Mouvements
retour accueil,
concepts en bref / déplacement
des animaux / formulation par cycle / observations
par cycle
1. concepts généraux sur le mouvement
en bref
Une expression du travail du vivant souvent prise comme
caractéristique des êtres vivants est le
mouvement. "C'est un être vivant parce qu'il
bouge". Ce qui ne veut pas dire bien sûr qu'un organisme
immobile n'est pas vivant, c'est d'ailleurs surtout un
problème d'échelle.
Le mouvement ne fait pas uniquement partie du travail de
relation, même si l'on considère la locomotion
d'un animal, car selon la finalité biologique il peut
être plutôt intégré à un
travail de nutrition (recherche de nourriture par exemple
ou circulation du sang ou de la sève) ou, à
un travail de reproduction (recherche d'un
partenaire sexuel ou déplacement d'un gamète). Il ne
faut donc pas hésiter à parler du déplacement
pour quelque chose (ou
de l'adaptation à): les oiseaux migrent pour chercher
des conditions plus favorables de disponibilité de
nourriture..., les sabots des ongulés ont une forme
très efficace pour une course rapide sur une surface
dure..., la forme en fuseau d'un dauphin ou d'un poisson est
particulièrement adaptée à la nage...
Remarque:
Une finalité très anthropomorphique souvent
citée est le jeu : les biologistes citent souvent les
jeux des mammifères pour lesquels la maturation du
système nerveux est lente et se poursuit longtemps
après la naissance, mais certains considèrent le jeu
chez de nombreux vertébrés (voir par exemple dans la
Hulotte : les jeux des chocards à bec jaune (n°50, p
33-39), des petits faucons pèlerins (n°46/47, p30) ou
d'une hermine (n°24, p16-17)). Les interprétations
habituelles sont la libération d'un excès
d'énergie, l'entraînement et le pré-exercice,
mais l'interprétation désormais en vogue (qui va avec
l'évolution sociale) se fonde sur la liberté (au sens
de non détermination) et la gratuité (au sens de non
production, de non rentabilité) : on parle ainsi volontiers du
jeu comme permettant l'établissement du lien social. Cette
terminologie a le gros défaut de mélanger le
comportement animal (éthologie) et la psychologie humaine qui,
pour des raisons philosophiques, me semblent devoir être
séparées. Je ne crois pas qu'il faille aller chercher
dans le monde animal des justifications aux comportements humains.
Par contre il est clair que l'homme n'a que sa raison et ses sens
pour comprendre le monde animal, dont il participe (j'avais
essayé pour les élèves de seconde de montrer
comment je vois cette séparation dans l'homme
et le kangourou). Pour un approfondissement consultez par exemple
les articles "Le jeu des animaux" et surtout "Le jeu chez l'enfant"
dans l'Enyclopédia universalis par exemple.
a. vocabulaire
Un peu de vocabulaire afin de fixer le domaine d'utilisation de
chaque mot et pouvoir ainsi exprimer les concepts :
- le mouvement est au sens propre "un changement de
position dans l'espace en fonction du temps, par rapport à
un système de référence" (Petit Robert, 1984)
: on peut alors définir, au sein d'un repère
d'observation, un état initial et un état final qui
doivent être différents (les plantes
présentent des mouvements à l'intérieur de
leurs cellules, entre leurs cellules au sein de l'organisme mais
aussi des mouvements d'ensemble de certains organes comme les
vrilles qui sont souvent des feuilles modifiées capables de
s'enrouler autour d'un support).
- le moteur est ce qui engendre le mouvement (le
moteur de la course de l'homme est le muscle qui réalise la
contraction musculaire ; le moteur du déplacement d'une
bactérie comme Escherichia coli est le flagelle qui est
animé d'une rotation extrêmement rapide) ; il est
évident qu'en biologie les moteurs ne sont que des
causes secondes et non des causes premières; ce qui
fait que l'on peut toujours trouver un moteur du moteur; le terme
de moteur étant basé sur une analogie
mécanique, en biologie (biomécanique) on se
contentera de parler du moteur comme structure
génératrice du mouvement; les aspects
énergétiques (disponibilité
métabolique de la source d'énergie), même
nécessaires, sortent alors de la définition, et sont
considérées comme des moyens, tout comme la commande
motrice et son intégration.
- la mobilité est un caractère (pour un
individu) ou la propriété (pour une structure) de ce
qui peut se mouvoir ou être mû (un être
vivant est souvent doué de mobilité)
- un mobile étant un corps qui peut se mouvoir ou
être mû (les êtres vivants sont souvent
mobiles mais parfois immobiles)
- la motilité est la faculté de se mouvoir
et doit donc être réservée aux individus
(les animaux sont des êtres vivants motiles, au moins
pendant une période de leur développement ; par
contre on devrait prendre soin de dire structure mobile (qui
confère la motilité ou mobilité de
l'organisme) et non motile (qui a la faculté de se
mouvoir), même si, considéré du point de vue
de la structure, l'usage en est possible, malgré le fait
qu'il revient à individuer la structure)
- la motricité est l'ensemble des fonctions
assurant les mouvements (l'étude de la motricité
musculaire comprend aussi bien celle de la circulation, la
respiration, la contraction musculaire, le métabolisme du
muscle au sens large....)
- se mouvoir se dit de la capacité à se
déplacer, c'est-à-dire changer de place et donc
d'effectuer un mouvement
- la locomotion est un nom plus récent est
désigne aussi l'action de se mouvoir pour un être
vivant.
b. concepts
Selon l'échelle d'observation (et donc le repère
d'espace et de temps) un corps peut sembler immobile ou mobile :
à l'échelle de l'écosystème côtier
une moule est immobile une fois fixée à son support ;
à l'échelle du rocher, la moule est capable de se
déplacer grâce à son byssus.
A l'échelle cellulaire, les structures et mécanismes
générant les mouvements ont une uniformité
étonnante (la distinction la plus prisée actuellement
étant celle que l'on peut faire entre une cellule procaryote
et une cellule eucaryote).
A l'échelle des organes et surtout des organismes, les
modalités des mouvements sont très nombreuses, ce que
l'on peut mettre en rapport avec la diversité des
finalités biologiques des mouvements ainsi que des milieux de
vie.
En terme de finalité biologique, je ne suis pas
sûr que la distinction classique entre mouvement actif
(se mouvoir) et mouvement passif (être mû)
reflète autre chose qu'une différence d'échelle.
