior but will not pass out of the cell ." Appar- ently, the dye is confined to a water-rich phase. Discussions of the existence of organization in cells-in particular egg cells-were common, especially in the literature emanating from the Marine Biological Laboratory in Woods Hole . The names of Conklin, Lillie, and Boveri are intimately associated with work done principally to determine whether the so-called organ-forming substances in ooplasm are distrib- uted nonrandomly and fixed in their positions . They found that although such intracellular substances as the yolk and pigment of Arbacia eggs could be stratified by centrifugation, the pattern of cleavage was not altered. In other words, a part of the ooplasm that did not move with the pigment went into the formation of the ingredients of asters and spindles and contributed to the content of micromeres and other cells of the 16-cell stage as though the egg had never been centrifuged (11, 40) (Fig . 3) . As E . B . Wilson (43) points out, "The difficulty of conceiv- ing how the prelocalized organization of the egg can be bound up in a liquid or semi-liquid substance, such as the hyaloplasm often seems to be, is obvious . Lillie and Conklin have accord- ingly argued in favor of a relatively firm condition of aggre- gation in the hyaloplasm, yet one of such a nature that the cytoplasmic inclusions can still move through it . `Flowing movements,' accordingly, whether in the normal egg or pro- duced by the centrifuge, are regarded as no more than granule- movements within this semi-solid framework of the ooplasm . Conklin has produced considerable specific evidence that such a persistent framework of more viscid protoplasm (hyaloplasm) exists in the clear substance of the Crepidula egg, and that it forms the basis of the true localizing activities. Its substance is assumed by Conklin to be elastic and contractile, and thus to produce the so-called flowing movements of living protoplasm and its contained granules . He emphasizes the fact, also noted by earlier observers, that if the centrifuging acts at a sufficiently early period and is not too long continued the dislocated egg-components tend to return more or less completely to their original position ; and this too is ascribed to the action of the contractile framework which has main- tained unchanged its original polarity."
|
||||
|
|
||||||
|
Ces études nous rappellent une des expériences effectuées un peu plus tard par H. W. Beams et R. L. King (1, 2). Ils exposèrent une grande variété de cellules à d'énormes forces centrifuges (400.000 g) et recherchèrent des preuves de survie. Cette expérience fût réalisée dans une petite centrifugeuse à entraînement par air développée par J. W. Beams, un frère de H. W. Beams. Le contenu de l'œuf d'Ascaris s'est stratifié en trois couches après 1 h à 400.000 g et est ensuite retourné à l'organisation initiale en 12 heures. À la fin des 48 h, 90% des œufs centrifugés s'étaient divisés. D'autres oeufs et des stades précoces de la segmentation centrifugés à 150.000 g pendant 4,5 jours ont survécu et se sont développés à la vitesse normale. Dans d'autres expériences, les œufs se sont segmentés durant leur centrifugation à 100.000 g. Apparemment, tout déplacement de la matrice cellulaire ou de son contenu réalisé à 100.000 g n'est pas suffisant pour perturber l'organisation du cytoplasme en empêchant la division cellulaire. Avec des forces supérieures un temps de récupération semble nécessaire avant une segmentation normale. Peut-être y-a-il quelque chose de vrai dans le fait d'appeler la matrice cellulaire le "spongioplasme." L'organisation nécessaire à la mitose est extraordinaire, et des changements de conditions relativement simples sont connues pour la perturber. Ainsi, comme nous le savons par des études plus récentes par Inoué et ses collaborateurs (21), la biréfringence du fuseau mitotique disparaît à basse température et que les chromosomes s'éloignent de leur position sur la plaque métaphasique. La chose étonnante est qu'ils retournent à leur position alignée sur la biréfringence lorsque les cellules reviennent à la température normale. C'est comme si une sorte de mémoire organisationnelle basique survivait à l'expérience et guidait la reconstruction du fuseau. Dans cette période intermédiaire, Marsland introduit et expérimenta de façon exhaustive les caractéristiques de la matrice à l'aide de pressions hydrostatiques. Ces expériences ont fait suite à la démonstration par Dugald Brown que le cytoplasme central de l'œuf d'Arbacia est relativement fluide par rapport à uns épaisse zone corticale (5 µm). Exposé à un faible champ centrifuge, les « composants granulaires » du cytoplasme central ont été facilement déplacés, tandis que ceux du cortex ne l'étaient pas. Lorsque la même expérience a été réalisée avec des niveaux croissants de pression hydrostatique, le cortex présente une liquéfaction qui augmente. À 10.000 p.s.i. le précédent gel cortical était presque indétectable. Des études ultérieures ont montré que la pression hydrostatique provoquait la liquéfaction de gels protoplasmiques dans un grand nombre de cellules (27, 30). Nous savons maintenant, bien sûr, comme l'ont démontré Tilney et coll. (42), que le désassemblage des microtubules est partiellement responsable de ce phénomène. L'effet est réversible et implique sans doute la dissolution d'autres composants de la cytomatrice que les microtubules. On considère que la transformation sol-gel représente un comportement similaire des composants de la matrice dans des conditions normales. Certaines observations de Pease (30) sur des œufs du nématode Rhabditis in utero sont pertinentes. Il a trouvé que le mouvement brownien, habituellement très limité, augmente très fortement dans les œufs centrifugés : « des forces centrifuges modérées solubilisent le réseau du gel, et le mouvement brownien n'est plus empêché, jusqu'à ce que le gel se forme lentement à nouveau, le processus prenant quelques minutes » [voir aussi Kitching et Pease (23)]. |
|
These studies remind one of experiments performed some- what later by H . W . Beams and R . L . King (1, 2). They exposed a wide variety of cells to huge centrifugal forces (400,000 g) and watched for evidence of survival . This was done in a small air-driven centrifuge developed by J . W. Beams, a brother of H . W. Beams . The contents of Ascaris eggs stratified into three layers during 1 h at 400,000 g returned to normal distribution over the next 12 h . At the end of 48 h, 90% of the centrifuged eggs had divided . Other eggs and early cleavage stages centrifuged at 150,000 g for 4 .5 d survived and developed at the normal rate . In other experiments, eggs were seen to undergo cleavage while centrifuged at 100,000 g. Apparently, any displacement of the cytomatrix or its contents achieved at 100,000 g is not suffi- cient to disrupt the organization of the cytoplasm essential for cell division . At the higher forces an interval of recovery appears to be essential for normal cleavage. Perhaps there is something in the concept that characterizes the cytomatrix as "spongioplasm ." The organization essential for mitosis is awesome, and variations in relatively simple conditions have been observed to disrupt it . Thus, as we know from more recent studies by Inou6 and his collaborators (21), the birefringence of the mitotic spindle disappears at low temperatures and the chro- mosomes drift away from their position at the metaphase plate . The amazing thing is that they return to their normal position along with the birefringence when the cells are re- turned to normal temperature . It is as though some basis of organizational memory survives the experiment and guides the restructuring of the spindle . In this middle period Marsland introduced and exhaustively used hydrostatic pressures to explore the characteristics of the matrix . These experiments followed the demonstration by Dugald Brown that the central cytoplasm of the Arbacia egg is relatively fluid compared with a thick (5-tum) cortex . When exposed to a weak centrifugal field, the "granular compo- nents" of the central cytoplasm were readily displaced, whereas those in the cortex were not. When the same experi- ment was performed with increasing levels of hydrostatic pressure, the cortex displayed increasing liquefaction . At 10,000 p .s .i . the erstwhile cortical gel was almost undetectable. Subsequent studies showed that hydrostatic pressure induces solation of protoplasmic gels in a wide variety of cells (27, 30) . We now know, of course, as first demonstrated by Tilney et al . (42), that microtubules disassemble as part of this phenomenon . The effect is reversible and probably involves solation of more than microtubule components of the cyto- matrix. The sol-gel transformation is thought to represent a similar behavior of matrix components under normal condi- tions. Some observations by Pease (30) on the in utero eggs of the nematode Rhabditis are pertinent . He found that Brownian motion, ordinarily very limited, increased dramat- ically in centrifuged eggs: "moderate centrifugal forces solated the gel network, and Brownian movement was unrestricted until the gel slowly reformed, the process taking some minutes to be completed" [see also Kitching and Pease (23)] . |
