La reproduction des êtres vivants
retour accueil, formulation
par cycle
1. Réflexions (se multiplier - seul
ou à deux - une flèche de vie -
panorama)
2. expériences et observations
1. Réflexions
a. se reproduire c'est se multiplier
Le travail de reproduction correspond
à la capacité des organismes vivants de se
multiplier.
La vie est extrêmement résistante, elle colonise tous
les milieux (l'homme, par son exploration spatiale, est en train de
coloniser le cosmos). On découvre de nos jours de nouvelles
formes bactériennes dans les inclusions fluides des roches
à plusieurs kilomètres de profondeur. On sait que la
vie peut résister à des températures de plus de
150°C dans des sources hydrothermales. La vie peut aussi par des
formes de résistance comme des graines rester plusieurs
centaines d'années en léthargie (graines d'herbiers du
XVème siècle).
On retrouve ici deux idées essentielles liées au
concept de reproduction : résister (à des
conditions défavorables) et coloniser (l'espace). La
troisième idée est celle de survivance au temps
: tout être vivant étant mortel, il survit en quelque
sorte à sa mort dans ses descendants.
Remarque:
Le terme de procréation, utilisé pour la
reproduction humaine est revenu dans les nouveaux programmes, il fait
nettement référence à la participation de la
créature à l'œuvre du Créateur (le terme
est formé en latin de la préposition pro
qui signifie pour, à la place de, en vue de; et de
creo, as, avi, atum, creare
: créer, donner l'être). Il reflète aussi le
fait que tout être vivant qui naît d'un autre est
fondamentalement différent de celui qui lui donne naissance,
ce que ne reflète pas le terme de transmission de la vie.
L'unicité des êtres vivants ne repose pas sur leur
unicité génétique et il n'y a pas lieu de faire
de différence entre deux bactéries nées par
scissiparité et une nouvelle plante issue d'un savant
croisement plus ou moins contrôlé. Par contre il est
probable que le terme de procréation fasse de plus en plus
référence chez de nombreuses personnes aux techniques
de procréation médicalement assistée, comme on
désigne de façon si inadéquate les techniques de
manipulation des cellules sexuelles et embryons humains.
b. on se reproduit seul ou à deux
On distingue classiquement deux grands groupes de
reproduction.
- D'abord la reproduction solitaire ou asexuée
existe chez les bactéries et les unicellulaires sous la
forme d'une division binaire (une cellule mère donne
deux cellules filles) mais aussi chez les pluricellulaires par la
libération de petits groupes de cellules (propagules par
exemple chez les végétaux ou bourgeons chez une
hydre) ou même de cellules isolées (spores par
exemple) qui reprennent le développement de l'organisme
"à zéro" (au point de départ, plus
précisément) pour donner naissance à un
nouvel individu complet. Chez les procaryotes on parle de
scissiparité alors que la division binaire
des cellules eucaryotes est parfois appelée à tort
mitose (la mitose ne désigne que les
phénomènes nucléaires, et il faut aussi tenir
compte des phénomènes cytoplasmiques: voir ci-dessous).
Chez les organismes pluricellulaires la division de nombreuses
cellules de l'organisme assure la formation de l'organisme adulte
au cours du développement et le maintien de celui-ci en vie
par renouvellement cellulaire (renouvellement des cellules mortes
chez les organismes à longue durée de vie).
Étant donné que l'hérédité ne
se réduit pas à l'hérédité
génétique (cytoplasme, organites) et que celle-ci
n'est pas non plus réduite à la
duplication-birépartition d'une séquence d'ADN
(plasmides, ADN mitochondrial et chloroplastique...), il est clair
que dire que la reproduction asexuée donne deux organismes
identiques est pour le moins une inexactitude. La reproduction
asexuée (en solitaire) donne deux individus
différents dont les caractères proviennent de la
cellule mère.
- Ensuite la reproduction en commun ou sexuée fait
intervenir une jonction des deux organismes avec des
échanges plus ou moins bien connus selon les
espèces: cette reproduction sexuée est connue chez
les bactéries (conjugaison, même si les
mécanismes l'apparentent plutôt à un
échange d'information appartenant au travail de relation),
les unicellulaires et les pluricellulaires. Chez les organismes
possédant des cellules à noyau (eucaryotes), la
reproduction sexuée est réalisée par une
gamétogenèse, formation des gamètes
dont l'étape essentielle est la méiose qui
est une série de deux divisions cellulaires suivie d'une
fécondation qui est une fusion cellulaire. Du point
de vue héréditaire et sans réduire la
transmission des caractères à une
hérédité mendélienne ou même
chromosomique, on peut cependant affirmer du point de vue
très général que la reproduction
sexuée donne des individus dont les caractères
proviennent à la fois de la mère et du
père.
c. la vie de chaque être vivant dessine
plutôt une flèche qu'un cycle
Dans le temps, les étapes du développement :
naissance, croissance, sénescence et mort se succèdent.
La vie se termine inéluctablement par la mort. En ce sens la
vie est une flèche pointée vers la mort.
Mais les biologistes ont pris l'habitude de présenter les
étapes du développement par des cycles ou "cycles de
vie" en faisant se rejoindre les étapes initiale et
reproductrice. Il y a certainement là des concepts
philosophiques liés notamment à l'observation des
cycles saisonniers.
La notion de cycle n'étant pas accessible avant le cycle 3 et
la notion de cycle de vie étant très discutable,
pourquoi ne pas abandonner tout simplement cette notion en primaire
?
Par contre les profonds changements saisonniers et la
cyclicité des périodes de reproduction peuvent
être abordés en primaire.
- Les données ne sont pas faciles à collecter chez
les Procaryotes et il semble bien que la reproduction va de
pair avec les conditions nutritives du milieu. Il existe aussi des
facteurs populationnels (une bactérie solitaire ne peut pas
être cultivée: soit on a une spore de
résistance, unique, soit on a une colonie : l'état
de cellule isolée est purement abstrait : l'état
social de la bactérie est bien la colonie). Sans avoir
d'autres renseignements, il est donc fréquent de dire que
les bactéries se reproduisent tant que des nutriments sont
disponibles et tant qu'il n'y a pas de signaux qui
empêcheraient la division. On considère ainsi qu'il y
a un état reproducteur permanent.