Le déplacement du pollen par le vent (pollinisation
anémophile) est l'utilisation d'une caractéristique du
milieu (le vent) et de caractéristiques de la structure
vivante (le pollen, extrêmement léger) pour
réaliser une certaine finalité biologique qui est la
pollinisation d'individus éloignés. Il est vain de
parler de la "volonté" du grain de pollen mais il est
légitime de parler du "déplacement anémophile"
de celui-ci. Il n'y a pas plus ni moins de finalité biologique
dans le déplacement d'un pigeon. Les mécanismes (moyens
pour atteindre le but ?) sont par contre différents en ce qui
concerne l'énergie dépensée notamment : l'un est
un "vol plané", l'autre est généralement un "vol
battu".
Comme pour tous les systèmes vivants (si
vous désirer approfondir, vous pouvez consulter les pages sur
les modèles)
on peut utiliser la distinction entre modèle externe et
interne. Le modèle externe décrit le mouvement
de l'organisme à partir des observations globales de celui-ci
alors que le modèle interne s'efforce d'atteindre les
mécanismes internes déterminant le mouvement. Comme
j'ai essayé de la montrer à l'occasion des
études de physiologie du cours de terminale, notamment sur
la pression artérielle
(introduction), on peut présenter la physiologie sous un
double aspect : le travail du vivant est à la fois une
réponse adaptative de l'organisme au milieu (travail
social, externe) et une expression de déterminismes
internes (capacité au travail, travail interne). Cette
distinction est à mon avis éclairante même pour
une formulation simple.
c. modèles externes
Du côté des modèles externes on trouve
la description cinématique des déplacements mais aussi
les études de biophysiques reliant les principaux
paramètres du mouvement d'un organismes aux composantes du
milieu.
Du point de vue cinématique (partie de la
mécanique qui étudie les mouvements des systèmes
matériels), les mouvements sont définis, du moins en
cinématique classique (où l'on définit un solide
invariable et un temps), sont définis donc par une
trajectoire, une vitesse et une accélération (ou
plusieurs, ce ce ne sont pas des mouvements uniformes) et une
durée. Les mouvements élémentaires étant
des translations, des rotations. Les mouvements complexes sont
définis par des outils mathématiques (torseurs).
Du point de vue énergétique le mouvement actif
est la résultante de la conversion de l'énergie
chimique en énergie mécanique: le rendement
énergétique ne dépasse guère 25%, les 75%
de l'énergie chimique consommée étant
évacués sous forme de chaleur (l'activité
musculaire produit beaucoup de chaleur).
Remarques sur la contraction musculaire
(d'après Schmidt-Nielsen)
La force développée par les muscles est
absolument indépendante de leur longueur mais dépend
directement de leur section. La force maximale
développée par des muscles (lisses ou striés)
composés de filaments d'actine et de myosine que l'on retrouve
chez les vertébrés et le plupart des
invertébrés est de l'ordre de 40 à 50 N.cm-2.
Lorsque l'on voit une fourmi soulever une masse plus lourde qu'elle
on peut estimer que ses muscles sont plus puissants que ceux de
l'homme, ce qui est faux dans l'absolu mais vrai relativement
à la taille. Le volume de l'animal est proportionnel à
la puissance cubique de sa taille. La section transversale d'un
muscle est par contre proportionnelle au carré de sa taille.
Lorsque l'on compare la force exercée chez deux animaux de
même forme mais de taille par exemple divisée par 200
(homme-fourmi), le volume est divisé par 8.000.000
(200x200x200), alors que la section du muscle n'est divisée
que par 40.000 (200x200) ce qui fait une force qui apparaît 200
fois plus importante, relativement (au volume de l'animal). Plus la
taille d'un animal augmente, plus sa force relative (ramenée
à l'unité de masse corporelle) diminue.
A l'inverse de la force maximale qui ne dépend que de la
section, le travail fourni (produit de la force par la
distance) dépend par contre directement de la longueur du
muscle.
Au total le travail possible d'un muscle est directement
proportionnel à son volume (produit de sa section par sa
longeur, en le supposant de forme globalement cylindrique).
La puissance est déterminée par le travail par
unité de temps, ce qui revient à faire le produit de
la force par la vitesse de contraction. On peut dire
approximativement qu'un muscle rapide développe plus de
puissance qu'un muscle lent. Or la vitesse de contraction est souvent
dépendante du métabolisme de l'animal et inversement
proportionnel à la taille (plus l'animal est petit, plus ses
muscles sont rapides).
Les muscles adducteurs des Bivalves sont composés
à la fois de fibres lisses et de fibres striées. C'est
la partie striée, seule présente dans les muscles
rétracteurs du byssus de la moule par exemple, qui semble
être capable de rester contractée de façon
continue sans nouvelle stimulation nerveuse (il s'agit d'une
tétanie particulière nommée catch dont le
mécanisme précis n'est pas encore éclairci).
Les muscles striés de la pince de crabe sont aussi
intéressants par leur disposition en plume (obliquement de
part et d'autre d'un axe central) qui leur permet de se contracter
dans le volume restreint délimité par la carapace non
extensible. Cette disposition se retrouve sur d'autres muscles de
vertébrés.
Quelques éléments de mécanique sur les
caractéristiques (qualifiées souvent de
contraintes) des différents milieux:
- l'eau est un milieu plus dense
(on dit parfois lourd de façon inadéquate) et
surtout plus visqueux que l'air. Les forces
nécessaires au déplacement du milieu (pour la
respiration) qui font référence à la
densité sont plutôt ici les forces nécessaires
au déplacement d'un corps dans le milieu et donc en
relation avec la viscosité. C'est la viscosité de
l'eau (forces tendant à s'opposer à la
déformation de la masse d'eau dans une direction
donnée) qui est responsable de la poussée qui permet
à un poisson de nager par ondulations par exemple. Pour des
organismes de petite taille les forces de viscosité
(tension de viscosité) sont de loin les plus importantes
pour expliquer son déplacement. Pour un organisme de grande
taille, la nage repose plus sur l'utilisation de l'eau comme
milieu portant, le déplacement des masses d'eau et surtout
l'inertie des masses d'eau déplacées. La
densité de l'eau étant plus voisine de celle de
l'organisme que l'air, les problèmes de maintien dans le
milieu sont par contre facilement résolus. La
flottabilité d'un organisme peut ainsi être
réalisé grâce à des mécanismes
lui permettant de modifier sa densité à
volonté (réserves de graisses, vessies natatoires
pleines de gaz....). La vitesse de déplacement dans l'eau
est surtout limitée par les frottements importants entre
l'organisme et l'eau, ce qui nécessite une dépense
d'énergie très forte, malgré la forme
hydrodynamique (qui tend à réduire les frottements)
de nombreux organismes aquatiques de grande taille.