||||
|
|
||||||
|
Plusieurs chercheurs, à commencer par Heilbronn en 1922 (19), puis Heilbrunn (20), jusqu'à Crick et Hughes en 1950 (12), ont tenté d'apprendre quelque chose sur l'élasticité de la matrice en introduisant des particules de fer dans le cytoplasme, puis en les manœuvrant à l'aide de champs magnétiques externes (20). Ces expériences semblent être faciles, mais selon Crick et Hughes les résultats ont été difficiles à quantifier. Ils ont réussi dans une certaine mesure, là où d'autres avaient échoué, en prenant des films qu'ils pouvaient ensuite analyser (12). Ce travail a été résumé ailleurs (35), ainsi : « À chaque fois qu'une particule était déplacée, il y avait un petit recul rapide lorsque le champ magnétique était mis hors tension. Le recul ne ramène jamais la particule magnétique à son point de départ; généralement le recul était d'environ un tiers de la distance originellement parcourue. Lorsqu'ils étaient répétés, les déplacements de particules induits magnétiquement semblent générer un plus grand espace de mouvement libre; c'est-à-dire que huit répétitions de mise-en-marche-arrêt du champ ont réduit la viscosité dans l'espace où les particules ont été déplacées. Ils [Crick et Hughes] en ont conclu que le cytoplasme est un gel thixotrope. Le recul limité pourrait être interprété comme le reflet de l'élasticité d'une zone de compression qui se développe dans un maillage filamenteux à l'avant de la particule en mouvement. La distance traversée moins le recul peut représenter une partie de la matrice dans laquelle la structure a été endommagée et la viscosité nettement altérée (un effet thixotrope). Bien évidemment, ces expériences comme d'autres décrivent la présence d'une matrice structurée et visqueuse dans le cytoplasme, une matrice avec une élasticité (ou une information structurelle) suffisante pour la ramener à sa forme initiale après différentes perturbations. L'ultramicroscope (fond noir) introduit au cours de la période intermédiaire a permis d'intéresser Strangeways et Canti (41) à la présence de petites structures réfringentes dans les régions les plus centrales des cellules en culture ainsi qu'à l'évidente absence de mouvement brownien dans une cellule normale indemne. Ils furent aussi les premiers à montrer que l'image n'était pas perceptiblement modifiée par la fixation des cellules avec des vapeurs de OsO4, ce qui n'était pas le cas pour les autres fixateurs. La biréfringence des cellules et en particulier de certains composants cellulaires a été explorée en premier par W. J. Schmidt (39). Des systèmes fibreux dans le cytoplasme ont été reconnus grâce à leur biréfringence, comme l'ont été plus tard les structures riches en microtubules, comme les axonèmes, les cils et les fuseaux mitotiques. Le flux de la biréfringence était assez évident, ce qui a conduit à de nombreuses discussions sur l'écoulement non newtonien du cytoplasme visqueux. Je ne dois pas mettre fin à mon analyse de cette période sans mentionner deux théoriciens, A. Frey-Wyssling et G.W. Scarth. Le premier était surtout expert dans le "joyeux mélange". Par exemple, il disait à propos des propriétés physiques du cytoplasme: « Le paradoxe du cytoplasme, c'est qu'il montre à la fois une fluidité et une élasticité. C'est un solide et un liquide en même temps dans une autre mesure de celle observée pour d'autres colloïdes. La tâche de morphologie submicroscopique consiste, par conséquent, à élaborer un schéma structural » (16). Son schéma montre la matrice comme un réseau extrêmement fin, dont les mailles contiennent des substances interstitielles comme l'eau, des sels et du glucose en solution. La différence fondamentale entre gels inanimés et vivants est que dans le cytoplasme vivant les jonctions entre filaments sont continuellement restructurées. Scarth (38) compare la structure à celle d'une brosse et dit que « Le mystère de l'organisation cellulaire, peut-être plus que tout autre problème biologique, semble reposer sur un point de vue vitaliste. Cependant, comme Driesch a fait remarquer, s'il pouvait être prouvé que l'organisation peut exister sans support physique, une explication mécaniste deviendrait ipso facto impossible." "Le type d'organisation qui est expérimentalement trouvé dans les œufs et d'autres cellules est celui de la polarité, de la localisation des substances et, dans le noyau, d'un arrangement en unités ordonnées linéairement. Ceci nécessite plus ou moins une base strucuturale. Ainsi, si elle devait se déployer dans un milieu liquide - comme certains qualifient le protoplasme - nous serions en face d'un miracle. |
|
Several investigators, beginning with Heilbronn in 1922 (19), including Heilbrunn (20), and ending with Crick and Hughes in 1950 (12), attempted to learn something about the elasticity of the matrix by introducing iron particles into the cytoplasm and then shunting them around with externally applied magnetic fields (20) . The experiments sound like fun, but according to Crick and Hughes the results were difficult to quantitate . They succeeded to some degree, where others had not, by taking movies that they could later analyze (12) . This work has been summarized elsewhere (35), as follows : "In each case, when a particle was moved there was a small, rapid recoil when the magnetic field was turned off. The recoil never returned the magnetic particle to its starting point ; usually recoil was about one-third of the distance originally traveled . Repeated, magnetically induced excursions of par- ticles seemed to generate a larger space for unimpeded motion ; ie, eight reiterations of the on-and-off action of the field lowered the viscosity in the space through which the particles had moved . They [Crick and Hughes] concluded from this that the cytoplasm is a thixotropic gel . The limited recoil one might interpret as reflecting the elasticity of a zone of compres- sion that develops in a filamentous meshwork immediately in advance of the moving particle . The distance traversed minus the recoil could represent a part of the matrix in which the structure was damaged and the viscosity markedly altered (a thixotropic effect) . Quite obviously, these experiments and others describe the presence of a viscous and structured matrix in the cytoplasm, a matrix with enough elasticity (or structural information) to return it to its undistorted form after some kinds of perturbations ." The ultramicroscope (dark field) introduced in the middle period helped to alert Strangeways and Canti (41) to the presence of small refractile structures in the more central regions of thinly spread cultured cells and also to the striking absence of Brownian motion in a normal uninjured cell . They were original also in showing that the image was not detectably altered by fixing the cells with vapors of OsO,, which was not true of other fixatives. The birefringence of cells and especially of certain cell components was pioneered by W . J . Schmidt (39) . Fiber systems in the cytoplasm became known for their form bi- refringence, as did later structures rich in microtubules, such as axonemes, cilia, and mitotic spindles . Flow birefringence was fairly apparent, and this initiated much discussion of non- Newtonian flow as displayed by the viscous cytoplast . I should not end my discussion of this period without mentioning two theorists, A . Frey-Wyssling and G. W. Scarth . The former was especially expert in adding it all up. For example, he says of the physical properties of cytoplasm : "The paradox of the cytoplasm is that it shows both fluidity and elasticity. It is a solid and a liquid at the same time to an extent not observed in any other colloid . The task of submi- croscopic morphology consists, therefore, of drawing up a structural schema" (16) . His scheme shows the matrix as an extremely fine network, the meshes of which contain such interstitial substances as water and salts and glucose in solu- tion . The fundamental difference between nonliving and liv- ing gels is that in living cytoplasm the junctions between the filaments are continuously restructured . Scarth (38) likens the structure to that of a brush heap, and says that "The mystery of cell organization, more perhaps than any other biological problem, seems to conduce a vital- istic viewpoint . However, as Driesch pointed out, if it can be proved that organization may exist without an adequate phys- ical mechanism, a mechanistic explanation of it becomes, ipso facto, impossible . "The kind of organization that is experimentally discovered in eggs and other cells is that of polarity, localization of substance and, in the nucleus, an orderly serial arrangement of units . For this to exist physically demands a more or less structural basis . Consequently, if it should be displayed by a liquid medium such as protoplasm is said to be-at least in some crucial instances-we are faced with a biological miracle . |
||||
|
|
||||||
|
Scarth (37) a pris part à un programme organisé en 1940 par la Société américaine de physiologie végétale (American Society of Plant Physiologists) (Fig. 4). À la fin de son discours, il a récité une parodie d'un obscur vers écrit par John Godfrey Saxe un siècle plus tôt, "Les aveugles et l'éléphant."