- Chez les unicellulaires les données sont
éparses et très diverses sans qu'il soit facile non
plus d'énoncer des généralités
simples. On considère souvent mais je ne sais pas si cette
opinion est vraiment fondée, que la reproduction
asexuée se fait dans des conditions favorables et la
reproduction sexuée lorsque les conditions de survie
(nutrition essentiellement) deviennent défavorables. Ceci
est justifié par le fait que l'on considère le
zygote (qui est souvent un kyste) comme une spore de
résistance. En fait il n'est pas certain que l'on puisse
généraliser et si certains unicellulaires ont des
kystes diploïdes, de nombreux kystes sont aussi
haploïdes et parfois même pluricellulaires (des
divisions cellulaires précédent l'enkystement).
Connaît-on réellement l'équipement
chromosomique de nombreux unicellulaires ? Étant
donné l'énorme variabilité ne serait-ce que
de la mitose chez ces organismes, je mets en doute notre
connaissance sur ce sujet.
- Chez les champignons, l'état reproducteur est
encore vu sous l'angle de la disponibilité des ressources
nutritives et des conditions physiques du milieu en l'absence de
données certaines (à ma connaissance) sur des
facteurs internes de maturation. On se limitera donc par exemple
à l'observation de l'apparition des spores chez les
moisissures (par exemple sur du pain mis à moisir) lorsque
les mycéliums sexués sont assez
développés et se rencontrent. De même
l'apparition des chapeaux (carpophores) des champignons de nos
régions est dépendante de conditions
d'humidité et de température qui ne sont
réunies qu'à l'automne pour la majorité des
espèces, en hiver pour certaines espèces et au
printemps pour d'autres. Les espèces développant des
appareils reproducteurs estivaux sont assez peu nombreuses.
- Chez les plantes, et du fait des applications
bio-technologiques (commerciales), la reproduction asexuée
est encore une fois considérée comme intervenant
surtout en période favorable (eau et nutriments en
quantité suffisante, absence de facteurs inhibiteurs),
alors que la reproduction sexuée est
considérée comme dépendant de facteurs
internes, héréditaires dont les supports ne sont pas
connus. On a pris l'habitude de classer les plantes en fonction de
types biologiques (Raunkier) qui regroupent les plantes selon les
adaptations qu'elles développent en période
défavorable (froid, sécheresse...). En fait cette
classification relève plutôt du travail de nutrition
que de reproduction pour lequel on distingue essentiellement les
plantes annuelles (qui se reproduisent par reproduction
sexuée au moins une fois par an et dont l'appareil
végétatif meurt chaque année; les plantes
bisannuelles qui fleurissent au bout deux ans (la
première année seul l'appareil
végétatif se développe), elles meurent
ensuite (on peut aussi considérer de rares espèces
trisannuelles qui ne se reproduiraient qu'au bout de trois ans) ;
et les plantes vivaces qui se reproduisent par reproduction
sexuée chaque année mais dont l'appareil
végétatif persiste d'une année sur l'autre et
ne meurt pas à la mauvaise saison : leur durée de
vie peut être très longue (certains arbres
pluricentenaires l'attestent).
|
classification des plantes
à fleurs (Angiospermes) en types
biologiques (par Raunkier)
|
|
thérophytes (du grec
"théros" = été) sont des
plantes annuelles qui passent la mauvaise saison sous
forme de graines ; elles sont rares sous les climats
froids et abondantes sous climat
méditerranéen (on parle
d'éphémérophytes dans les
régions arides).
|
|
plantes vivaces ou bisannuelles dont l'appareil
végétatif de dépasse pas 25 cm
(couche éventuelle de neige) et qui comprennent 3
types :
|
|
les géophytes (de
"gé"= terre) qui passent la mauvaise saison
sous forme de rhizomes, bulbes et tubercules
chargés de réserves ; elles sont abondantes
en climat tempéré
|
les hémicryptophytes (du grec
"hémi"= moitié et
"cryptos"=caché) qui ont des bourgeons au
niveau du sol; surtout des bisannuelles dans les
régions tempérées et des vivaces en
montagne, dans les régions
méditerranéennes et tropicales.
|
les chamæphytes (du grec
"chamaï"= rampant) sont de petits "arbustes"
(ligneux); ils sont surtout abondants en climat alpin et
très sec
|
|
plantes vivaces de plus de 25 cm de haut ou
phanérophytes (du grec
"phaneros"=visible): arbres, arbustes, grandes
plantes herbacée et épiphytes (qui vivent
sur d'autres plantes), surtout présents en milieu
tropical humide mais certains sont adaptés
à des climats tempérés en
réduisant les pertes d'eau par des
différentiations comme des écorces
épaisses (chêne vert) ou des feuilles
réduites (aiguilles des conifères) ou
encore la perte des feuilles en hiver (feuillus à
feuillage caduque).
|
- Chez les animaux aussi l'aptitude à se reproduire
sexuellement (puberté ou stade adulte) et le nombre
de phases de reproduction permet de distinguer trois grands
types de modes de vie :
|
reproduction sexuée -
générations et durée de vie
|
|
vie courte de durée inférieure
à un an
(possibilité de plusieurs
générations par an)
|
une ponte massive (par exemple les papillons)
|
|
plusieurs pontes successives (par exemple le
criquet)
|
|
vie de durée égale à un
an
(espèces annuelles)
|
une ponte massive (par exemple la seiche)
|
|
plusieurs pontes successives
|
|
vie de durée supérieure à
un an
(espèces pérennes)
|
une ou plusieurs pontes par an (par exemple
l'étoile de mer ou le lombric)
|
|
une seule phase de reproduction atteinte à une
âge de maturité sexuelle, supérieur
à deux ans (par exemple des annélides, mais
aussi l'homme)
|
d. une vue de la reproduction des êtres
vivants sous le double éclairage évolutif et
écologique : du milieu aquatique au milieu aérien et
retour
Chez tous les êtres vivants, la reproduction peut
s'envisager principalement sous le double éclairage
évolutif et écologique de sa dépendance
vis-à-vis du milieu et plus particulièrement de sa
dépendance vis-à-vis du milieu aquatique.