- l'air est un milieu de faible
densité (on dit parfois léger de façon
inadéquate) et de faible viscosité. Si le
déplacement dans l'air est facile, demande peu
d'énergie, la portance de l'air est par contre
faible ce qui nécessite d'augmenter la surface
spécifique (surface rapportée à la masse). La
poussée, force exercée sur le corps en
mouvement dans l'air, par les masses d'air
déplacées, dépend beaucoup de la taille de
l'organisme. Elle est très faible pour des organismes de
petite taille qui ont du mal à ne pas être le jouet
des caprices du vent (qui lui, ne correspond jamais à des
forces néglgeables, les événements
récents en France et familiers à d'autres pays, en
sont les témoins). La poussée est par contre plus
importante pour des organismes de grande taille déplacant
de grosses masses d'air. Les forces de frottement dans
l'air peuvent paraître négligeables à faible
vitesse mais elles deviennent vite prépondérantes
lors d'un vol en piqué ou d'un vol au long cours d'un
oiseau migrateur. Elles ne sont pas non plus négligeables
pour un animal qui court à vitesse
élevée.
Il est important de noter que la plupart des organismes
"aériens", qui se déplacent dans l'air, se posent
à un moment ou à un autre sur un substrat solide et
présentent donc d'autres modes de locomotion
éventuellement.
- le milieu terrestre est varié :
la déplacement terrestre le plus courant est celui d'un
organisme à la surface de la terre; dans ce cas la
locomotion varie en fonction de la cohésion du
substrat qui permet de développer une force de
réaction à la gravité exercée
par les organismes en contact avec le substrat (elle permet aussi
d'emmagasiner de l'énergie élastique dans des
structures dites élastiques): mais aussi de diminuer les
forces de frottements qui sont assez faibles sur des
substrats durs mais très fortes sur de la vase par exemple;
l'essentiel du travail fourni par l'organisme est celui
exercé sur ses propres structures; mais on peut aussi
considérer les organismes fouisseurs ou les innombrables
organismes qui vivent fixés sur d'autres organismes
aériens ou aquatiques...
d. modèles internes
Du côté des modèles internes, on trouve
d'abord la description du fonctionnement intégré des
structures mobiles mais on peut aussi étudier de cette
façon les relations de mobilité entre organismes au
sein de l'écosystème en tenant compte des diverses
finalités individuelles et de leur interactions.
Bref panorama des mouvements au sein du monde vivant :
- Les bactéries ne semblent pas
avoir de mouvements internes de structures dans leur
cytoplasme : pas de vésicules en transit, pas de
division d'organites, inexistants... mais, il est cependant
certain que l'on ne peut concevoir de métabolisme sans flux
moléculaire, ce qui se traduit au moins par des
mécanismes de diffusion. Si les protéines
membranaires ne se renouvellent peut-être pas (vite ?) ,
lors de la croissance nécessaire à la phase
préparatoire à une division par scissiparité,
il y a bien expression-traduction du génome et donc des
mouvements moléculaires qui se traduisent pas des
mouvements cellulaires comme l'élongation ou le
rétrecissement de la zone de scission. Par contre on n'a
pas trouvé chez les bactéries de protéines
voisines de l'actine ou de la myosine des eucaryotes.
De plus, de nombreuses bactéries sont capables de se
déplacer au moyen de flagelles (le flagelle
bactérien ne cesse d'être étudié:
il semble que sa rotation extrêmmement rapide de plusieurs
dizaines de milliers de tours par minute soit obtenue grâce
à un rotor constitué par une molécule d'ATP
synthase qui utilise le flux protonique transmembranaire pour
réaliser sa rotation rapide) ou par glissement.
- Les unicellulaires sont doués
tout d'abord de mouvements intracytoplasmiques
extrêmement riches : traffic des vésicules de
transition du reticulum lisse vers le golgi, et des
vésicules golgiennes du golgi vers la membrane
cytoplasmique ; ces mouvements sont réalisés
grâce à des molécules motrices
(dynéine(des cils et flagelles) et myosine (des muscles)
par exemple qui convertissent de l'énergie chimique en
énergie mécanique), à des molécules
polymérisables comme les microtubules, ainsi
qu'à un réseau de guides (microfilaments) qui
structurent le cytoplasme et forme le cytosquelette. Les
phénomènes de mouvements intracellulaires sont
facilement observables au microscope dans une classe : la vacuole
contractile de la paramécie ou le mouvement de "cyclose"
des chloroplastes de l'Elodée par exemple.
Les unicellulaires sont aussi doués de mouvements
cellulaires au sein de leur milieu grâce à des
structures complexes comme les flagelles ou les cils
(composés d'un axe de microtubules très
ordonnés au sein d'une expansion cytoplasmique plus ou
moins rigidifiée dont le battement peut être
synchronisé entre flagelles ou cils voisins), mais aussi
à l'émission de pseudopodes (expansion
cytoplasmiques déformables dont les mécanismes
d'émission-rétraction utilisent aussi des
microtubules cytoplasmiques associés à des filaments
du cytosquelette). D'une façon très
générale, tous les mouvements des cellules par
déformation - mouvements qualifiés
d'amoeboïdes par référence aux amibes
(Amoeba protens, étant l'amibe commune des intestins
humains par exemple) - qu'ils donnent une impression de
glissement, de reptation ou de roulement, semblent basés
sur des mécanismes identiques de déformation du
cytoplasme réalisés grâce aux cytosquelette et
aux microtubules.
- Les champignons, bien que munis d'une
paroi (chitineuse et donc souple) qui s'oppose partiellement
à la déformation cellulaire, sont capables de
coloniser de très grands volumes de substrat en
étalant leur mycélium (filament qui constitue la
forme la plus habituelle de la structure pluricellulaire des
champignons). Les phénomènes de croissance,
bien qu'assez lents, n'en sont pas moins très performants,
voir par exemple la vitesse de croissance d'un "chapeau"
(carpophore) d'une champignon de sous-bois ! Certains champignons
possédent des phases unicellulaires flagellés
lors de la reproduction sexuée (gamète ou cellule
sexuelle mobile).
- Les plantes, bien que munis d'une paroi
(cellulosique) - qui devient assez rigide par imprégnation
de substances comme la lignigne, après avoir
été souple, lorsqu'elle n'est composée que de
cellulose et de pectines - sont, elles aussi, capables de
phénomènes de croissance très rapides
(plantes annuelles par exemple dont l'appareil
végétatif pousse en quelques semaines) et l'on
connaît aussi les mouvements classiques d'enroulement des
tiges (plantes volubiles comme le houblon) ou des feuilles
(vrilles du pois ou de la vigne par exemple) autour de support.
