Cette page présente un extrait de l'ouvrage - aux erreurs
de copie près -
Histoire de la notion de
vie, André Pichot, Gallimard, 1993,
pp 937-954
LA NOTION DE VIE
AUJOURD'HUI
Repères personnels non intégrés au
texte ajoutés (voir version
personnalisée de cette page)
À la fin de ce périple, que peut-on dire de la
notion de vie dans la biologie moderne ? Apparemment. elle est
ignorée. Aujourd'hui, plus que jamais, la biologie suit la
recommandation de Cl. Bernard (citation 8 page 697) : la biologie
doit être une science expérimentale et n'a donc pas
à donner une définition de la vie; ce serait là
une définition a priori et "la méthode qui consiste
à définir et à tout déduire d'une
définition peut convenir aux sciences de l'esprit, mais elle
est contraire à l'esprit même des sciences
expérimentales" ; en conséquence "il suffit que l'on
s'entende sur le mot vie, pour l'employer" et "il est illusoire et
chimérique, contraire à l'esprit même de la
science d'en chercher une définition absolue".
Suivant cette prescription, la biologie moderne ignore donc la notion de vie et se contente de l'analyse d'objets que le sens commun lui désigne comme vivants, analyse montrant qu'ils possèdent un certain nombre de caractères physico-chimiques identiques. La définition de la vie - si elle est parfois évoquée - est reportée à l'infini, comme but et fin ultimes de la biologie. Cette méthode, exclusivement analytique et expérimentale, a considérablement renforcé l'efficacité et la scientificité du travail du biologiste; elle a cependant amené une "physicalisation" telle que l'on a parfois l'impression que, pour rendre scientifique la biologie, il a fallu nier toute spécificité à son objet.
La biochimie (surtout dans sa forme de "biologie moléculaire") est sans doute le stade ultime de cette méthode analytique et expérimentale; les différentes autres disciplines biologiques, comme la physiologie, tendent à y être réduites. Elle a mis en évidence la parfaite identité de nature de la matière, et des lois qui la régissent, dans les êtres vivants et les objets inanimés. Cette matière présente simplement dans les êtres vivants un certain nombre de molécules (et de réactions inhérentes) qui, tout en suivant les mêmes lois physico-chimiques que les autres, ne se trouvent pas aujourd'hui dans les objets inanimés (c'est le cas notamment des macromolécules telles que les acides nucléiques et les protéines).
L'unité de composition de la matière des êtres vivants est telle qu'on peut admettre que les légères variations biochimiques, soit entre les espèces, soit entre deux individus d'une même espèce, ou encore chez un même individu à différents moments de sa vie, ne sont pas suffisantes pour altérer l'unité du phénomène vivant. Ces caractères physico-chimiques propres aux êtres vivants sont liés les uns aux autres avec une assez grande cohérence. Ils forment un tableau tout à fait vraisemblable d'un "être-vivant-en-général", constitué de tout ce qu'il y a de physico-chimique commun à tous les êtres vivants et à eux seuls (acides nucléiques et protéines, processus enzymatiques associés à ces macromolécules, etc.). Cependant, vouloir limiter la spécificité de l'être vivant à de telles caractéristiques physico-chimiques revient à la nier, puisque c'est la ramener à une différence qualitativement analogue à celle existant entre deux objets inanimés. Il n'y aurait alors qu'une collection d'objets ne différant les uns des autres que par leurs caractères physico-chimiques; on voit mal pourquoi on diviserait cette collection en deux classes, vivant et inanimé, - plutôt qu'en trois ou en quatre, - d'autant plus que les critères selon lesquels se ferait cette répartition ne sont pas précisés de manière absolue et que la raison de leur choix reste inconnue, puisqu'elle ne se fonde que sur le sens commun qui désigne comme vivants des objets qui, à l'analyse, présentent certains caractères physico-chimiques identiques. Ces caractères physico-chimiques propres aux êtres vivants, aussi justes soient-ils, ne sont que des indices utiles mais jamais suffisants pour la définition du vivant relativement à l'inanimé. Ils ne permettent pas à eux seuls de cerner la spécificité de l'être vivant (si elle existe) ; ils ne sont pas à l'origine du choix de l'objet de la biologie, ils ne sont que des tentatives de justification physico-chimique du choix fait par le sens commun qui qualifie tel ou tel objet de vivant.