- Les Procaryotes sont aquatiques (libres ou symbiotes
(au sens large voir cours PE1: b - les
causes des maladies) d'organismes supérieurs comme
certains lichens ou des plantes de la famille des
légumineuses (Fabacées)) et se reproduisent
seuls par division binaire (scissiparité) et
uniquement dans un milieu aquatique. Ils ont par contre des formes
de résistance : les spores, qui leur permettent de
résister au milieu aérien et d'être
disséminés par voie aérienne. On observe des
accolement des bactéries (conjugaisons) avec
établissement de ponts cytoplasmiques et des
échanges d'ADN qui sont plutôt à mon avis
à interpréter comme du domaine du travail de
relation (communication) que de la reproduction (voir bibliographie
: Échanges de gènes entre
bactéries, Robert MILLER, 1998, La Recherche, 245, mars
1998, 60-65 ).
- Les Unicellulaires sont aussi aquatiques au sens large
de milieu libre (milieu marin ou milieu d'eau douce) et milieu
intérieur d'organismes supérieurs (symbiose au sens
large). Ils présentent la plupart du temps une reproduction
solitaire à laquelle s'ajoute, pour certains une
reproduction à deux, sexuée. Des formes de
résistance qualifiées de spores (souvent les
zygotes) sont connues pour certains.
La reproduction solitaire la division binaire
(phase M du cycle cellulaire) est d'une
part la division du noyau dont les
phénomènes chromosomiques sont bien connus car c'est
pendant cette unique étape de la vie de l'organisme que les
filaments d'ADN deviennent visibles en microscopie optique (c'est
la mitose (du grec "mitos" = fil, par
référence à la forme filamenteuse des
chromosomes, ou caryodiérèse), et d'autre part la
division du cytoplasme (ou cytodiérèse) qui
n'est pas obligatoirement égale. Ces deux mécanismes
qui peuvent être très séparés dans le
temps (des cellules à deux ou multiples noyaux (plasmodes)
sont fréquentes) sont sous la dépendance des
microtubules organisés parfois en organites (les
centrioles, que l'on retrouve notamment chez les animaux
(et pas à tous les stades du
développement: ils manquent souvent dans les
premières divisions de l'œuf voir cours
reproduction de l'homme remarques sur
le concept de méiose) mais qui manquent toujours
chez les plantes). (Remarque : on
connaît depuis les années 1960 une preuve
indiscutable d'une hérédité non
génétique chez les Paramécies (Biologie
moléculaire de la cellule, Alberts et al., 1995
(3ème éd.), Médecine-Sciences-Flammarion, p
820) : par une expérience de greffe on peut modifier
l'orientation du battement de quelques rangées de cils qui
entourent la totalité de la cellule : ces modifications se
transmettent à chaque division pour au moins 100
générations alors que l'information
génétique n'est pas modifiée : on pense que
l'information spatiale de l'orientation des cils se trouve dans
les centrioles). Mais de nombreux unicellulaires ont des
mitoses atypiques: pour certains, notamment des Diatomées
(unicellulaires chlorophylliens à petite coquille (test)
siliceux) l'enveloppe nucléaire persiste
(Biologie moléculaire de la cellule,
Alberts et al., 1995 (3ème éd.),
Médecine-Sciences-Flammarion, p 942).
La reproduction à deux (sexuée) fait
intervenir deux étapes : une
gamétogenèse : formation de gamètes ou
cellules sexuelles et une fécondation ou fusion des
cellules sexuelles.
- En ce qui concerne la gamétogenèse :
son mécanisme fondamental est la méiose
qui n'est pas l'homologue ni l'analogue de la mitose :
méiose vient du grec "méiosis" qui
signifie "décroissance" et qui fait
référence à une théorie de
l'hérédité chromosomique établie au
début des années 30 : la méiose est une
suite de deux divisions qui aboutissent à la
réduction du nombre de chromosomes: on passe d'une
cellule diploïde (2n chromosomes) à une -ou quatre
théoriquement - cellule(s) haploïde(s) (à n
chromosomes). Ce concept n'est pas adapté au primaire et
je me suis efforcé de montrer qu'il n'était qu'un
concept et que donc on pouvait s'en passer dans le cours
sur la reproduction de l'homme).
La gamétogenèse, notamment chez les champignons,
n'est pas associée à une libération de
cellules spécialisées et peut être
plutôt qualifiée de maturation
sexuelle.
- Ce sont les caractères de la
fécondation qui sont intéressants à
observer: selon la taille respective des gamètes (en cas
de doute le plus gros est qualifié de femelle mais de
nombreux gamètes sont absolument identiques) et selon
leur moyen de déplacement (des flagelles le plus souvent
mais, de nouveau, le gamète immobile est, en cas de
doute, qualifié de femelle, et le gamète mobile
de mâle), ou encore le mode de fusion (accolement, fusion
des membranes, réalisation d'un siphon....). La
diversité n'empêche pas l'omniprésence du
milieu aquatique dans lequel a lieu cette
fécondation.
- Les Champignons ont sans aucun doute conquis le milieu
aérien. Leur reproduction est parfois une fragmentation de
leur mycélium (filament pluricellulaire qui naît par
divisions successives de chaque cellule par des mécanismes
de divisions voisins de ceux des unicellulaires).
Mais le plus souvent leur reproduction solitaire est basée
sur la production de spores qui sont aussi des formes de
résistance qui sont disséminées dans les
milieux humides terrestres.