D'une façon générale on distingue chez les
plantes les nasties (flexions d'organes
déterminées par la structure anatomique de l'organe
lui-même et non par une différence dans le milieu; un
exemple type : l'ouverture ou la fermeture de fleurs), les
tropismes (réactions d'orientation des organes dues
à des inégalités de croissance entre
différentes parties du végétal et
liées à une anisotropie du milieu,
c'est-à-dire une différence de valeur d'un
paramètre du milieu entre différents points de la
plante ; l'exemple classique est le phototropisme,
mécanisme d'orientation de la fleur de tournesol par
exemple en fonction des mouvements relatifs du soleil au cours de
la journée) et enfin les tactismes
(déplacement de l'organisme entier). Les tactismes sont
très limités chez les végétaux
à l'échelle de l'organisme et de son milieu
immédiat (en fait seules les gamètes, souvent
flagellés dans de nombreux groupes, ou les
structures qui les comportent comme le grain de pollen, ont des
véritables déplacements importants), par contre
la colonisation des milieux et la répartition des
espèces en fonction des conditions du milieu montrent de
véritable déplacements orientés à
l'échelle de quelques générations, souvent
obtenues par multiplication asexuée.
A l'échelle de l'organisme il existe d'autres mouvements
comme les mouvements de circulation des fluides au sein du
végétal : sèves (brute dans des vaisseaux
constitués de cellules mortes ; élaborée
à travers le cytoplasme de cellules vivantes) ou gaz (dans
les espaces lacunaires entre les cellules).
- Les animaux sont sans doute les
organismes doués de mouvements les plus riches :
locomotion rapide dans tous les milieux (eau, air, terre) tout
en ayant des mouvements cellulaires internes au moins aussi
riches que les autres groupes d'organismes (circulation sanguine
et lymphatique, péristaltisme digestif, mouvements
ciliaires des épithéliums des voies respiratoies ou
des trompes chez la femme...). Les mécanismes de locomotion
sont tous basés sur la contraction musculaire
réalisée grâce à des structures
protéiques contractiles de type myosine-actine.
2. le déplacement des animaux
Etudier le déplacement pour lui-même n'a pas beaucoup
de sens en primaire et il est certes préférable de
relier le déplacement à un travail (nutrition,
relation-défense, ou reproduction).
Les mouvements de l'homme ont déjà été
étudiés dans le chapitre "je peux
bouger"; l'accent notamment a été mis sur
l'unité entre les différents éléments
nécessaires au mouvement musculaire : muscles (et
tendons), squelette articulé et système
nerveux (de commande et de perception).
Par milieu, à partir d'exemples et en partant de ce que
l'enfant peut le plus facilement imaginer par référence
à son corps, on peut étudier différents
modes de déplacement: marche, course, saut, reptation,
fouissement, nage, vol.
Sources:
* La Hulotte voir par exemple:
- sur le vol des oiseaux :
acrobaties aériennes (de l'autour: n°27, du chocard:
n°50, du faucon pélerin : n°45), le balbuzard
fluviatile: n°42/43, le busard saint-martin n°20, la
buse variable n°27, 40 et 41, le butor étoilé
n°30, le faucon crécerelle n°31, le faucon
pélerin n°42/43 et 46/47, le grand corbeau, le corbeau
freux, et la corneille noire n°50, le coucou gris n°38
et 39, l e baron gris (busard cendré) n°63,
l'adaptation de l'hirondelle à la vitesse n°42/43 et
67 ou ses sorties alimentaires pendant son temps de couvaison
n°62, ou ses migrations n°6, les acrobaties de la
sitelle n°8, le martinet n°13, les pics n°11, la
pie n°16, voir aussi les migrations (n°6 et 70
(hirondelles), 13 (libellules), 18 (foulque), 24 (oies
cendrées), 38-39 (coucou), 50 (corneille mantelée),
57 (grues), 67 (héron cendré)), la grue et la
cicogne n°56
- sur la nage:
des poissons : la chasse du brochet n°30, ou la nage des
canards n° 18 ou d'autres oiseaux aquatiques n°27: la
foulque n° 18, la grèbe huppée n°71 et 72;
ou encore de reptiles comme la tortue d'eau douce n°75; et
d'amphibiens : la grenouille rousse n°19; enfin la "nage"
d'invertébrés comme le gyrin n°21 ou la
notonecte n°21
- sur les mammifères :
la chasse de la belette n°41, le chat n°24, de la loutre
n°18, du renard n°31 et 33/34, du sanglier n°31, du
chat sauvage n°12 et 41, les acrobaties de l'écureuil
n°36/37, voir aussi l'index à empreintes, la course du
lièvre n°15, les deux façons de courir des
petits rongeurs n°27, les 4 rongeurs qui montent le mieux aux
arbres n°18, la fuite du chevreuil n°66, le
hérisson n°77
- sur d'autres vertébrés :
les bonds du crapaud accoucheur n°53 , la marche sur l'eau du
cincle n°10, les deux façons de marcher des petits
oiseaux n°27
- et d'invertébrés:
chasse de la larve de dytique et de la nèpe n°21, de
la larve de libellule n°13 et 21, de l'araignée
Pholcus n°54, de la chrysope n°19, la marche sur l'eau
du gerris ou du gyrin n°21
* Collection : "L'éveil par les activités
scientifiques" dirigée par Raymond Tavernier, Bordas,
1975: "Les animaux, les élevages",Guide du maître
(les animaux se déplacent : p 167-195)
- fiches de 2ème niveau : 1.1 (des traces sur le sable);
1.2; 2 (marche du cheval); 3 (marche du chien); 4 (saut de
l'homme); 5 (saut de la grenouille); 6.1 (comment sautent-ils ?);
6.2; 7 (le kangourou se déplace)
- fiches de 3ème niveau : 4.1 et 4.2 (les membres
postérieurs); 5.1 et 5.2 (les membres antérieurs); 6
(ailes); 10.1, 10.2 et 10.3 (les migrations des oiseaux)
* Enseigner la biologie et la géologie à
l'école élémentaire, Guide des professeurs
des écoles, I.U.F.M., R. TAVERNIER, Larousse-Bordas, 1996
(Chapitre 3 : les êtres vivants se déplacent, p
233-278);
* Le Moniteur des sciences, Bernadette Bornancin, cycle 3,
"le monde vivant", niveaux 1, 2 et 3, (fichiers d'expériences
1 et 2 et fichier ressources), Nathan, 1998; un sujet un peu
délaissé et surtout des fiches vraiment pas
convaincantes
Les compléments qui suivent sont des informations pour le
maître et n'ont pas pour but d'être utilisées
directement pour des séances.
|
La marche du chien
|
|
|
marche à 4 pattes (quadrupède, c'est un
vertébré tétrapode) sans intervention
directe de la queue, une patte (les trois autres restant au
sol) puis deux pattes (l'animal reste en équilibre
sur deux pattes, une de chaque côté) sont
soulevées en même temps alternativement ;
marche sur les doigts (digitigrade)
|
* observation très difficile sans un support
graphique, même à partir d'un animal vivant ou
d'un film
* la notion d'équilibre peut aussi être acquise
plus facilement avec des modèles déformables
en caoutchouc que l'on plie à volonté
* le travail sur les traces-empreintes est très
intéressant car il permet de voir où se situe
le poids du corps de l'animal à un moment
donné (équilibre) et quelles parties de la
patte reposent au sol (marche digitigrade).