La biochimie montre qu'il n'y a pas de fantôme pour commander la machine, et que la spécificité de l'être vivant ne peut donc résider dans un tel fantôme. Elle montre l'universalité des lois qui régissent la matière, et que la spécificité de l'être vivant ne peut résider dans des lois physico-chimiques qui lui seraient propres, pas plus que dans une force vitale contrôlant son organisation matérielle. Mais, une fois qu'elle a dit cela, elle s'interdit, par ce seul propos, de parler de la spécificité de l'être vivant. Elle ne peut donc la présenter comme un ensemble de caractères physico-chimiques que de tels caractères différencient un être vivant d'un objet inanimé de manière qualitativement analogue à la différenciation de deux objets inanimés) ; mais elle ne peut la nier que sous forme de "fantôme" ou de lois physico-chimiques spécifiques; sinon elle sort de son domaine de compétence.
Il ne s'agit pas là d'une épistémologie tatillonne, mais d'un problème qui reste au cœur de la biologie, même s'il n'est pas explicitement énoncé. On peut en voir un symptôme dans le recours à la notion d'émergence qui est souvent fait devant la difficulté de concilier l'évidente originalité du vivant et sa stricte observance des lois physico-chimiques. La vie est alors présentée comme une qualité nouvelle apparaissant à partir d'un certain degré de complexité de l'organisation physico-chimique, sans qu'il soit davantage précisé comment se fait cette émergence, en quoi elle est nécessaire (si ce n'est qu'il est difficile de nier la spécificité du vivant), ni même en quoi cette vie émergente (qui n'est pas définie) est qualitativement nouvelle comparativement à ce qui est du seul domaine physico-chimique. Dans ces conditions, il est permis de se demander si cette émergence n'est pas simplement un moyen de se débarrasser de la notion de vie, dont on ne sait que faire dans le travail scientifique proprement dit: l'articulation du physico-chimique et du biologique étant supposée se faire par un saut qualitatif (qui relève plus de la magie que de la dialectique de la nature), elle est naturellement mise hors la science. Après avoir montré qu'il n'y a pas de fantôme dans la machine, les biologistes l'en ont fait émerger; comme si les fantômes émanant d'une organisation physico-chimique étaient plus conformes au matérialisme épistémologique que ceux qui sont censés présider à cette organisation. Bien rarement est posée la question de savoir s'il y a encore une machine quand on en a chassé le fantôme, et si ce n'est pas pour garder la machine que parfois on en fait émerger un ; tant est forte, depuis Galien, la conception machinique en biologie.
Une autre attitude, tout aussi symptomatique de la difficulté de concilier l'originalité de l'être vivant et son respect des lois physico-. chimiques, consiste à distinguer divers niveaux de pertinence. Cependant, le fait que la biochimie n'ait pas à se préoccuper de la notion de vie dans son travail, qui est du domaine physico-chimique (niveau moléculaire), mais qu'en revanche cette notion de vie pourrait avoir un sens à un autre niveau (le niveau cellulaire, par exemple) ne fait que déplacer la question: comment articuler la biochimie à cet autre niveau de sorte que d'insignifiante la notion de vie devienne signifiante? Doit-on invoquer ici encore une émergence ?
Le problème de la spécificité de l'être vivant n'est donc pas encore réglé par la biologie moderne. Il est seulement occulté; soit qu'on nie illicitement cette spécificité (ou, ce qui revient au même, qu'on l'assimile à la possession de certains caractères physico-chimiques), soit qu'on la fasse émerger, soit qu'on admette plusieurs niveaux de pertinence. La question reste ouverte: y a-t-il une spécificité de l'être vivant qui ne soit pas un caractère physico-chimique, ni cependant une force vitale plus ou moins surnaturelle?