Leur reproduction à deux (sexuée) est assez complexe
car, si la plupart des mycéliums sont haploïdes,
certains sont cœnocytiques (ont de nombreux noyaux). La
fécondation de deux mycéliums haploïdes donne
des spores diploïdes (zygospores) ou se réalise en
deux temps. D'abord la formation d'un mycélium très
particulier composé de cellules à deux noyaux (stade
dicaryote) qui forme, par exemple chez nos champignons "à
chapeau", l'appareil reproducteur visible : le carpophore (ou
chapeau que l'on cueille et que l'on mange parfois à nos
risques et périls)). Ensuite la fusion des deux noyaux
(caryogamie) haploïdes. Si la méiose intervient
immédiatement après la caryogamie il y a formation
des spores haploïdes (ascospores chez les champignons
à asques comme la truffe noire ou basidiospores comme chez
le champignon de Paris). Les spores germent et donnent à
leur tour un mycélium. On ne peut donc pas vraiment parler
de gamétogenèse chez les champignons car il n'y a
pas de phase de libération de gamètes qui iraient se
féconder. La maturation complexe des mycéliums et
leur fécondation, réalisée entre deux
cellules appartenant à deux mycéliums qui s'accolent
et sont parfois reliés par des ponts cytoplasmiques ou
siphons, est vraiment spécifique des champignons et tout
à fait adaptée à un milieu humide mais non
aquatique.
- Contrairement à une idée courante,
l'innovation majeure des Plantes n'est pas la
photosynthèse qu'elles partagent avec de nombreuses
bactéries et unicellulaires chlorophylliens, mais leur
progressive (dans le sens d'inégalement
achevée)
indépendance vis-à-vis
du milieu aquatique, surtout pour leur
reproduction. Ce que l'on peut aussi qualifier d'adaptation
au milieu aérien ou encore de conquête du
milieu aérien, si l'on se réfère aux
étapes de l'évolution telle que l'on la comprend
actuellement (l'adaptation au milieu aérien n'est bien
sûr pas uniquement basée sur les caractères de
la reproduction: on peut citer aussi la présence de lignine
dans la paroi qui permet le port dressé et la
réalisation de tissus conducteurs...).
La reproduction asexuée
des plantes se fait par des mécanismes
particulièrement efficaces faisant appel parfois à
des organes spécialisés. Elle fait l'objet d'un
chapitre spécial.
La reproduction sexuée des plantes peut se
considérer aux niveau de chacun des mécanismes
fondamentaux: gamétogenèse et fécondation:
- en ce qui concerne la gamétogenèse, il
n'y a pas chez les plantes une lignée germinale dont les
cellules pourraient être suivies depuis l'embryon
jusqu'à la plante à l'état reproducteur.
Des cellules apicales (situées aux sommets ou apex) qui
participent à la croissance de la plante changent de
potentialité en donnent des organes reproducteurs de
structure variées mais contenant une ou des cellules qui
subissent une maturation sexuelle avec notamment une
réduction du nombre de chromosomes (méiose). Les
cellules haploïdes ainsi formées sont des
spores. L'individu qui forme ces spores est
appelé le sporophyte. Le sporophyte est donc issu
du développement du zygote. Les spores
haploïdes donnent un individu haploïde par divisions
cellulaires (mitoses et divisions cytoplasmiques). C'est au
sein de cet individu que se passe la formation des
gamètes d'où son nom de
gamétophyte. La gamétogènèse
se passe donc en deux temps : formation des spores chez le
sporophyte et formation des gamètes chez le
gamétophyte.
La diversité de la gamétogenèse chez les
végétaux peut être considérée
en regard de l'importance de chacune de ces
générations (sporophytique et
gamétophytique), l'une par rapport à l'autre et
selon chaque sexe. Globalement on peut dire que le
gamétophyte est très réduit chez les
algues brunes comme une laminaire (c'est un petit
filament de quelques files de cellules qui produit des
gamètes aquatiques) et même presque inexistant
chez le Fucus (où la spore n'est pas
libérée mais se différencie directement en
gamète dans les cryptes reproductrices de l'algue). Le
gamétophyte est par contre dominant chez les
Bryophytes (mousses et hépatiques) et le
sporophyte se développe alors sur le gamétophyte
femelle ("en parasite"). Chez les Ptéridophytes
(fougères et prêles) les individus les plus
développés sont des sporophytes, le
gamétophyte étant réduit à une
petite lame de cellules (prothalle). Enfin, chez les plantes
à fleurs, le gamétophyte est réduit, au
grain de pollen pour le gamétophyte mâle qui est
libéré, et au sac embryonnaire pour le
gamétophyte femelle qui n'est pas libéré
par le sporophyte (le gamétophyte est contenu dans
l'ovule).
Il est difficile de voir un lien entre le milieu et
l'importance relative de chacune de ces
générations mais il est cependant possible de
distinguer des évolutions (régression du
gamétophyte, non libération du gamétophyte
femelle) en comparant ces structures, notamment en fonction de
leur apparition et par rapport à des groupes
fossiles.
- en ce qui concerne la fécondation, la
reproduction sexuée des plantes peut être
présentée comme un
affranchissement progressif du milieu
aérien (reprise dans les noms utilisés
pour la classification des plantes donc je brosse ici les
grandes lignes, non pour vous donner des éléments
à mémoriser mais pour justifier mes propos sur
les concepts simplifiés qui seront donnés en
primaire):
- chez les algues brunes
(Thallophytes: plantes à
thalle : organe végétatif non organisé
en axe feuillé) elle met en jeu la
libération de gamètes
aquatiques (spermatozoïdes flagellés)
suivie d'une fécondation
aquatique (on peut d'ailleurs facilement voir cette
fécondation à partir des organes reproducteurs
du Fucus sous le TRIDACT par
exemple).
- chez les embryophytes (division reprise par de
nombreux botanistes modernes et qui regroupe les plantes
à embryon) on observe une phase où un nouvel
individu se développe aux dépens d'un individu
dont il est issu (d'où le terme d'embryon). Les
embryophytes comprennent les Bryophytes (mousses et
hépatiques) et les Trachéophytes (plantes
à vaisseaux conducteurs) qui regroupent d'une part
les Ptéridophytes (ou Cryptogames vasculaires, qui
sont les fougères et prêles) et d'autre part
les plantes à graines: les Spermaphytes (du
grec "sperma"= graine) qui sont aussi les plantes
à fleurs (au sens large).
Chez les Bryophytes la
fécondation est
aquatique à la faveur d'un film d'eau
nécessaire à la survie des mousses mais
seuls les spermatozoïdes
flagellés se déplacent, l'
oosphère (gamète femelle) reste au fond
d'une structure en forme de bouteille (l'
archégone) sans être
libérée).