Who went to see the Protoplast (though all of them were blind) That each its structure might observe to satisfy his mind. The first advancing hurriedly and happening to fall Right through its soft interior at once began to bawl "God bless me! But the Protoplast is very like a Sol." The second poked the animal and felt his staff repel Its tough and springy cortex, so he began to yell "Tis evident the Protoplast is very like a gel." The third approaching gingerly did only pinch and squeeze Its slippery oleaginous hide when he began to wheeze "It seems to me the Protoplast is just a lump of grease." The fourth man, having punched and probed and proved its plastic state, Watery yet indissoluble, did thus asseverate "The Protoplast is a compound, complex co-a-cerv-ate ." And so these fundamentalists disputed loud and long Each in his own opinion exceedingly stiff and strong, Though each was partly in the right and all of them were wrong . D'autres ouvrages ont rapporté les synthèses réalisées dans le but de résumer ce qui avait été appris dans les années 30 avant que la Première Guerre mondiale ne perturbe la vie normale et de la recherche. Un de ces ouvrages qui résume le mieux les progrès de la période intermédiaire comprend 31 essais présentés à Sir Frederick Gowland Hopkins (1937) lors de son 75e anniversaire. Un des essais était de Joseph Needham, l'auteur de l'Embryologie Chimique (29). Il se démarque de la chimie au point de dire "qu'un véritable mouvement a eu lieu au cours des dernières années dans le sens d'une base de l'organisation protoplasmique à partir de molécules fibreuses ou filamenteuses, ce qui rend plus facile de concevoir une structure tridimensionnelle en cours de construction par des fibres orientées plutôt que par des sphères cohésives, ce qui est fondamental pour la morphologie. " Ainsi on peut aisément imaginer que le chemin se fasse vers le terme de "squelette cellulaire." En fait, il avait utilisé le terme cytosquelette pour la première fois dans ses "Terry lectures" un an plus tôt. Sir Rudolph Peters (31) dans son article a exprimé son impatience vis-à-vis des « conceptions actuelles purement colloïdales de la cellule." Il erre dans des analogies fantaisistes, comme celle de l'existence d'un système nerveux intracellulaire composée d'une certaine façon de protéines cytoplasmiques, et finit par un plaidoyer pour l'étude des cellules intactes. Le réticulum endoplasmique lisse et son contrôle du Ca libre" rejoint aujourd'hui la notion de système nerveux intracellulaire de Peters. Malheureusement, la Seconde Guerre mondiale interrompit en Angleterre et ailleurs un grand nombre des recherches qui auraient été très utiles pour comprendre la nature du cytosquelette. |
|
Scarth (37) took part
in a program organized in 1940 by the American Society of
Plant Physiologists (Fig . 4) . At the end of his talk he
recited a parody on a little light verse written by John
Godfrey Saxe a century earlier, "The Blind Men and the
Elephant." It was four fundamentalists to learning much
inclined, Who went to see the Protoplast (though all of them
were blind) That each its structure might observe to satisfy
his mind. The first advancing hurriedly and happening to
fall Right through its soft interior at once began to bawl
"God bless me! But the Protoplast is very like a Sol." The
second poked the animal and felt his staff repel Its tough
and springy cortex, so he began to yell "Tis evident the
Protoplast is very like a gel." The third approaching
gingerly did only pinch and squeeze Its slippery oleaginous
hide when he began to wheeze "It seems to me the Protoplast
is just a lump of grease." The fourth man, having punched
and probed and proved its plastic state, Watery yet
indissoluble, did thus asseverate "The Protoplast is a
compound, complex co-a-cerv-ate ." And so these
fundamentalists disputed loud and long Each in his own
opinion exceedingly stiff and strong, Though each was partly
in the right and all of them were wrong . Other volumes grew
out of conferences in response to the urge to sum up what
had been learned in the 30 years before World War 11
disrupted normal life and research . One of these volumes
that summarized better than others the progress of the
middle period comprised 31 essays presented to Sir Frederick
Gowland Hopkins (1937) on his 75th birthday . One essay was
by Joseph Needham, the author of Chemical Em- bryology (29)
. He departs from chemistry enough to say "that a whole
movement has been taking place in recent years towards the
conception of fibre or thread molecules as the basis of
protoplasmic organization and since it is easier to conceive
of a three-dimensional structure being built of ori- ented
fibers than of cohering spheres the result is of no small
importance for morphology ." From this point one can easily
imagine him working his way to the term "cell skeleton ."
Actually, he had used the term cytoskeleton for the first
time in his Terry lectures a year earlier . Sir Rudolph
Peters (31) in his paper expressed impatience with "the
current purely colloidal conceptions of the cell." He
wandered into fanciful analogies, e .g., that of the
existence of an intracellular nervous system composed in
some way of cytoplasmic proteins, and emerged at the end
with a plea for the study of intact cells . The smooth
endoplasmic reticulum and its control of free Ca" would
coincide today with Peters' notion of an intracellular
nervous system . Unfortunately, World War II interrupted in
England and elsewhere much of what might have been very
valuable in- vestigations into the nature of the
cytoskeleton . Page de titre d'une monographie réalisée sur la base des documents présentés lors d'un symposium parrainé par l'American Society of Plant Physiologists qui s'est tenue à Philadelphie le 30 Décembre 1940. |
||||
|
|
||||||
|
La période initiale Je vais maintenant remonter aux tout débuts (la période initiale) et je serai bref. Il est difficile d'identifier les premières observations ou les plus significatives au sujet de la matrice optiquement vide de la cellule. Elle a peut-être attiré les yeux perçants de Van Leeuwenhoek dès 1670. Les observations significatives, cependant, ont du attendre le développement de meilleurs microscopes et, notamment, des lentilles achromatiques. Beaucoup ont été attirés par l'utilisation de ces nouveaux microscopes dans la première moitié du 19e siècle. Les noms de quelques-uns sont connus: C. F. Wolff (1733-1794), B. De Mirbel (1776-1854), Lamarck (1744-1829), Dutrochet (1824), J. P. F. Turpin (1775-1840), Meyen (1804-1840), von Mahl (1805-1872), Brown (1773-1858). Ils ont tous, par leurs observations sur la structure cellulaire des plantes et des animaux, précédé Schleiden et son Beitrage de 1838 de plusieurs années. Félix Dujardin (1801-1860) est allé plus loin que la plupart des autres avec ses grossières tentatives d'apprendre quelque chose sur la nature physique du protoplasme. Il l'a appelé d'une façon globale « sarcode » (la chair de la cellule) et l'a caractérisé comme une « substance claire, gluante et diaphane.» La description par Dujardin de la matrice cytoplasmique, publiée en 1835 (14), repose sur des examens microscopiques et des preuves expérimentales de corps cellulaires chez certains Foramenifères. Alors que les autres biologistes cellulaires de son temps avait fait référence simplement comme un gel vivant (ce qui est toujours aussi vrai aujourd'hui), Dujardin avait observé qu'il était insoluble dans l'eau, contracté en masses globuleuses, collant dans le sens où il adhérait aux aiguilles de dissection, tendu comme le mucus, et présent dans toutes les cellules examinées (les animaux inférieurs) (Fig. 5). |