* le travail de mime permet de comprendre la notion
d'équilibre mais aussi celle de coordination.
* pour un cycle 2 ou 3 on peut même aller
jusqu'à un travail d'anatomie comparée sur la
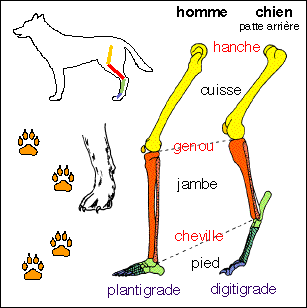
position de chaque segment et articulation. On prendra garde
à la position de l'articulation de la cheville
placée très haut chez le chien et que les
enfants ont tendance à confondre avec le genou.
* on peut aussi faire un travail de mesure de la vitesse de
déplacement du chien (estimation de la longeur d'une
foulée)
* ce peut être un excellent exercice
d'évaluation que de demander un dessin d'un chien au
repos (4 pattes reposant au sol) et lors de la marche
(l'aspect dynamique du dessin peut aussi tenir compte de la
position de la tête et de la queue qui ne sont pas
forcément quelconques).
|
|
"film" dessiné Tavernier, 1996, p241
fiche du maître, Tavernier 1975, p168-169 et fiches
de l'élève : 2ème niveau 1.1,1.2, 3
|
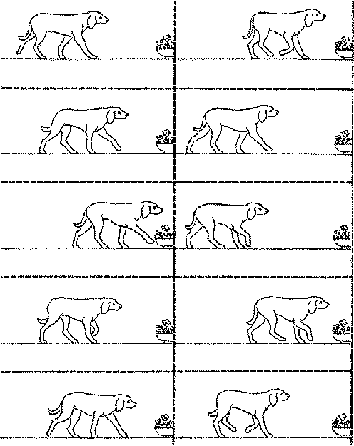
|
|
de la course du chien au galop du cheval ou du
lévrier en passant par la course bondissante des
antilopes
|
|
Lors de la course une bonne partie du travail est
du à la montée et à la descente du
centre de gravité à chaque foulée. Un
moyen pour ne pas trop perdre d'énergie
cinétique lors de ces mouvements est de la stocker
sous forme élastique (rebond).Ce travail est bien
sûr augmenté lors d'une course en montée
(voir généralités ci-dessus, le travail
est presque doublé pour un chien qui doit courir sur
un plan incliné de 15°).
De plus, le travail d'allongement et de retrait de chaque
membre est nettement plus important que dans la marche car
il se fait avec une plus forte accélération
(en phase d'avancée) et de
décélération (en phase de retrait).
Les frottements contre un sol ferme sont négligeables
à l'inverse des frottements sur un sol meuble (sable
par exemple).
Les frottements contre l'air sont assez importants à
vitesse élevée (pour un lévrier mais
pas pour un caniche !).
Lors d'un galop ou d'une course bondissante, le travail
des membres est identique à celui de la course.
|
Le galop est défini comme une course
basculée à trois temps; il se
décompose en:
- poser du postérieur gauche;
- poser simultané du postérieur droit et de
l'antérieur gauche;
- poser de l'antérieur droit.
Le galop étendu est à quatre temps
comme le galop ralenti: le poser de l'antérieur
gauche et du postérieur droit sont
dissociés.
Les formes du galop peuvent considérablement varier,
en fonction tant de la vitesse que de
l'équilibre.
Lorsque la phase pendant laquelle aucun des membres ne
touche le sol s'allonge l'animal progresse par une
série de bonds successifs : on peut parler de
course bondissante .
|
|
Lors d'un sprint chez l'homme on estime à
13% les pertes d'énergie dues aux frottements de
l'air (frottements aérodynamiques).
|
|
Du point de vue de l'anatomie comparée
(voir par exemple A. Beaumont et P. Cassier,
Biologie animale, Dunod, 1987) et uniquement en ce
qui concerne le squelette, l'adaptation à la
course quadrupède (chez les
vertébrés) est réalisée par
:
- un allongement des membres qui permet principalement
d'augmentéer la longueur de la foulée
- le relèvement progressif de la partie terminale de
la jambe (autopode) : c'est-à-dire que la course est
d'autant plus rapide que le pied avant ou arrière
repose moins à plat sur le sol : la course est moins
rapide chez les plantigrades (homme, mais pour lui s'ajoute
la bipédie : fait de marcher sur deux jambes et non
sur quatre; hérisson, ours) que chez les digitigrades
(chien, chat, nombreux rongeurs...) et surtout que chez les
onguligrades (cheval, cerf, antilope...).
- la réduction du nombre de doigts (4 doigts porteurs
chez l'hippopotame, 2 doigts porteurs et deux doigts
latéraux chez le porc, uniquement deux doigts
porteurs chez la vache, et une seul doigt chez le
cheval).
|
|
sauter ...
|
|
On retrouve l'importance de
l'élasticté au niveau des muscles des
vertébrés, y compris chez l'homme, tout comme
dans le vol (voir ci-dessous). Lorsqu'une balle de tennis
s'écrase au sol, sa déformation absorbe une
partie de l'énergie cinétique et la restaure
lors du rebond.
De façon analogue, les composants élastiques
des muscles, des tendons et du squelette de la jambe, sont
capables d'emmagasiner une partie de l'énergie
cinétique lors que l'on fléchit par exemple
les jambes et la restaurer lors de l'extension
"rebondissante". (Des mesures réalisées
chez un homme ont montré qu'il y avait une
économie de l'ordre de 22% de la consommation de
dioxygène s'il utilisait le rebond (par rapport
à une très courte pause entre la phase de
flexion et la phase d'extension) ce qui correspondrait
à une variation de 0,19 à 0,6 du rendement
énergétique). Lors d'un exercice de petits
sauts à pieds joints et jambes tendues,
l'énergie cinétique est aussi stockée
sous forme élastique dans les structures des pieds et
des mollets. Celle-ci est estimée à 60% du
total de l'énergie cinétique de
déplacement du corps.