Chez les Ptéridophytes les gamètes
mâles sont aussi flagellés et libres et
la fécondation est
aquatique (mais cachée dans une crypte
d'où le nom de "cryptogame") mais les gamètes
femelles (oosphères) sont toujours immobiles au fond
d'un canal rempli de mucilage.
Les Spermaphytes sont scindés habituellement
en Gymnospermes (graines nues du grec "gymno"=
nu) qui ne possèdent pas de carpelles et qui donnent
des ovules nus (gymnovulie serait un terme plus
adapté que gymnospermie) et Angiospermes (du
grec "aggeion" = "petite urne") dont les graines sont
contenues dans des fruits qui résultent du
développement des carpelles qui entourent les ovules
(angio-ovulie serait encore un terme plus adapté).
Dans les deux cas la germination du grain de pollen conduit
à la libération, au voisinage de
l'oosphère, du gamète mâle, presque
réduit à son noyau, et à une
fécondation non
aquatique qualifiée de siphonogamie. Seule
la reproduction sexuée des Angiospermes sera
étudiée un peu en détail pour aborder
les concepts de fleurs, de graine et de fruit
notamment.
- il existe des cas de développement
de tissus ayant déjà commencé leur
maturation sexuelle et qui redonnent un individu complet sans
fécondation. L'aposporie correspond au
développement d'un cellule issue du sporophyte (une
spore restée diploïde par exemple), ce
phénomène est notamment connu chez des
fougères. L'apogamie correspond au
développement d'une cellule diploïde issue du
gamétophyte (le plus souvent du sac embryonnaire): A ne
pas confondre avec les cas de parthénocarpie (du
grec "parthenos"="vierge" et "carpos"="fruit")
où le fruit ne contient pas de graine (il se
développe sans fécondation et donc sans graine,
ce qui est recherché par l'homme et obtenu pour les
mandarines, la banane... mais ce ne sont pas des
phénomènes naturels fréquents mais bien
des anomalies extrêmement rares). De même
l'homme s'est efforcé de réaliser des cultures in
vitro de cellules de pollen ou autres cellules sexuées
qui relèvent de la multiplication
végétative (in vitro) et non de la reproduction
sexuée.
- Chez les animaux on retrouve dans de nombreux
embranchements la même caractéristique que pour les
plantes concernant le rapport avec le milieu.
La reproduction en solitaire (asexuée) est assez
fréquente chez les invertébrés : par
scissiparité (Vers plats, divers Annélides)
ou par bourgeonnement (Spongiaires, Cnidaires, certains
Annélides, Bryozoaires) principalement. Elle est donc
absente notamment chez les Mollusques, les Arthropodes et les
Vertébrés. On peut éventuellement lui
associer les phénomènes de
régénération qui sont une
capacité de régénération
complète d'un individu à partir d'un fragment chez
une hydre mais qui se réduisent à une
régénération partielle chez une
Annélide ou une étoile de mer (Echinodermes
Stellérides: un bras par exemple), ou encore très
localisée comme chez les Crustacés (une pince par
exemple).
La reproduction sexuée qui fait intervenir la
gamétogenèse et la fécondation.
Les cas de développement à partir de cellules
sexuelles non fécondées ne sont pas rares: c'est la
parthénogenèse (développement d'un
œuf (au sens de gamète femelle) vierge (du grec
"parthénos" = "vierge") sans
fécondation, qui est une apogamie pour les
végétaux). On connaît aussi des cas de
polyembryonie qui est une reproduction asexuée qui
affecte l'individu au stade embryonnaire: dans l'œuf
fécondé se développent plusieurs embryons en
parasite d'un embryon qui dégénère (cas par
exemple de quelques insectes Braconidæ).
On ne confondra pas avec les techniques artificielles
d'androgenèse (développement du gamète
mâle: développement d'un zygote dont on
détruit le noyau pronucléus femelle) et la
gynogenèse (de la même manière: la
fécondation à lieu du point de vue cytologique avec
activation de l'ovocyte mais sans pénétration du
noyau du gamète mâle) qui par contre est connue dans
la nature chez certains poissons, amphibiens et reptiles
(expérimentalement on la réalise chez des poissons
d'élevage comme la truite ou la carpe). Récemment
des techniques de clonage et de transplantation
nucléaire ont été
développées notamment chez les
Mammifères.
Depuis A. Weissmann (1892) il est d'usage de considérer
qu'il existe très précocement chez l'embryon des
cellules dont la destinée sexuelle exclusive les fait
regrouper au sein d'un ensemble qualifié de lignée
germinale (germen) par opposition aux autres
cellules formant le soma (lignée
somatique). Ces cellules colonisent des tissus et
forment avec eux les gonades ou glandes sexuelles : les
ovaires chez la femelle et les testicules chez le
mâle. Chez les espèces hermaphrodites (on dit
aussi monoïques comme pour les végétaux car ils
possèdent deux types de gonades) les individus sont
bisexués (mais il peut y avoir une séparation des
sexes dans le temps: un individu n'étant pas en même
temps mâle et femelle). Chez les espèces
gonochoriques (ou dioïques) les sexes sont
séparés. La gamétogenèse a
toujours lieu dans les gonades (ovogenèse, qui produit des
ovocytes et spermatogenèse qui produit des
spermatozoïdes) mais elle est de durée très
variable. Elle peut commencer très précocement
(avant la naissance chez le fœtus par exemple) mais la
capacité à se reproduire et donc à
réaliser la fécondation grâce à
la production de gamètes est acquise lors de la
puberté. Il n'y a jamais, comme chez les plantes, une
génération haploïde (gamétophytique).
L'individu sexué (diploïde) produit toujours des
gamètes. Par contre les modalités de la
fécondation sont variées et les gamètes ou le
zygote ne sont pas toujours libérés. Le zygote
contient -ou est entouré par- des réserves
nutritives (oviparité) ou se développe
aux dépens de l'organisme femelle
(viviparité). Les états intermédiaires
sont aussi possible (ovoviviparité) avec maintien de
l'œuf qui se développe en embryon à
l'intérieur d'une enveloppe protectrice dans l'organisme
maternel mais qui est tout de même pondu et achève
son développement à l'extérieur
(l'éclosion à lieu en dehors de la mère) :
c'est le cas par exemple chez certains insectes (la mouche
tsé-tsé) ou poissons.