Si l'on excepte le cas où est admise une notion d'émergence (qui n'apporte pas grand-chose), la biochimie répond par la négative à cette question. Une telle négation ressortit à une conception où l'on n'accepte aucune discontinuité entre vivant et inanimé pour conserver un univers cohérent et unifié. On y admet donc une gradation progressive entre l'inanimé et le vivant, tant dans les formes actuelles (les virus, censés être à la limite de l'inanimé et du vivant) que dans l'apparition de la vie sur terre (cette apparition y est comprise comme une phase prébiotique sans discontinuité marquée). Cette négation de la spécificité du vivant se veut matérialiste; en fait elle confond matérialisme épistémologique et science de la matière.
Aujourd'hui, on a l'impression que ce que vise la biologie n'est pas tant l'étude de la vie (ou des êtres vivants en ce qu'ils ont de spécifique relativement aux objets inanimés) que sa pure et simple négation, le nivellement et l'unification de l'univers par la physico-chimie. Comme si, pour unifier, il valait mieux nier les solutions de continuité que les comprendre. Qu'on ne se méprenne pas; nous ne méconnaissons pas l'intérêt de la biochimie; ce que nous critiquons, c'est cette singulière perversion de la biologie qui consiste à lui donner pour fin la négation de son objet et, par conséquent, d'elle-même en tant que science autonome. Un réductionnisme suicidaire qui n'est pas tant imposé par les résultats de la biochimie que par la médiocre "philosophie spontanée" de maints biochimistes.
Une telle situation est difficilement compréhensible si l'on se réfère à ce que nous avons présenté dans les différents chapitres de cet ouvrage, et notamment dans celui consacré à Lamarck. C'est en effet chez cet auteur qu'on trouve, pour la première fois, une conception de la vie qui reconnaît son originalité comparativement à l'inanimé sans pour autant la faire déroger aux lois de la physique. On aurait pu s'attendre à ce qu'une biologie, se voulant scientifique, expérimentale et matérialiste, s'attache à développer cette conception, qui avait tout naturellement sa place dans son projet. L'histoire (?) ne l'a pas voulu ainsi, pour des raisons diverses (scientifiques: le traitement du temps y était trop complexe pour l'époque; idéologiques : Lamarck était trop lié aux Lumières et à la Révolution pour plaire au XIXe siècle; assez troubles: le comportement de Darwin et des darwiniens à l'égard de Lamarck ne fait honneur ni à leur intelligence ni à leur honnêteté intellectuelle). La biologie, depuis un siècle, s'est faite amnésique; elle a voulu oublier son histoire; à l'en croire, rien n'aurait existé avant CI. Bernard et Darwin; et encore, car même les thèses de ceux-ci, et celles qui ont immédiatement suivi, ont été fâcheusement réécrites.
Les raisons en sont assez claires: la biologie moderne s'est voulue la descendante de l'animal-machine cartésien, faussement compris comme une physiologie mécaniste. La théorie de l 'animal machine, qu'on la décline en termes de machine hydraulique ou de machine chimique, est une théorie incohérente, faite d'un mélange de galénisme et de mécanisme (chez Descartes, elle n'intervenait qu'à titre de physiologie provisoire). Cette incohérence est flagrante chez CI. Bernard; sa conception sera réécrite de manière à n'éclairer que l'aspect mécaniste de l'animal-machine, en laissant dans l'ombre l'aspect galénique. L'incohérence est moins évidente chez Darwin, du fait de la "mollesse" de sa thèse; ce sera cette mollesse même qui sera utilisée pour absorber l'aspect galénique de l'animal-machine (l'origine de la machine expliquée par la sélection naturelle, et le germe vitaliste de CI. Bernard remplacé par un génome).