|
Early Period In going back now to the beginnings (the early period), I shall be brief. It is difficult to identify the first and most significant observations on the optically empty matrix of the cell . It may have attracted the sharp eyes of van Leeuwenhoek as early as 1670. Meaningful observations, however, had to await the development of better microscopes, and, especially, of achromatic lenses . Many were attracted to the use of these new microscopes in the first half of the 19th century . The names of some are familiar: C . F. Wolff (1733-1794), B . de Mirbel (1776-1854), Lamarck (1744-1829), Dutrochet (1824), J . P. F . Turpin (1775-1840), Meyen (1804-1840), von Mahl (1805-1872), Brown (1773-1858) . All of these, with their observations on the cellular structure of plants and animals, preceded Schleiden and his Beitrage of 1838 by several years . Felix Dujardin (1801-1860) went further than most of the others with crude attempts to learn something about the physical nature of protoplasm . He called it inclusively "sar- code" (the flesh of the cell) and characterized it as a "clear glutinous diaphanous substance ." Dujardin's description of the cytoplasmic matrix, published in 1835 (14), was based on microscope examinations and experimental probings of the cell bodies of certain Foramenifera . Whereas other cell biol- ogists of his time had referred to it simply as a living gel (equally true today), Dujardin observed that it was insoluble in water, contracted into globular masses, was sticky in that it adhered to dissection needles, stretched like mucus, and occurred in all the cells (les animaux inférieurs) he examined (Fig . 5) . |
||||
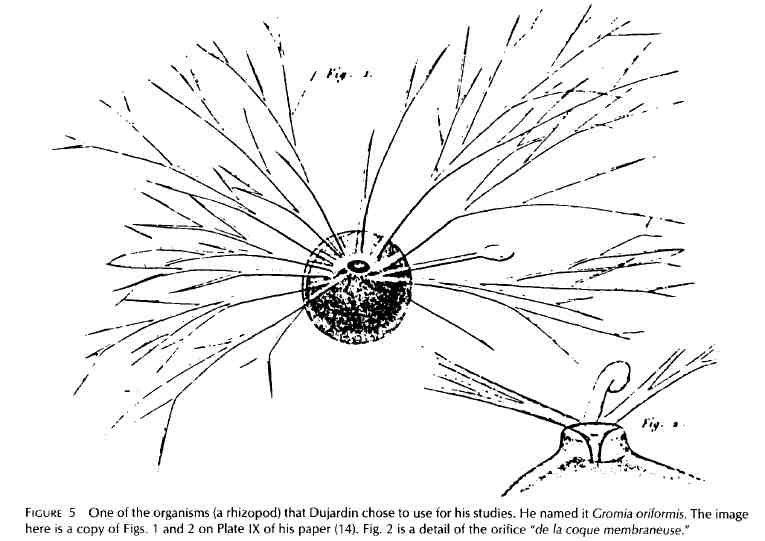 Un des organismes (Rhizopode) que Dujardin a utilisé pour ses études. Il l'a nommé Gromia oriformis. Cette image est une copie des figures 1 et 2 de la Planche IX de son article (14). La fig. 2 est un détail de l'orifice "de la coque membraneuse". |
||||||
|
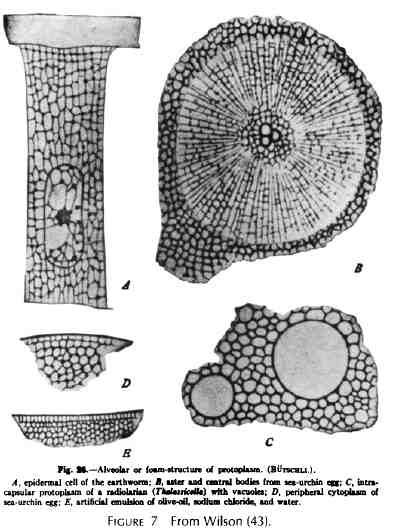
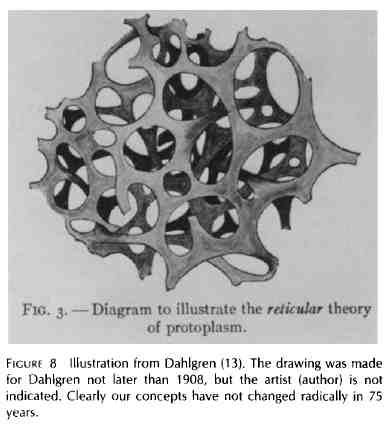
Dujardin était en avance sur son temps, mais pas totalement isolé dans ses observations sur la matrice. Je soupçonne que l'importance de ce qu'il a dit à ce sujet a été en grande partie minimisée par ses contemporains. Comme la théorie cellulaire a progressivement émergé des observations et de la pensée des contemporains de Dujardin, il était raisonnable que le « sarcode » fût, en plus de ses autres propriétés, très organisé. Ce qui, selon E. Brucke (1861) (4), était essentiel pour les diverses activités affichées par les cellules et afin de maintenir la cellule entière au-delà des processus purement physiques et chimiques. Cette affirmation a été répétée de nombreuses fois depuis 1861. Quarante ans plus tard Hofmeister suggéré dans un article de 1901sur l'organisation chimique de la cellule [cité dans Wilson (43)] que « d'un côté le morphologiste s'efforce d'élucider la structure du protoplasme à l'aide des plus fins détails; alors que de l'autre côté le biochimiste, avec ses méthodes de recherche apparemment grossières, bien que plus expérimentales, cherche à déterminer les fonctions chimiques du même protoplasme ; en gros, ils travaillent sur les deux faces opposées d'un même objet.» Beaucoup d'encre a coulé à ce sujet avec des mots et des dessins. Le morphologiste a vu la cellule comme possédant une substance indifférenciée, y compris le noyau, les mitochondries, et les centrioles, qui sont capables de croître et de se reproduire, ainsi qu'un protoplasme différencié, auto-entretenu et capable de réaliser des fonctions spéciales. Comme les techniques de coloration s'amélioraient en même temps que les microscopes, l'intérêt pour comprendre et définir la nature de la matrice visqueuse et limpide a perduré. Le hyaloplasme est devenu la substance visqueuse fondamentale ou matrice. Dans la plupart des cellules, ce hyaloplasme a été reconnu comme étant suffisamment visqueux pour empêcher le mouvement brownien. Structurellement certains biologistes pensaient qu'il était fondamentalement fibrillaire et les représentations élégantes (des dessins) ont été produites pour illustrer cette propriété (Heidenhain) (18) (Fig. 6). Pour d'autres (Bütschli) (6) il
était alvéolaire, comme une éponge. On
l'a appelée la théorie du protoplasme mousseux
(fig. 7). Pour d'autres, impressionnés par les
nombreuses mitochondries parfois observées, il
était essentiellement granulaire. Dahlgren (13) dans
son ouvrage sur l'histologie comparée a fourni un
dessin intéressant de la matrice cytoplasmique (il
s'agit de mon interprétation). La source de l'image
n'est pas donnée. S'il elle a été
obtenue à l'Université de Princeton, où
Dahlgren était professeur, c'était
probablement une synthèse des idées de
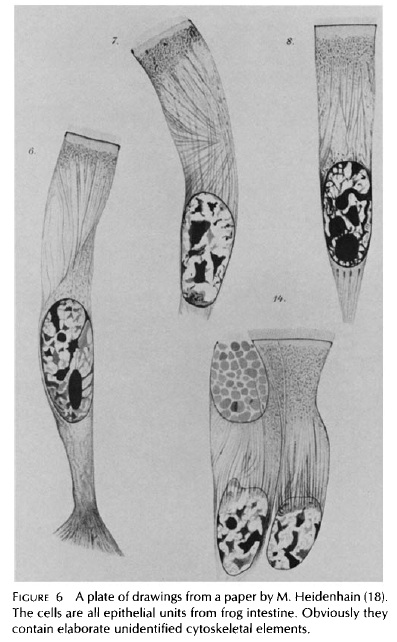
Dahlgren et Conklin (Fig. 8). Figure 6.Une planche de dessins à partir d'un article de M. Heidenhain (18). Les cellules sont toutes les sous-unités épithéliales de l'intestin de grenouille. Évidemment elles contiennent des éléments cytosqueletiques non identifiés. |
|
Dujardin was ahead of his time, though not totally alone in making observations on the matrix . I suspect that the importance of what he said about it was largely wasted on his contemporaries . As the Cell Theory gradually emerged from the observations and thinking of Dujardin's contemporaries, it was reasoned that the "sarcode" must, in addition to its other properties, be highly organized . This, according to E . Brucke (1861) (4), was essential for the various activities that cells display and set the intact cell above and apart from mere chemical and physical processes. This statement has been repeated many times since 1861 . Forty years later Hofmeister suggested in a 1901 paper on the chemical organization of the cell [cited in Wilson (43)] that "the morphologist on the one hand strives to elucidate the structure of protoplasm down to its finest details; the biochemist on the other hand, with his apparently cruder yet still more searching methods seeks to determine the chemical functions of the same protoplasm ; broadly speaking they are only dealing with two different sides of the same thing . " A lot of ink has been expended on this topic in words and drawings . The morphologist has viewed the cell as possessing an undifferentiated substance, including nucleus, mitochon- dria, and centrioles, which is able to grow and reproduce, and also a differentiated protoplasm, self-perpetuating and capable of performing special functions. As the techniques of staining improved along with microscopes, the interest in penetrating and defining the nature of the clear viscous matrix continued . The hyaloplasm became the viscid ground substance or ma- trix . In most cells, this hyaloplasm was observed to be suffi- ciently viscous to inhibit Brownian motion . Structurally it was thought by some biologists to be fundamentally fibrillar and elegant images (drawings) were produced to illustrate this property (Heidenhain) (18) (Fig. 6) . To others (Butschli) (6) it was alveolar, like a sponge. This was labeled the foam theory of protoplasm (Fig. 7) . For others, impressed by the sometimes numerous mitochondria, it was basically granular. Dahlgren (13) in his textbook on comparative histology, provided an interesting drawing of the cytoplasmic matrix (my interpretation) . The source of the image is not given . If created at Princeton University, where Dahlgren was a pro- fessor, it was probably a synthesis of Dahlgren's and Conklin's ideas (Fig . 8). |