Remarque:
Il est clair que cette distinction commode ne rend
pas réellement compte du développement de l'embryon
à l'intérieur de la structure œuf. En effet
l'œuf de poule contient une embryon (au stade blastula) qui a
déjà 20 heures de vie intra-utérine. Le
développement s'arrête lors de la ponte (pendant une
dizaine de jours au maximum pour être provisoire) et ne
reprend que s'il y a incubation.
Mais une grande caractéristique des animaux est que la
fécondation n'est pas le terme de la reproduction
sexuée des animaux. Le zygote n'est pas
libéré dans le milieu de façon
générale, même pourvu de réserves et de
protections (œuf au sens le plus large de structure pondue
par la femelle). Très souvent le développement des
animaux est assez original et on observe des stades de
développement dont le milieu de vie et l'organisation sont
franchement différents de l'adulte (définit comme
l'individu pubère): ces individus sont appelés
larves par opposition aux jeunes qui ne
diffèrent de l'adulte ni par le mode de vie, ni par le
milieu de vie. Le terme de métamorphose
désigne alors les transformations comportementales
(éthologiques), anatomiques, histologiques et
physiologiques que subissent les larves lorsqu'elles passent d'un
stade larvaire à un mode-milieu de vie particulier
(qualifié de larvaire) à un autre (identique
à celui de l'adulte). Ce type de développement avec
métamorphose est qualifié de développement
indirect par opposition au développement direct
qui fait passer de l'embryon au jeune et à l'adulte sans
changement de milieu-mode de vie.
On peut mettre en relation les caractéristiques
cytologiques de l'œuf (au sens de cellule sexuelle femelle
mûre) avec le développement. Sa taille (en
fait son volume) tout d'abord et ses réserves
qui sont liées à ce paramètre. Un cellule
embryonnaire typique humaine a un diamètre d'une dizaine de
micromètres alors que la cellule œuf en mesure
typiquement 120 (le rapport entre le volume de ces deux cellules
supposées sphériques est de 1/1000 car je n'ose
rappeler que le volume augmente avec le cube du rayon de la
sphère). On a un rapport assez équivalent pour
l'œuf d'oursin (Echinoderme) ou de Cnidaire. Une cellule
embryonnaire d'amphibien ne semble pas être fondamentalement
de taille différente à celle d'un embryon humain et
donc voisine d'une à quelques dizaines de
micromètres alors que la cellule œuf d'amphibien
atteint de un à quelques millimètres (ce qui fait un
rapport de volume de l'ordre de 1/8000). On a un rapport voisin
pour les Annélides, les Mollusques Lamellibranches, les
Agnathes.... Les œufs de très grande taille (oiseaux,
reptiles, Céphalopodes, Sélaciens,
Monotrèmes...) atteignent quelques centimètres pour
une cellule embryonnaire de quelques centaines de
micromètres tout au plus, ce qui fait un rapport volumique
de plus de 1/ 1.000.000 (quoique l'on s'éloigne de plus en
plus du modèle sphérique). Pour les arthropodes
(dont les insectes), l'œuf, franchement allongé et
atteignant quelques millimètres, semble présenter un
rapport voisin de celui des amphibiens. Cet accroissement
volumique peut être relié directement à la
quantité de réserves. Les catégories
cités plus haut correspondent aux classiques œufs
alécithes, oligolécithes,
hétérolécithes, télolécithes et
centrolécithes (présentés ci-dessous) qui
font aussi référence à la position du
noyau de l'œuf dans la cellule géante. Ces
catégories, très perfectibles, sont essentielles
pour comprendre les premières étapes du
développement embryonnaire. On citera cependant les cocons
des Plathelminthes, par exemple, qui contiennent une œuf
entouré de cellules vitellines complètes... et qui
ne rentrent pas dans ce cadre trop étroit
Si l'on cherche à dégager les grandes tendances
évolutives de la reproduction des animaux on
considère :
- d'une part le nombre de gamètes, leur
libération et leur mobilité qui va
de pair avec les caractères de la fécondation
(externe-interne, aquatique-non aquatique); on peut dire
qu'il y a toujours plus de gamètes mâles que de
gamètes femelles (des chiffres de 20
millions de spermatozoïdes pour l'homme par jour et 7
millions d'ovocytes pour la femme dont seulement quelques
centaines seront ovulés entre la puberté et la
ménopause) et qu'il y a d'autant moins de
gamètes femelles que ses réserves sont
importantes (2-3 millions d'œufs pour
l'esturgeon par an et par kilo de poids de la femelle, 2-4.000
œufs pour la grenouille par an pour 300 au maximum pour
une poule pondeuse sélectionnée ).
Remarque:
Les stratégies
démographiques de Cole (1954) (r et K) sont parfois
utilisées abusivement. Ces sont des modèles
mathématiques des évolutions populationnelles
extrapolés même aux populations fossiles. Il est
hors de question de les employer en dehors de leur contexte
évolutif et sans les précautions scientifiques
habituelles. Je renvoie donc à des ouvrages
d'écologie spécialisés (par exemple:
Précis d'écologie, Roger Dajoz, Dunod, 196, p
233-246).
- et d'autre part la quantité des réserves
que les zygotes contiennent (lié aux
caractéristiques de l'œuf) dépendant du
devenir du zygote (libération - oviparité;
maintien dans les voies génitales maternelles -
ovoviviparité- viviparité), et bien sûr le
développement de l'embryon, direct ou indirect,
avec enfin le soin aux jeunes.
|
une classification basée sur les types
d'œufs en rapport avec leur devenir
|
|
type d'œufs
|
taille et rapport entre une cellule
embryonnaire et cette cellule
géante
|
caractères
|
exemples
|
devenir de l'œuf
|
|
œufs holoblastiques
=
segmentation totale
|
alécithes
|
0,1 mm
1/1.000 à 1/10.000
|
perte secondaire du vitellus ?
|
Mammifères placentaires (homme...)