Lors de la course d'un chien filmé et dont on
enregistre les forces exercées sur le sol au point
d'appui avant le saut d'un obstacle de 3m, on a pu mettre en
évidence que la composante élastique est
probablement stockée dans les tendons des pattes
postérieures, dernières à être en
contact avec le sol. Des résultats analogues ont
été obtenus pour le kangourou.
Pour le saut des puces (capable
d'accélérer à près de 2 km.s-2
soit 200 g, g étant l'accélération de
la pesanteur) et d'autres insectes comme les taupins
(capables de se retourner brsuquement lorsqu'ils sont
placés sur le dos), il a été mis en
évidences des structures élastiques
articulaires (avec une protéine qui est la
résiline) suceptibles de stocker l'énergie
cinétique et de la libérer brusquement par
déclic (système voisin d'une catapulte).
Le record de saut en hauteur semble être celui du
galago, petit lémurien d'Afrique qui pour 250
g saute 2,25m de haut, départ arrêté
(soit trois fois plus qu'un homme ou qu'une puce). Sa
capacité semble être due principalement
à sa très grande masse musculaire.
|
|
le saut de la grenouille, du lièvre et du
kangourou...
|
|
Les dessins proposés par Tavernier pour
étudier le saut de la grenouille décomposent
le saut d'une grenouille depuis la terre ferme dans une
mare, ce qui n'est pas un saut mais un plongeon.
La grenouille est tout aussi capable de sauter en faisant
un bond et en retombant en équilibre sur ses pattes
arrières, comme la kangourou, sans se servir de ses
pattes avant pour la réception au sol.
|
Deux constantes semblent se retrouver pour les animaux
adaptés au saut:
- la forme de la patte "en Z", c'est-à-dire une
longueur équivalente entre les trois segments des
pattes arrières adaptées au saut
- la présence de systèmes musculaires,
tendineux ou articulaires de stockage de l'energie
élastique
|
|
grimper, escalader, courir en montée ...
déplacer son centre de gravité vers le haut...
un travail épuisant
|
|
Le travail nécessaire pour
élever le corps d'une hauteur h, que ce soit lors
d'une course en montée ou d'un grimper, est
proportionnel à la masse de l'animal (et non
à sa taille). C'est pour cela que le coût
métabolique (énergie dépensée)
rapporté à une unité de masse pour
monter de 10 cm est le même pour un cheval que pour
une souris. On a ainsi mesuré que lorsqu'une
souris de 30 g court sur un tapis roulant
incliné à 15° elle ne consomme
guère plus de dioxygène que lorsqu'elle court
sur une surface plane. Pour un chimpanzé de 17
kg la consommation de dioxygène double lors de la
course sur le plan incliné à 15° par
rapport au même plan horizontal. Les calculs
réalisés pour un déplacement vertical
à 2 km.h-1 donnent une augmentation de la
consommation de dioxygène de 23% chez la souris, 189%
chez le chimpanzé et 630% chez le cheval. Il
est donc aisé de comprendre pourquoi
l'écureuil, du fait de sa petite taille,
grimpe aisément aux arbres, ce qu'un singe
saïmiri peut encore faire mais qu'il devient
impossible à un orang-outan de réaliser
sans un très gros effort.
|
|
ramper...
|
|
le lombric (annélide) raccourcit et allonge
successivement les différents segments de son corps,
tout comme le boa (serpent = ophidien) et progresse
parce qu'il prend appui sur le substrat: par des soies (4
paires par anneau) dans le cas du lombric (on peut les
entendre crisser si l'on fait se dépacer le lombric
sur une feuille de papier - on notera que
les soies sont composées de chitine, la
même protéine qui compose principalement la
cuticule des insectes ou la paroi des champignons...)
et par ses écailles dressées pour le
serpent.
|

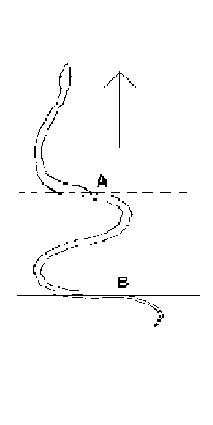
Déplacement d'un lombric (à observer par
dessous une plaque de verre)
in
Schmidt-Nielsen
Le lombric rampe par contractions et
allongements successifs de son corps et en
s'accrochant avec ses soies locomotrices.
Le déplacement du lombric se fait avec de
nombreuses contorsions. Il s'arrête souvent. Avant de
repartir, l'avant (l'extrêmité la plus fine) se
soulève, se balance de droite à gauche, touche
le sol, cherche sa voie.
|
|
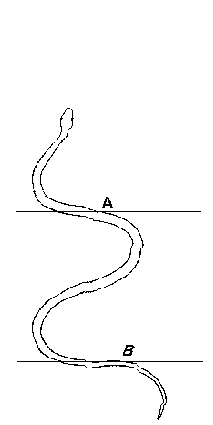
les serpents ondulent aussi fortement leur corps.
Le déplacement du crotale et de certaines
vipères est assez particulier: la partie
antérieure du corps (en avant du point A) reste
immobile et prend appui sur le substrat; en arrière,
la partie restante du corps ondule et se raccourcit. Puis
c'est au tourde la partie située en arrière du
point B de prendre appui sur le sol en restant immobile et
c'est la partie en avant de ce point B qui s'allonge, ce qui
fait avancer le reptile sur le sol (in
PP. Grassé, Précis de Zoologie, Masson, 1976,
t. 2, p 361)
|
|
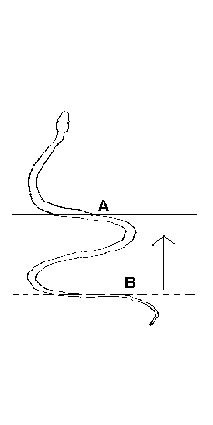
|
|
|
nager ...
|
|
Nager et voler sont des
déplacements très similaires étant
donné que l'air et l'eau sont deux fluides, donc
facilement déformables. L'eau, plus dense, ne permet
pas d'atteindre des vitesses de déplacement aussi
élevées que l'air.
Le battement de la queue ou d'une nageoire latérale
d'un poisson est tout à fait assimilable au battement
d'une aile. La musculature des poissons à une
disposition très particulière (chevrons) et il
semble que l'absence de tendons
(Téléostéens, c'est-à-dire la
majorité des poissons courants) soit compensée
par l'accrochage des muscles à la peau,
elle-même rigidifiée par des fibres de
collagène, qui tire à son tour sur la colonne
vertébrale.