||||
|
|
||||||
|
Ces différents entretiens ont attiré l'attention des rares qui ont choisi de ne rien croire de tout cela et, comme mentionné précédemment, qui se sont donné pour tâche de prouver que ces images étaient différents artefacts liés à la fixation ou la déshydratation ou les deux. Le côté positif est qu'ils ont probablement involontairement fait beaucoup en encourageant les études sur les cellules vivantes qui semblent dominer la biologie cellulaire de la période suivante. C'est à ce moment que M. Lewis et W. Lewis (25) ont regardé des cellules vivantes proliférer et se déplacer dans les conditions de culture in vitro. Ils étaient capables de discerner le comportement des mitochondries, les caractéristiques du mouvement cellulaire et quelques manifestations de la différenciation. Leurs observations ont été épargnées par les critiques de Fischer et Hardy. Mais il est vrai que les Lewis et leurs étudiants ne se sont pas beaucoup livrés à des spéculations sur la nature fondamentale de la matrice cytoplasmique. Ce sujet controversé avait semble-t-il déjà perdu à l'époque un peu de sa popularité antérieure. |
|
These various discussions attracted the attention of a few who chose to believe none of it and, as mentioned earlier, set about to prove that these various images were artifacts of fixation or dehydration or both . On the positive side, they probably did a lot inadvertently to encourage the studies of living cells that seemed to domi- nate the cell biology of the next period . It was at this time that M . Lewis and W . Lewis (25) were watching live cells proliferating and moving about under the conditions of in vitro culture . They were able to discern the behavior of mitochondria, the characteristics of cell motion, and some manifestations of differentiation . Their observations were not subject to the criticisms of Fischer and Hardy . On the other hand, the Lewises and their students did not indulge in much speculation on the fundamental nature of the cytoplasmic matrix . That controversial subject at that time had seemingly lost some of its earlier popularity .
|
||||
|
|
||||||
|
La période récente La période récente dans cette division arbitraire de l'histoire de l'étude de la cytomatrice a commencé juste avant et pendant la Seconde Guerre mondiale. Albert Claude (9), à l'Institut Rockefeller, avait isolé et étudié les propriétés de l'agent antitumoral I du poulet, qui sera appelé plus tard le virus du sarcome de Rous. À cette époque, il n'était pas considéré comme un virus. Pour obtenir un matériau témoin à partir d'une source normale, Claude avait broyé des foies d'embryons de poulet sains et soumis les homogénats à différentes centrifugations. À sa grande surprise, la fraction contenant l'agent de la tumeur et la fraction d'embryon de poulet, isolées de la même façon, ont fondamentalement les mêmes propriétés, sauf que la fraction obtenue à partir de tissus normaux ne peut pas produire de tumeurs. Ceci poussa Claude à commencer à une longue série d'études dans lesquelles ol a développé des techniques utiles pour l'isolement des noyaux, mitochondries et microsomes (fragments du réticulum endoplasmique). Il n'avait eu aucune formation académique en biologie cellulaire et confondait au début les mitochondries des homogénats de foie avec des granules de sécrétion (il y en a très peu dans le foie). Cette erreur a été corrigée par la suite par Palade et Hogeboom. Le fait essentiel est que Claude avait développé un bon procédé pour isoler et "purifier" les composants de la cellule. Ce qu'il a réalisé avec beaucoup de patience était une nette amélioration des procédés publiés six ans plus tôt par Andy Bensley Hoerr (3). La matrice cytoplasmique que nous abordons maintenant faisait partie du deuxième ou troisième surnageant, qui était généralement jeté. Elle a ensuite été nommée la fraction cytosolique. Malheureusement, le terme a été transféré du contenu du tube de centrifugation à la cellule vivante ainsi assimilée à un hyaloplasme ou une cytomatrice alors qu'il n'est pas un sol, du moins la plupart du temps. Ce qui s'est passé depuis, au sujet de la cytomatrice, est de l'histoire actuelle et sera mentionné par d'autres auteurs de ce supplément. Leur quête d'une compréhension de la matrice a été dominée par des approches biophysiques et biochimiques En dépit des progrès évidents rendus possibles par le fractionnement cellulaire tel qu'il a été pratiqué par Claude et ses disciples, il a persisté dans l'esprit de certains cytologistes l'idée que les fractions d'organites cytoplasmiques doivent être sévèrement endommagées par les procédures. Cette idée, entre autres, a inspiré Zalokar (47) pour tenter un fractionnement dans la cellule intacte. Les hyphes de Neurospora se sont avérés être un matériau favorable. Les cellules n'étaient pas détruites par la centrifugation à haut débit, et les différents composants sédimentaient en plusieurs couches facilement identifiées comme les mitochondries, les noyaux, le glycogène, et la composante basophile (ergastoplasme ou réticulum endoplasmique rugueux). Il y avait une cinquième ou une sixième couche qui était relativement libre des structures apparentes appelée surnageant et qui avait l'apparence d'un "treillis en nid d'abeilles". Des tests y ont révélé la présence de phosphatases. Les cellules viables après centrifugation, ont réorganisé leur contenu et continué à se propager. Des études similaires par Kempner et Miller (22) sur Euglena ont donné des résultats similaires. La cinquième couche, ou couche claire de leurs sédimentations s'est révélée être exempte de protéines et sans aucune structure, avec les moyens d'étude disponibles alors. À la suite de la centrifugation, dans les cellules qui avaient survécu, les divers organites sont retournés à leur distribution normale. [Voir aussi Clegg (10).] Une autre approche a été fournie par la microscopie électronique qui a commencé vers 1945 avec des observations sur des cellules entières en culture (32). Les premières micrographies obtenues étaient de peu de valeur, mais en peu de temps, comme nous avons appris quelques trucs pour la préparation, elles se sont améliorées et nous avons reconnu le réticulum plasmique et retrouvé ses caractéristiques ainsi que quelques petits systèmes fibreux (fibres de stress de Warren Lewis). Les techniques de manipulation étaient difficiles, et le séchage des cellules à l'air que l'on pratiquait était destructeur. Des études ultérieures ont assimilé la fraction microsomale avec le réticulum endoplasmique rugueux et avec la substance chromidiale (également appelée ergastoplasme) des cytologistes antérieurs. L'obtention de sections prêtes fines automatiques (autour de 1953) a ouvert un nouveau et vaste panorama pour l'exploration et de la découverte. Les choses les plus évidentes ont été d'abord étudiées notamment toutes les structures limitées par une membrane. Finalement, les microfilaments et les microtubules furent l'objet d'une certaine attention, en particulier à la suite de l'introduction du glutaraldéhyde comme un fixateur. La matrice cytoplasmique, cette partie sans aucun filament identifiable identifiable était moins intéressante à explorer. Elle apparaissait légèrement teintée, voire pas du tout, et essentiellement non structurée. |
|