|
œufs peu nombreux
fécondation interne
viviparité
|
|
oligolécithes
|
vitellus peu abondant et réparti
uniformément, noyau légèrement
excentré
|
éponges, oursins, étoiles de mer
|
œufs nombreux
fécondation externe et aquatique
(généralement)
oviparité
|
|
hétérolécithes
|
1 mm
1/10.000
|
vitellus abondant réparti surtout à
un pôle (végétatif), noyau
excentré (pôle animal)
|
Annélides, Mollusques Gastéropodes,
certains poissons, Amphibiens
|
|
oeufs méroblastiques
=
segmentation partielle
|
centrolécithes
|
1 à plusieurs mm
1/10.000
|
vitellus abondant, noyau central, cytoplasme
rejeté à la périphérie
|
insectes, crustacés
|
œufs nombreux
fécondation interne
oviparité
|
|
télolécithes
|
1 à plusieurs cm
1/1.000.000
|
œufs volumineux à vitellus très
abondant, protoplasme refoulé à la
surface de l'œuf
|
Mollusques Céphalopodes, Sélaciens et
Poissons téléostéens, Oiseaux,
Reptiles, Mammifères ovipares
(Ornithorhynque)
|
œufs moins à peu nombreux
fécondation externe ou interne
oviparité ou
ovoviviparité
|
2. Expériences et
observations
Une référence: Travaux Pratiques de Biologie,
Didier Pol, Bordas, 1994, p 56-59 - quelques mois favorables
au recueil de gamètes permettant une fécondation
expérimentale in vitro en classe: on voit que le
Finistère est privilégié pour ces
observations...
|
01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
|
oursin
|
|
|
|
|
|
|
|
|
oursin
|
|
|
|
moule
|
|
|
|
|
|
|
|
|
|
|
annelides tubicoles
|
|
|
|
|
|
|
|
|
|
|
|
|
|
huître
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Fucus
|
|
|
|
Ascophyllum
|
|
|
|
|
|
|
|
|
|
Fécondation chez des
Annelides Polychètes tubicoles de nos rivages bretons
(voir aussi TP de la classe de
seconde sur le site lycée associé)
On peut trouver assez facilement aux grandes marées
(coefficient supérieur environ à 90) dans la zone
infralittorale (qui n'est découverte qu'aux marées
de vives-eaux) et en dessous, des colonies d'Annelides
(vers annelés) tubicoles (vivant dans des tubes)
Polychètes (soies nombreuses sur le corps). Ces
colonies forment des amas sur les rochers au voisinage des plages de
sable fin. (Par exemple, je n'ai eu aucun mal à trouver une
colonie sur la plage la plus proche de chez moi: plage de Kersidan,
commune de Trégunc, entre Pont-Aven et Concarneau, sur le
côté abrité du cordon rocheux latéral mais
au contact de la zone sableuse où, bien sûr, l'animal
trouve du sable pour camoufler son tube).
Si le prélèvement est LIMITÉ à
quelques individus pour un TP (chaque tube bien formé est
habité, la répartition mâle-femelle étant
homogène, il suffit de prendre un tube par
élève), il ne me paraît pas qu'il y ait de danger
pour l'espèce mais il ne faut pas se leurrer : on
détruit une partie des colonies en prélevant ces vers
pour les TP. Les tubes sont prélevés avec la lame d'un
couteau car le sable qui enrobe chaque tube organique interne
(marron), où se trouve l'animal, est bien induré. Les
tubes habités peuvent se conserver une bonne semaine à
4-5°C ou au bas du réfrigérateur en milieu humide
mais non sous l'eau. Penser à prélever aussi de l'eau
de mer pour les observations. J'ai conservé aussi les tubes
dans l'eau de mer sans dommage pendant 3 jours (on peut certainement
les conserver plus longtemps dans un aquarium d'eau de mer
oxygénée).
On trouve des hermelles: Sabellaria alveolata et
d'autres espèces.
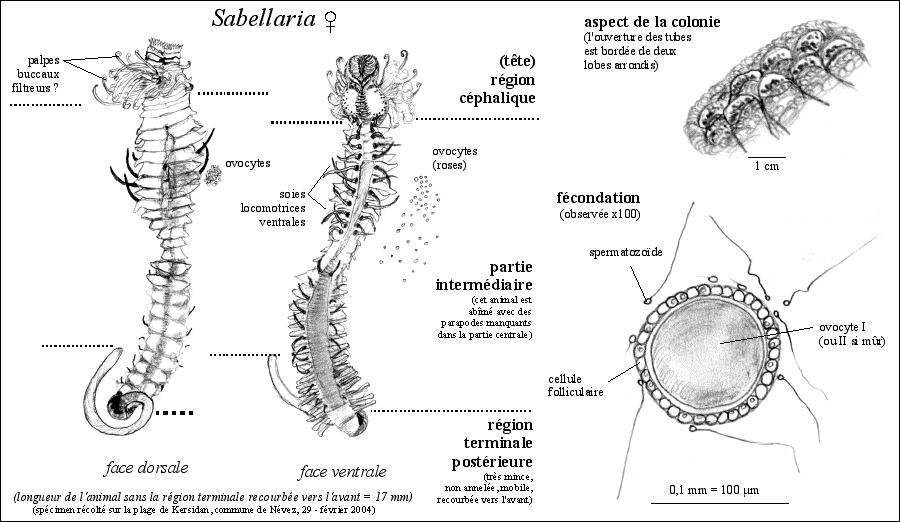
Ces annélides tubicoles sont
caractérisées par une différenciation en
segments non identiques (métamérie
hétéronome). Certains segments ont de très
nombreuses soies alors que d'autres ont des soies réduites.
On peut ainsi comparer cette morphologie
particulière et la mettre en relation avec le mode de vie. On
la comparera par exemple avec l'organisation régulière
de la segmentation et de la disposition des soies chez les
annélides polychètes libres comme la
Néréis. La region terminale est plus mince que
les deux autres régions: la tête, qui porte les palpes
buccaux et la région intermédiaire la plus longue qui
porte peu de soies mais des parapodes (appendices locomoteurs et
respiratoires).
Ces annélides sont filtreurs (consomment des
diatomées planctoniques principalement) grâce aux
tentacules situés autour de la bouche.