Un mécanisme intéressant pour les poissons
consiste dans l'utilisation de sa vessie gazeuse qui lui
permet d'ajuster sa flottabilité (densité) aux
caractéristiques de l'eau et de la
profondeur.
|
|
la nage de la grenouille
|
la poussée des pattes arrières est
augmentée par la palmure (analogie avec la brasse,
position des pieds...); l'animal flotte aisément du
fait de l'air emagasiné dans ses poumons et son
pharynx. La brusque détente des pattes
postérieures lors de la nage ressemble très
fortement à leur détente lors du saut.
Certaines grenouilles (restant toujours dans l'eau) ont
même les mains palmées.
|
|
la nage des poissons
|
Le corps fusiforme (forme hydrodynamique) favorise la
pénétration dans l'eau, visqueuse, en
minimisant les turbulences. Pour les poissons aplatis
latéralement, comme le saumon, et munis d'une
nageoire caudale verticale, c'est l'ondulation du corps
(soutenu par la colonne vertébrale qui joue le
rôle d'une lame élastique) et prolongé
par la nageoire caudale (queue) qui est propulsive
principalement; le battement (horizontal) conserve le
principe du battement vertical des ailes d'un oiseau. Les
nageoires latérales servent principalement à
la direction. Le centre de gravité d'un poisson est
d'ordinaire non loin de la tête et plus près de
la face dorsale que de la face ventrale, ce qui l'oblige
à utiliser en permanence ses nageoires pour lutter
contre ce déséquilibre.
Les poissons aplatis dorso-ventralement nagent par
ondulation de tout le corps (turbots, soles) ou de nagoires
pectorales hypertrophiées (raies).
Un élément essentiel de la locomotion des
poissons est leur flottabilité. Les poissons
plus denses que l'eau soit accumulent des graisses (de
densité plus faible que la moyenne de leur chairs),
soit possédent une vessie natatoire, soit encore
compensent la tendance à tomber au fond en nageant
sans cesse. Les poissons munis de vessie natatoire essaient
d'ajuster (par des changements de composition des gaz qui la
remplissent) leur densité à celle de l'eau
où il nagent afin d'obtenir une flottabilité
neutre, celle qui minimise les besoins
énergétiques. Les poissons moins denses que
l'eau de mer, ont souvent une vessie natatoire mais certains
en sont dépourvus et doivent nager continuellement.
Certains poissons comme les torpilles ou les raies, qui
vivent sur le fond ont naturellement une flottabilité
neutre.
|
|
la nage de la couleuvre
|
l'ondulation de l'ensemble du corps assure la propulsion,
la tête reste hors de l'eau au niveau des narines et
des yeux
Remarque: il existe d'autres reptiles excellents
nageurs comme les tortues et les crocodiles...
|
|
voler ...
|
|
Le vol est utilisé par près
d'un million d'espèces dont les 3/4 sont des
insectes.
L'aptitude a voler s'est au moins développé
dans 3 classes: Insectes, Reptiles (Ptérosaures
et Archéoptéryx),
Oiseaux et Mammifères (Chauve-souris). L'on ne parle
pas des poissons-volants dont ce n'est pas le mode de
locomotion habituel, ni des écureuils volants et
autres mammifères planeurs.
La masse musculaire pectorale responsable des mouvements des
ailes représente 15% de la masse totale de l'oiseau
(près de 30% pour les colibri qui ont un vol
vibré stationnaire nécessitant une puissance
mueculaire encore supérieure). A l'inverse de la
course, le milieu fluide ne restituant pas d'énergie
car il est facilement déformable, le vol
nécessite un plus grand travail que la course.
Les muscles du vol des insectes les plus petits (comme les
abeilles, mouches et moustiques) sont très
particuliers du fait qu'ils sont asynchrones : ils
présentent plusieurs contractions pour une seule
stimulation nerveuse et sont reliés non pas
directement aux ailes mais aux éléments
articulés et élastiques du thorax sur lesquels
s'insérent les ailes, ce qui leur permet d'atteindre
des fréquences de 100 à plus de 1000
battements par seconde.
|
voir quelques idées pratiques dans le défisciences
sur l'air
|
|
le vol de la mouette
|
|
|
|
le vol du faucon
|
|
|
|
le vol de l'abeille
|
|
|
|
records
Les animaux champions, la vie
secrète des bêtes, Hachette, 1993
Architecture animale, Karl Von Frisch, Albin Michel,
1975
|
|
vol
|
le plus rapide est un oiseau
le faucon pélerin en piqué peut atteindre
320 km/h mais un miqué ordinaire se fait à 250
km/h et un vol de reconnaissance à 50 km/h
|
le martinet atteint 100 km/h, l'étourneau vole
habituellement à 82 km/h, les oies à 90 km/h
et les canards colvert déjà 75 km/h;
l'aigle royal atteint 160 km/h en piqué
|
un poisson volant sort de l'eau à près de
70 km/h
certains insectes comme le papillon nommé sphinx,
peuvent dépasser 50 km/h et certaines grandes
libellules atteignent 35 km/h
|
|
nage
|
un espadon en fuite a été
chronométré à une vitesse de 130 km/h,
le thon, bon voilier (on nomme ainsi les nageurs longue
distance), peut nager à 70 km/h sur de très
longues distances
|
l'orque est le mammifère marin le plus rapide et
atteint 64 km/h sur de faibles distances
les dauphins ne restent souvent qye moins de 10 min sous la
surface, et ne nagent qu'à 37 km/h au maximum, mais
ils spont capables de faire des bonds de plusieurs
mètres de hauteur au-dessus de la surface.
|
les grandes tortues marines peuvent déjà
nager à 32 km/h et rester 4 heures sous la surface
sans respirer
|
|
course
|
le guépard atteint 112 km/h ce qui lui permet
d'attrapper toutes les gazelles et autres ongulés qui
ne dépassent pas le 80 km/h sauf l'antilope de
Mongolie qui atteint 90 km/h mais surtout qui est capable de
maintenir une vitesse de 64 km/h pendant 20 min, ce dont le
guépard est incapable
|
|
les tortues parcourent moins d'un kilomètre en une
heure mais leut lenteur est battue par le paresseux qui peut
parcourir, au sol, même en "fuite" moins de 100 m
à l'heure. Normalement les paresseux restent dans les
arbres et si, par mégarde, ils tombent, ils peuvent
mettre plusieurs jours à remonter. Leur vitesse
moyenne de dépacement dans les arbres est de 80 m
à l'heure
|
|
saut
|
|
|
|
|
fouissement
|
une famille de blaireau a creusé plus de 310 m de
galeries sur un demi hectare à une profondeur qui ne
dépasse jamais les 2 m.
|
si les termites sont connus pour les édifices de
certains d'entre qui peuvent atteindre plus de 7 m de haut,
on connaît aussi des colonies qui avaint
creuisé une galerie (aux parois consolidées)
dans le sable meuble du désert jusqu'à 40 m de
profondeur afin de couvrir leurs besoins en eau.
|
certains oiseaux comme les guêpiers nichent dans
les falaises le long des cours d'eau et sont capables, comme
le guêpierde Nubie de creuser avec leur becs un
terrier de plus d'1 m de profondeur, dans les paroiis
sableuses des rives.
|
3. formulations par cycle
N.B.