Recent Period The recent period in this arbitrary division of the history of the study of the cytomatrix began just before and during World War II . Albert Claude (9), at The Rockefeller Institute, had been isolating and studying the properties of the chicken tumor I agent, later to be called the Rous sarcoma virus. In those days it was not considered a virus . To obtain control material from a normal source, Claude ground up liver and chick embryos and subjected the homogenates to differential centrifugation . To his surprise the fraction containing the tumor agent and the chick embryo fraction, similarly isolated, had essentially the same properties, except that the fraction from normal tissues did not produce tumors. This prompted Claude to initate an extended series of studies in which he developed useful techniques for the isolation of nuclei, mitochondria, and microsomes (fragments of the endoplasmic reticulum) . He had had no formal training in cell biology and intially confused mitochondria from liver homogenates with secretory granules (of which there are very few in liver). This error was subsequently corrected by Palade and Hogeboom . The essential fact is that Claude had developed a good pro- cedure for isolating and "purifying" cell components . What he achieved with much patience was a distinct improvement over the procedures published six years earlier by Bensley and Hoerr (3) . The cytoplasmic matrix we now address was part of the second or third supernatant, which was usually dis- carded . It was later named the cytosol fraction . Unfortunately, the term was transferred from the contents of the centrifuge tube to the living cell and equated with the hyaloplasm or cytomatrix which is not a sol, not most of the time, anyway . What has happened since, as far as the cytomatrix is con- cerned, is current history and will be mentioned by the other authors in this supplement . Their pursuit of an understanding of the matrix has been by biophysical and biochemical ap- proaches . In spite of the obvious achievement made possible by cell fractionation as practiced by Claude and his followers, there persisted in the minds of some cytologists the thought that the fractions of cytoplasmic organelles must be severely dam- aged by the procedures involved . This thought, among others, inspired Zalokar (47) to attempt a fractionation within the intact cell . The hyphae of Neurospora turned out to be a favorable material . The cells were not destroyed by high-speed centrifugation, and the various components stratified to pro- duce several layers easily identified as mitochondria, nucleus, glycogen, and the basophilic component (ergastoplasm or rough endoplasmic reticulum) . There was a fifth or sixth layer that was relatively free of formed structures called supernatant and that had the appearance of a "honeycomb lattice ." It tested positive for the presence of phosphatases . The cells, viable after centrifugation, reorganized their contents and continued to propagate . Similar studies by Kempner and Miller (22) on Euglena yielded similar results. The fifth, or clear layer of their strati- fications tested negative for protein and was structure free, as near as they could tell . Following centrifugation, which these cells survived, the various organelles returned to their normal distribution . [See also Clegg (10) .] Another approach has been provided by electron micros- copy, which began around 1945 with observations on whole cultured cells (32) . The first micrographs obtained were of limited value, but before long, as we learned some tricks of preparation, they got better and we recognized the endo- plasmic reticulum and its characteristics and a few small fiber systems (stress fibers of Warren Lewis) . The manipulative techniques were difficult, and drying the cells in air, as was then the practice, was destructive . Subsequent studies equated the microsome fraction with the rough endoplasmic reticulum and with the chromidial substance (also called ergastoplasm) of the earlier cytologists . The ready availability ofthin sections (around 1953) opened up a vast new world for exploration and discovery . The more obvious things were studied first and included all membrane-limited structures. Eventually, microfilaments and microtubules got some attention, espe- cially after glutaraldehyde was introduced as a fixative . The cytoplasmic matrix, that part exclusive of identifiable filaments, was least attractive for investigation . It appeared faintly stained, if at all, and essentially unstructured . |
||||
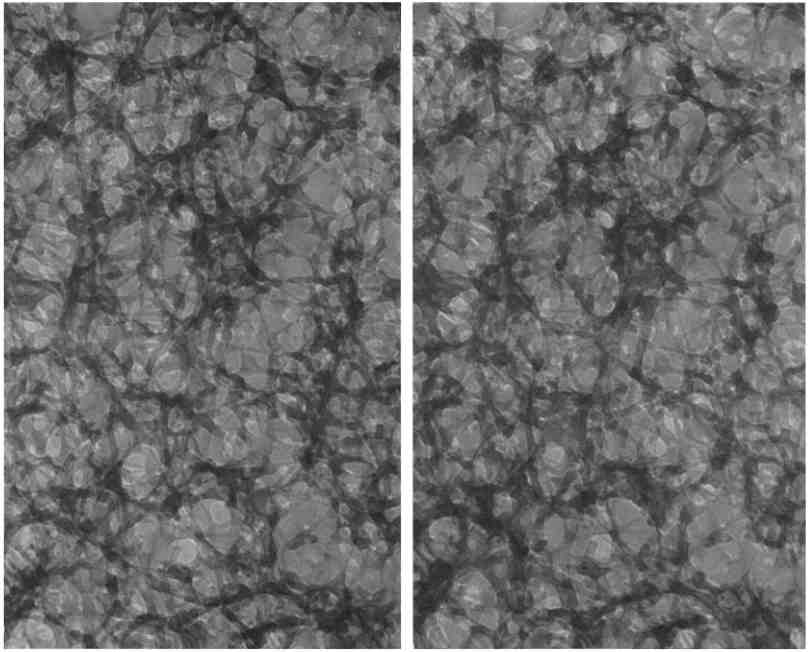 FIGURE 9 Micrographies électroniques stéréo (à haut voltage) décrivant la structure de la matrice cytoplasmique dans une bande étroite d'une culture de cellules NRK (rein de rat nouveau-né). La cellule a été fixée dans du glutaraldéhyde, figée dans le propane réfrigéré à -185°C et enfin séché pendant 1 jour 1/2 tout en étant maintenue à -95°C pour éviter la cristallisation. Dans cette expérience, nous avons évité le séchage au point critique et donc tous les artefacts qu'il peut induire. Dans le même temps, nous avons obtenu une image de la matrice fixée au glutaraldéhyde pour comparaison avec les cellules préservées uniquement par lyophilisation (voir Fig. 10). Il est évident que la morphologie résultant de la fixation au glutaraldéhyde est très similaire à celle obtenue après la seule lyophilisation. Un certain resserrement des trabécules est apparent dans ces images et dans l'analyse morphométrique de leurs dimensions (33). Les microtubules peuvent être identifiés dans ces figures et dans les suivantes comme des fibres de diamètre relativementdiamètre uniforme. x 80.000. |
||||||
|
"Wispy" (intrruduisaible) était un adjectif favori pour ce que l'on voyait là. Les microscopistes électroniques ajoutaient habituellement une coloration à l'uranyle et au plomb dans leurs protocoles, et, si une chose n'était pas colorée, elle ne méritait pas l'attention ou n'existait pas. Cependant, quelques desperados en recherche de sensations fortes sont entrés dans la mêlée. Surtout, ils se sont interrogés sur la structure fine de l'axoplasme et l'apparition de mécanismes possibles du transport axonal. Paul Burton et ses collègues (5) formèrent un groupe et Yamada, Spooner, et Wessels (46) un autre. J. Metuzals (28), tout seul à Ottawa, faisait le même type de travail. Ils ont tous publié des images montrant des réseaux ou des tissages de fils minces entre les neurofilaments et neurotubules. Je ne peux pas dire que beaucoup de leurs contemporains aient été convaincus, voire même intéressés. Nous trouvons convaincant l'argument selon lequel quelque chose dans la cellule doit rendre compte de la répartition non aléatoire des structures telles que le réticulum endoplasmique, l'appareil de Golgi, les microtubules, et les réseaux de microfilaments, en d'autres termes, de l'organisation (10). J'ai toujours été impressionné par le fait que les microfilaments (fibres de stress) apparaissent et disparaissaient au fur et à mesure que les cellules bougeaient et changeaient de forme. Pour moi, ainsi, il devrait y avoir une structure unitaire, un cytoplasme, construit autour du centre de la cellule et comprenant une population de corps denses (des centres organisateurs des microtubules) répartis de manière caractéristique pour chaque type de cellule. Le cytoplasme contient les composantes les plus visibles et contrôle leur assemblage ou désassemblage selon les besoins. Tout cela semble un peu tiré par les cheveux, mais nous avons été encouragés à conserver cette