Ces annélides tubicoles forment des colonies sur des rochers
ou substats stables (coquilles, galets) dans des zones où l'on
trouve du sable en suspension. On pense que les larves sont
attirés par les substances organiques des tubes des adultes de
la colonie et viennent se métamorphoser à leur
contact.
De nombreuses annélides présentent des
phénomènes de régénération
et de reporoduction asexuée.
Les sexes sont séparés chez la plupart des
Polychètes. Les produits génitaux , ovules (le
terme d'œufs est plus simple et plus
général car il indique toute structure pondue
(libérée); de même que l'on réservera le
terme de zygote pour un œuf fécondé) et
spermatozoïdes, se développent aux dépens
de la paroi péritonéale et tombent précocement
dans le cœlome où ils poursuivent leur
développement. Chez la femelle, les œufs arrivés
à maturité sont pondus soit par rupture du
tégument soit par des papilles néphridiales, la femelle
ne survivant pas à la ponte (les auteurs divergent). Chez le
mâle, l'émission des spermatozoïdes s'effectue par
les orifices des papilles périanales pour certains auteurs et
par rupture du tégument pour d'autres. En tout cas,
placés dans des verres de montre contenant de l'eau de mer,
les mâles libérent une gelée blanche
(à jaunâtre) contenant des spermatozoïdes mobiles
(pendant quelques heures mais pas plus... l'observation doit
être rapide, surtout s'il s'agit de vers conservés au
réfrigérateur; dans ce cas de l'eau de mer propre et
OXYGÉNÉE me paraît utile si l'on veut observer
des spermatozoïdes mobiles et une fécondation) et les
femelles une gelée rosâtre (à violette)
moins abondante contenant des œufs (il est normalement inutile
d'appuyer sur le corps de l'animal pour faciliter une expulsion des
gamètes mais cela peut se faire).
La grande quantité d'œufs produits (stratégie
de reproduction dite de type r, par opposition à la
stratégie de type K caractérisée par la
ponte d'un petit nombre d'œufs de plus grande taille
(réserves !?) et protégées par la femelle) -
voir ci-dessus) en relation avec une
fécondation externe et la dispersion de formes larvaires
(larves trochophores) dispersées au gré des
courants.
Les ovocytes (ou cellules-œufs car ces cellules ne sont pas
forcément matures, c'est-à-dire fécondables)
sont entourés de cellules folliculaires et contiennent des
granules corticaux emplis de glycoprotéines sulfatées.
Lorsque les œufs sont libérés l'ovocyte est
habituellement au stade ovocyte I (première division de
méïose bloquée au stade prophase...?) mais n'est
plus habituellement entouré de cellules folliculaires. A la
fécondation les granules corticaux se déversent
à l'extérieur et conduisent à la formation d'une
gelée protectrice autour de la larve (pas de vraie membrane de
fécondation). Le diamètre moyen d'un ovocyte I
libéré est d'environ 70 micromètres et atteint
100 micromètres lorsque la gangue protectrice
gélatineuse se gonfle. Sur un dessin d'observation fait par
les élèves il ne faut bien sûr pas faire
apparaître membrane plasmique (voir ci-dessous) mais le terme
gangue protectrice. Il semble que chez les hermelles la
fécondation puisse intervenir aussi bien au stade ovocyte I
qu'au stade ovoicyte II ou même au satde ovule.
Les spermatozoïdes sont de forme ovoïdes (limites
peu discernables, ne pas oublier que l'on ne VOIT PAS la membrane
plasmique au microscope optique car elle a une épaisseur
d'environ 8 nanomètres inférieur au pouvoir
séparateur du microscope optique (ou
résolution optique qui est de 0,2 micromètres au
maximum soit 200 nanomètres, QUELQUE SOIT le
GROSSISSEMENT; ce que l'on voit c'est une différence de
biréfringence entre les deux milieux: eau de mer et
cytoplasme... voir par exemple la fiche
de seconde sur le microscope; ne pas
oublier non plus que la membrane plasmique n'est qu'une structure
interprétée et modélisée (un concept !):
voir page sur les modèles).
Le flagelle peut aussi est discerné par biréfringence.
La taille de la cellule est d'environ 6 micromètres sans son
flagelle et de 30 micromètres avec son flagelle.
(Toujours au sujet de la différence
entre ce qui est observable et ce qui est extrapolé, il n'est
à mon avis pas souhaitable que dans un dessin d'observation on
fasse apparaître le terme membrane plasmique en
légende; c'est une limite de la cellule mais
pas la membrane plasmique qui est un concept correspondant
à une structure qui n'est pas visible au microscope otptique.
Par contre sur un schéma d'interprétation il n'y a pas
d'obstacle à faire figurer membrane plasmique en
légende).
La première division (segmentation spirale) du
zygote en embryon semble intervenir au bout d'une heure
environ après la formation du zygote vrai par caryogamie
(environ 75 min après la fécondation). Les
premières larves nageuse en sont obtenues que 48 heures plus
tard (le stade trochophore est situé dans les 150
heures après la fécondation). Ce qui signifie que tout
d'abord, lors d'un TP, il faut garder les stades embryonnaires dans
de l'eau oxygénée et faire des
prélévements successifs pour voir les
différentes étapes mais on ne pourra pas les suivre
directement sur le même individu. Un lame à
concavité pourrait permettre une observation plus longue.
Le développement est indirect: la larve trochophore,
pélagique, qui nage en pleine eau et se nourrit
d'unicellulaires plactoniques (Diatomées, comme l'adulte),
s'allonge puis donne naissance à une larve benthique qui rampe
sur le fond à la recherche d'un substrat solide et d'un
environnement favorable (sable en suspension). Le chimiotactisme
positif vis-à-vis des protéines du tube des adultes
formant la colonie facilite l'installation de nouvelles larves au
voisinage de la colonie constituée. La larve fixée
cesse de s'alimenter, subit sa métamorphose, les segments se
développent et l'animal construit un tube.
Un site à visiter sur le développement embryonnaire
de l'hermelle: http://www.snv.jussieu.fr/bmedia/SiteSabellaria/Sabelbm.htm
des photos légendées y sont
présentées
retour accueil, formulation
par cycle
se multiplier - seul ou à
deux - une flèche de vie - panorama