Je suis résolument pour une approche didactique globale du
travail du vivant pour laquelle il n'existe pas de concept
simplifié comme voudraient leur faire croire les concepteurs
des modules insight américains repris par les adeptes de "La
main à la pâte". A chaque enseignant (ou plutôt
chaque équipe pédagogique car c'est un choix qui doit
être cohérent pour l'ensemble de la progression d'un
élève à l'école primaire) de
décider quand et comment il parlera du mouvement animal et du
travail du vivant (et d'une façon plus générale
comment l'équipe souhaite présenter la
vie).
|
concepts
|
cycle 1
|
cycle 2
|
cycle 3
|
|
mouvement et travail du vivant
|
le mouvement peut souvent s'observer chez les êtres
vivants. Il cesse à la mort. Il n'y pas que les
êtres vivants à pouvoir bouger et l'homme peut
construire des machines qui ont un mouvement.
Comme le mouvement n'est pas un travail qui appartient
uniquement à l'être vivant (il ne lui est pas
spécifique), il peut aussi bien être
réalisé pour se nourrir (travail de
nutrition), pour communiquer ou se défendre (travail
de relation) ou pour se re reproduire (travail de
reproduction).
|
|
mouvement dans les différents règnes
|
Tous les êtres vivants ne se déplacent pas
à la même vitesse et de la même
manière. Les bactéries sont souvent
très mobiles comme la plupart des unicellulaires,
malgré leur petite taille. Les champignons et les
plantes ne se déplacent pratiquement et ont sourtout
des mouvements de croissance. Les animaux se
déplacent souvent. On appelle locomotion le
déplacement d'un être vivant; ce terme est
surtout employé pour les animaux.
|
Les bactéries se déplacent habituellement
par des flagelles.
Les unicellulaires peuvent avoir des cils ou des flagelles
ou encore se déformer et ramper avec des
prolongements souvent très fins appellés
pseudopodes.
Les champignons et les plantes ont surtout des mouvements de
croissance lente mais que l'on peut filmer au ralenti et
projeter en accéléré.
Les animaux se déplacent souvent vite et peuvent,
selon leur milieu, marcher, courir, sauter, ramper, fouir,
nager et voler.
|
|
locomotion humaine
|
A la naissance l'enfant n'est pas capable de faire des
mouvements précis et coordonnés, ce qu'il
devient capable de faire au cours des premières
années de sa vie. Habituellement, il sait marcher
entre la fin de sa première année et le milieu
de sa deuxième année. Il peut courir
dès deux ans et peut apprendre à sauter plus
ou moins haut ensuite.
S'il n'a pas peur de l'eau, même peu de mois
après la naissance, il flotte souvent naturellement
et, s'il apprend à agiter ses bras et ses jambes en
cadence, il peut nager efficacement dès 4 ans
environ.
Mais il est très lourd et ne pourra jamais voler sans
l'aide d'un appareil comme le deltaplane.
|
Pour te déplacer tu utilises non seulement tes
membres mais tous tes organes des sens qui te permettent de
te repérer dans l'espace.
Le corps est soutenu par un squelette composé d'os
articulés entre eux. Les mouvements du corps sont
rendus possibles par les mouvements du squelette.
A l'intérieur du corps de nombreux autres organes
participent au mouvement. Par exemple, tu as besoin
d'énergie que tu trouves dans l'alimentation et d'air
que tu inspires. Lors d'une course tu t'essoufles plus vite
que lors de la marche parce que tu dois respirer plus d'air
lors d'un effort plus important.
|
Le mouvement nécessite :
* des muscles
* des os articulés
* un système de commande et d'intégration qui
est le système nerveux.
Le mouvement nécessite le travail de nombreux
autres organes qui participent à la nutrition des
organes du mouvement : poumons, appareil digestif et
appareil circulatoire principalement.
Selon l'intensité et le type de mouvement certains
muscles sont plus sollicités que d'autres.
L'organisme adapte sa nutrition (circulation, rytme
cardiaque, rythme respiratoire) à l'effort
demandé. L'alimentation du sportif doit être
adaptée au type d'effort qu'il doit fournir.
|
|
Pour marcher, courir ou sauter, tu utilises surtout tes
jambes. Pour nager tu utilises aussi beaucoup les bras.
Mais en fait tout ton corps participe au mouvement: la
tête, le tronc, les membres (bras et jambes).
|
|
locomotion animale
|
Les animaux qui vivent dans l'eau savent aussi nager.
Ceux qui vivent dans l'air savent parfois voler. Ceux qui
vivent sur terre savent marcher, courir, sauter ou ramper.
Certains s'enfouissent en creusant des galeries dans la
terre.
|
La locomotion intervient dans d'innombrables exemples
de travail de relation (fuite, défense,
communication...), de nutrition (recherche de nourriture:
chasse, affût, broutage....) ou de reproduction
(recherche du partenaire, parades nuptiales, soin aux
jeunes....).
La locomotion est une des composantes de l'adaptation
de l'animal à son milieu et nécessite une
utilisation coordonnée de très nombreuses
fonctions.
|
|
La nage des poissons est une ondulation de tout le corps
mais aussi un battement des nageoires.
La marche, la course et le saut des animaux terrestres se
fait grâce aux pattes articulées.
Le vol des oiseaux est possible grâce à leurs
ailes. Leurs pattes leur permettent de s'accrocher aux
branches des arbres ou de se déplacer à
terre.
|
Comme pour l'homme on cherchera à montrer la
profonde unité du corps vivant qui, à partir
d'une certaine taille, utilise toujours pour la locomotion
:
* des muscles,
* un squelette (qui peut être souple et liquide comme
le squelette d'un vers composé de poches remplies de
liquide; ou assez souple et externe comme la cuticule des
insectes ; ou encore rigide, externe comme les coquilles des
mollusques ou enfin interne comme les os),
* un système de perception et de contrôle qui
est le système nerveux.
Tous les organes intervenant dans le mouvement sont
nourris grâce aux appareils respiratoires et
circulatoires.
|
4. observations par cycle
|
|
cycle 1
|
cycle 2
|
cycle 3
|
|
|
voir activités proposées sur la page
"je peux bouger" à
réaliser bien sûr en relation avec l'EPS
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|