idée par des observations de cellules pigmentaires, qui nous disent que les granules pigmentaires individuels ont des positions fixes dans la matrice qui les déplace (34). À tout le moins, les arguments en faveur d'une matrice structurée (et il y en a plusieurs, de mentionnés dans cette histoire) nous ont encouragés à jeter un oeil, non pas à des cellules incluses dans la résine, mais à des cellules entières en culture séchées par la méthode du point critique ou autrement. Avoir à notre disposition un microscope à haut voltage a été d'une grande aide (fig. 9-11) (45). Nous avons trouvé et rapporté que nous pouvions affecter l'intégrité du réseau en l'exposant à de basses températures et que le réseau revient immédiatement à la forme normale lorsque la cellule est réincubée (36). Sa structure change en réponse à la cytochalasine et aux variations de la concentration de Ca2+ et de Mg2+ dans le milieu. Le retour à la normale lorsque les conditions normales sont restaurées est étonnamment rapide. C'est, autrement dit, un gel vivant. Les artefacts sont présents, bien sûr, mais ils ne semblent pas être importants. Le dessin de base et les dimensions sont très similaires que la cellule ait été fixée chimiquement ou conservée par congélation-séchage ou lyophilisation-substitution (33). Il n'est pas étonnant que le cil, en tant qu'extension du cytoplasme, possède son propre réseau composé de ce qu'on appelle des rayons. Ils ont une disposition ordonnée et personne ne remet en question leur existence dans le cil. Il y a plusieurs leçons à tirer de ces rayons. David Luck et ses collègues (26) ont montré que dans certains des mutants de Chlamydomonas les rayons n'existent pas ou apparaissent sous une forme raccourcie. Ils ont déterminé par leur double approche génétique et biochimique qu'environ 17 polypeptides différents sont impliqués dans la structure de rayon. Bien que la disposition ordonnée de ces polypeptides puisse facilement être vue dans des coupes fines, elle est très difficile à discerner dans des microphotographies du flagelle entier (Fig. 11). Cela ne devrait pas nous surprendre, si un peu d'ordre au sein de la matrice des trabécules se retrouvait, même non visible, dans les micrographies de cytoplasmes entier, épais. Conclusions Il me semble que la preuve d'un cytoplasme structuré (matrice) est incontestable. Il est structuré comme un gel qui intègre des filaments formés afin de donner une orientation aux mouvements intracellulaires, une anisométrie à la forme des cellules, et servir aux variations de viscosité. Il s'agit d'un gel vivant, à la fois dynamique et capable de répondre par des changements structurels à de nombreux stimuli, tout en préservant la capacité de revenir à une forme habituelle. L'auteur est heureux de souligner le soutien du Centre international Fogarty lors de la préparation de ce manuscrit et de l'organisation de la Conférence. Merci également à Marie Ulrich pour le travail sur les illustrations et à Joyce Albersheim pour les projets répétés de ce manuscrit. |
|
"Wispy" was a favorite adjective for what was there. Electron micros- copists habitually included staining with uranyl and lead in their procedures, and, if something failed to stain, it was not there or not worth attention . However, a few desperados in search of excitement entered the fray . Mostly they were inter- ested in the fine structure of axoplasm and the appearance of possible mechanisms for axoplasmic transport . Paul Burton and colleagues (5) formed one group and Yamada, Spooner, and Wessells (46) another. J . Metuzals (28), all alone in Ottawa, was doing the same type of work . They all published images showing networks or meshworks of slender strands between neurofilaments and neurotubules. I cannot say that very many of their contemporaries were convinced or even interested . We find compelling the argument that something in the cells has to account for the nonrandom distribution of formed structures such as the endoplasmic reticulum, the Golgi, the microtubules, and bundles of microfilaments, in other words, for the organization (10). I have always been impressed by the fact that microfilaments (stress fibers) appear and disappear as the cell moves and changes its shape . For me, there- fore, there should be a unit structure, a cytoplast, built around the cell center and including a population of dense bodies (microtubule-organizing centers) distributed in a manner characteristic for the cell type . The cytoplast contains the more visible components and controls their assembly or dis- assembly as required. All this sounds a little far-fetched, but we have been encouraged to stay with it by observations of pigment cells, which tell us that individual pigment granules have fixed positions in the matrix that moves them (34) . At the very least, the arguments for a structured matrix (and there are several, as mentioned in this history) have encouraged us to take a look, not at resin-embedded cells, but at whole cultured cells dried by the critical-point method or otherwise . Having at our disposal a high-voltage microscope has been a great help (Figs. 9-11) (45) . We have found and reported that we can affect the integrity of the lattice by exposing it to low temperatures and that the lattice immediately returns to normal form when the cell is reincubated (36) . Its structure changes in response to cytochalasin and to variations in the concentration of Ca" and Mg" in the medium . The return to normal when normal conditions are restored is amazingly rapid . It is, in a few words, a living gel . Artifacts are present, of course, but they seem not to be gross . The basic design and dimensions are very similar whether the cell is chemically fixed or preserved by freeze- drying or freeze-substitution (33). It is pertinent that the cilium, as an extension of the cytoplast, has its own lattice consisting of what are called spokes . They are ordered in their disposition and no one questions their existence in the cilium . There are several lessons to be learned from these spokes . David Luck and colleagues (26) have shown that in some mutants of Chlamydomonas the spokes either do not exist or appear in some abbreviated form . They have determined by their dual genetic and biochemical approach that about 17 different polypeptides are involved in the spoke structure . Though the orderly disposition of these polypeptides is readily seen in thin sections, it is discerned with difficulty in micro- graphs of the whole flagellum (Fig . 11). It should not surprise us, therefore, if some order among the matrix trabeculae is present, though not visible, in the micrographs of whole, thick cytoplasts. Conclusions It seems to me that the evidence for a structured cytoplast (matrix) is incontrovertible . It is structured like a gel that incorporates formed filaments for purposes of giving direction to intracellular motion, anisometry to cell form, and useful variations in viscosity . It is a living gel, at once dynamic and capable of responding by structural changes to numerous stimuli and yet preserving the capability of reverting to a preferred form .
The author is pleased to acknowledge the support of the Fogarty International Center during the preparation of this manuscript and the organization of the Conference . Thanks also go to Mary Ulrich for work on the illustrations and to Joyce Albersheim for repeated drafts of this manuscript . |
||||

FIGURE 10
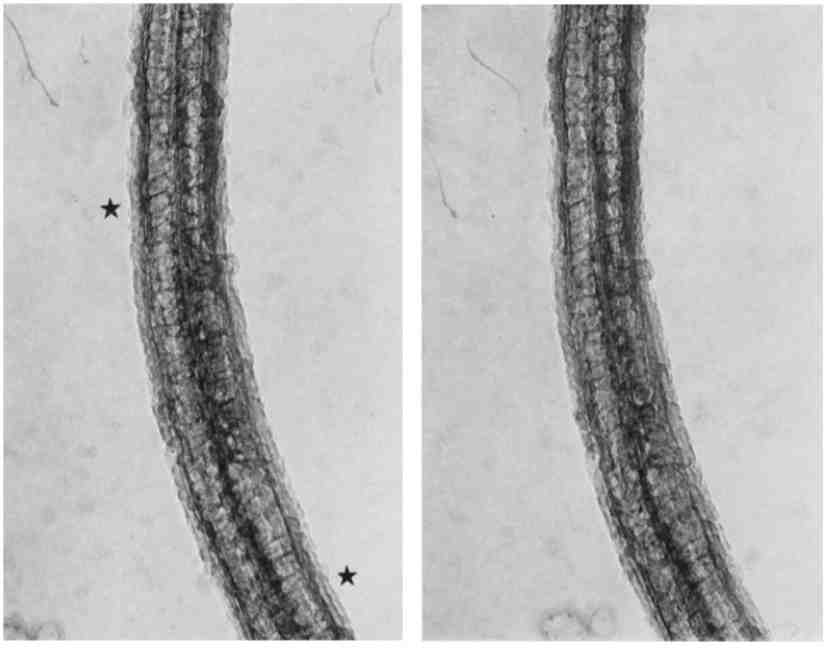 FIGURE 11 Paire d'images stéréo du flagelle d'un spermatozoïde d'oursin de mer fixé dans du glutaraldéhyde et séché par la méthode du point critique. Les rayons sont évidents (voir les étoiles), mais seulement en quelques endroits, et même pas clairement selon leur ordre en triplet repété, caractéristique des sections fines. Évidemment, la superposition de la structure contribue à la confusion dans les micrographies du flagelle entier. Cela, et le fait que l'orientation variable des rayons ne conduit qu'occasionnelement à les voir dans le plan image selon leur grand profil, rend compte du fait que la totalité de l'organisation des rayons de peut être vu. x 80.000. |
||||||
|
RÉFÉRENCES 1 . Beams, H. W. 1943 . Ultracentrifugal studies on cytoplasmic components and inclusions . In Biological Symposia: Frontiers in Cytochemistry, Vol . 10. J . Cattell, editor. Jaques Cattell Press, Lancaster, PA . 71-90 . 2 . Beams, H. W., and R. L. King. 1936 . Survival of Ascaris eggs after centrifuging. Science (Wash. DC) . 84:138 . 3 . Bensley. R. R., and N. J . Hoerr. 1934 . Studies on cell structure by the freeze-drying method . V. The chemical basis of the organization of the cell, Anal. Rec. 60:251-266. 4. Brucke, E . 1961 . Wiener Sitz . Ber. 44. 5 . Burton, P. R., and H. L . Fernandez. 1973 . Delineatio n by lanthanum staining of filamentous elements associated with the surfaces of axonal microtubles . J. Cell Sci. 12 :567-583 . 6. Butschli, O. 1898 . Untersuchungen uber Strukturen. Leipzig. 7. Chambers, R. 1940. The micromanipulation ofliving cells. In The Cell and Protoplasm. American Association for the Advancement of Science, No. 14. F . R. Moulton, editor. Science Press, Lancaster, PA . 20-30. 8 . Chambers, R., and H. B . Fell . 1931 . Micro-operations on cells in tissue culture . Proc. Ray. Soc, Land. 109:381 . 9. Claude, A. 1940 . Particulate components of normal and tumor cells. Science (Wash. DC) . 91 :77-78 . 10. Clegg, J . S. 1982. Interrelationships between water and cell metabolism in Artemia cysts. IX . Evidence for organization of soluble cytoplasmic enzymes. Cold Spring Harbor Symp . Quant. Biol. 46:23-37. 11. Conklin, E . G. 1940. Cell and protoplasm concepts : historical account. In The Cell and Protoplasm . American Association for the Advancement of Science, No . 14 . F. R. Moulton, editor . Science Press, Lancaster, PA . 6-19. 12. Crick, F. H. C., and A. F. W. Hughes. 1950. The physical properties of cytoplasm. A study by means of the magnetic particle method . I. Experimental. Exp. Cell Res. 1 :37- 80. 13 . Dahlgren, U., and W. A. Kepner . 1908 . Principles of Animal Histology. The Macmillan Company, New York, 14. Dujardin, F. 1835 . Recherches sur les organismes inferieurs . Ann. Sci . Nall. Zool. Biol. Anim. 4 :343-377. 15. Fischer, A. 1899. Fixierung, Fdrbung and Bau des Protoplasmas. Fischer Verlag, Jena. 16. Frey-Wyssling, A. 1948 . Submicroscopic Morphology of Protoplasm and Its Derivatives, 2nd ed . Elsevier Publishing Company, New York . 17. Hardy, W. B . 1899 . Structure of cell protoplasm . J. Physiol. (Land.) . 24:158 . 18. Heidenhain, M. 1899 . Leber die Struktur der Darmepithelzellen. Arch . Mikrosk. Anal. Entwicklungsg.54 :184-224 . 19. Heilbronn, A. 1922. A new method for the estimation of viscosity in living protoplasts. Jahrb . Wiss. Bot . 61 :284 . 20. Heilbrunn, L . V. 1956 . The Dynamics of Living Protoplasm . Academic Press, Inc . New York . 21 . Inoue, S., and H. Sato . 1967 . Cell motility by labile association of molecules. J. Gen. Physiol.50 :259-292 . 22. Kempner, E. S ., and J . H. Miller . 1968 . The molecular biology of Euglena gracilis. V . Enzyme localization. Exp. Cell Res . 51 :150-156 . 23 . Kitching, J . A., and D. C. Pease. 1939 . The liquefaction of the tentacles of suctorian protozoa at high hydrostatic pressures . J. Cell Comp. Physiol. 14 :1-3 . 24. Kopac, M. J . 1938 . The Devaux effect at oil-protoplasm interfaces. Biol. Bull . 75 :372 . 25 . Lewis, M. and D. Warren . 1915 . Mitochondria (and other cytoplasmic inclusions) in tissue cultures . Am. J. Anal . 17 :339 . 26 . Luck, D. J ., G. Piperno, Z. Ramanis, and B . Huang . 1977 . Flagellar mutants of Chlamydomonas: studies of radial spoke-defective strains by dikaryon and revertant analysis . Proc . Nall. Acad. Sci. USA . 74:3456-3460. |
|
27 . Marsland, D. A. 1942 . Protoplasmic streaming in relation to gel structure in the cytoplasm . In The Structure of Protoplasm: A Monograph of the American Society of Plant Physiologists. W. Seifriz, editor. Iowa State College Press, Ames, Iowa . 127-161 . 28 . Metuzals, J . 1969 . Configuration of a filamentous network in the axoplasm of the squid (Loligo pealii L.) giant nerve fiber. J Cell Biol. 43 :480-505 . 29 . Needham, 1 . 1937 . Chemical aspects of morphogenetic fields. In Perspectives in Biochemistry. Needham and Green, editors. Cambridge University Press. 66-80. 30. Pease, D. 1943 . The physical structure of nematode egg cytoplasm . Anat. Rec. 87 :36. 31 . Peters, R . A. 1937 . Proteins and cell organization . In Perspectives in Biochemistry. Needham and Green, editors . Cambridge University Press. 36-44 . 32. Porter, K. R., A. Claude, and E. F. Fullam . 1945 . A study of tissue culture cells by electron microscopy. J. Exp. Med. 81 :233-246 . 33 . Porter, K. R., and K. L. Anderson . 1982 . The structure of the cytoplasmic matrix preserved by freeze-drying and freeze-substitution. Eur. J Cell Biol. 29 :83-96 . 34 . Porter, K. R., and M. A. McNiven. 1982 . Th e cytoplast: a unit structure in chromatophores. Cell. 29:23-32 . 35 . Porter, K. R., M. Beckerle, and M. A. McNiven. 1983. Th e cytoplasmic matrix . In Spatial Organization of Eukaryotic Cells. Modern Cell Biology, Vol. 2. J. R. McIntosh, editor. Alan R. Liss, Inc. New York . 259-302 . 36 . Porter, K. R., and J. B . Tucker . 1981 . Th e ground substance of the living cell. Sci . Am. 244 :57-67 . 37 . Scarth, G. W. 1942. Structural differentiation of cytoplasm. In The Structure of Protoplasm : A Monograph of the American Society of Plant Physiologists. W. Seifriz, 1 2s THE JOURNAL OF CELL BIOLOGY - VOLUME 99, 1984 editor. Iowa State College Press, Ames, Iowa . 99-107. 38. Scarth, G. W. 1927. The structural organization of plant protoplasm in the light of micrurgy . Protoplasma. 2 :189-205 . 39 . Schmidt, W. J. 1937. Die Doppelbrechung von Karyoplasma, Zytoplasma and Metaplasma. Berlin . 40 . Seifriz, W. 1945 . The structure of protoplasm . Bor. Rev. 11 :231-259 . 41 . Strangeways, T. S . P., and R. G. Canti. 1927 . The living cell in vitro as shown by darkground illumination and the changes induced in such cells by fixing reagents . Q. J. Miocrosc . Sci. 71 :1-14. 42. Tilney, L. G., Y. Hiramoto, and D. Marsland . 1966. Studies on the microtubules of Heliozoa. Ill . A pressure analysis of the role of these structures in the formation and maintenance of the axopodia of Actinosphaerium nucleofrlum (Barret). J. Cell Biol. 29:77-95 . 43. Wilson, E . B. 1928 . The Cell in Development and Heredity, 3rd ed. The Macmillan Company, New York, 1232 . 44. Wilson, J . W. 1965 . The Graduate Journal, University of Texas. 17 :68-85 . 45 . Wolosewick, J. J ., and K. R. Porter. 1979. The microtrabeculae lattice of the cytoplasmic ground substance : artifact or reality . J Cell Biol. 82 :114-139 . 46. Yamada, K. M., B. S. Spooner, and N. K. Wessells. 1971 . Ultrastructure and function of growth cones and axons of cultured nerve cells . J. Cell Biol. 49:614-635. 47. Zalokar, M. 1960 . Cytochemistry of centrifuged hyphae of Neurospora. Exp. Cell Res. 19 :114-132. |
||||