Si elle peut être réécrite, l'histoire ne peut être rejouée. Aussi n'insisterons-nous pas trop sur cette question, pour nous intéresser plutôt à ce qu'il serait possible de dire de la notion de vie à partir des résultats de la biologie moderne, en les replaçant dans la perspective d'une biologie qui reconnaîtrait la spécificité de l'être vivant tout en s'articulant véritablement avec la physique (contrairement à ce que fait la thèse de l'animal-machine). Après tout, une bonne partie des résultats de la biologie du XVIIIe siècle, obtenus dans des théories préformationnistes ou vitalistes, ont été conservés, il a suffi de les réinterpréter. Il est sans doute possible de faire de même avec les résultats de la biologie actuelle, en oubliant cette thèse absurde de l'animal-machine.
Le principe est de reprendre l'idée lamarckienne de l'orientation du jeu des lois physiques dans des voies déterminées, étant donné que ce jeu des lois physiques est nécessairement temporel. Cette "orientation" permet d'expliquer comment l'être vivant se définit en une entité distincte, se différenciant de son milieu tout en entretenant avec lui de nombreux échanges. Soit une notion de relation-séparation entre l'être vivant et son milieu extérieur (séparation car il est distinct de ce milieu, relation car il entretient avec lui de nombreux échanges, - c'est le problème de la définition d'un système ouvert).
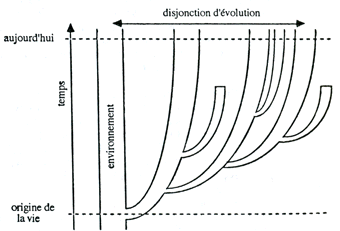
Cette séparation-relation résulte d'une disjonction de leurs évolutions. En effet, l'être vivant, dans son évolution, individuelle ou spécifique, "canalise" le jeu des lois physico-chimiques dans certaines voies aux dépens des autres possibles, alors que l'environnement, lui, évolue en suivant toutes les voies possibles selon les proportions voulues par le libre jeu de ces lois et des équilibres qu'elles régissent.
Cette "canalisation" du jeu des lois physico-chimiques serait explicable en reprenant la thèse néo-darwinienne de la manière suivante. Dans le développement individuel, un programme génétique commande la production d'enzymes qui catalysent certaines réactions aux dépens des autres possibles à partir des mêmes substrats. Ces réactions catalysées (et leurs produits) acquièrent ainsi une importance qu'autrement elles n'auraient jamais eue. Dans l'évolution des espèces, certains micro-événements (mutations) sont sélectionnés et leurs effets amplifiés au sein de la population en fonction d'un critère qu'on peut résumer par "meilleure aptitude à vivre". Ces micro-événements acquièrent ainsi une importance que leur seule valeur physico-chimique n'aurait pu leur donner. Dans les deux cas (dont l'un prolonge l'autre grâce à une engrammation dans le génome), la canalisation du jeu des lois se fait par la sélection et l'amplification d'une des possibles voies d'évolution, alors que l'environnement se répartit dans toutes ces voies possibles selon les proportions voulues par les principes des équilibres physico-chimiques (1).
Quoi qu'on pense de l'explication néo-darwinienne (et de l'utilisation qu'elle fait de la théorie de l'information), il n'en est pas moins incontestable et vrai que l'évolution (individuelle et spécifique) des êtres vivants ne suit pas simplement celle de l'environnement. Elle en est autonome, car elle a ses principes propres (ceux de cette "canalisation''). Elle n'en est cependant pas totalement indépendante, car les êtres vivants, tout en évoluant de manière disjointe, ont de constantes relations avec leur milieu (de nombreux échanges de matière et d'énergie). D'un point de vue thermodynamique, ce sont d'ailleurs ces. relations qui permettent aux êtres vivants d'évoluer de manière disjointe, et donc de se distinguer de l'environnement. Sont ainsi conciliés les deux moments de l'(auto)définition des êtres vivants: leur distinction d'avec l'environnement (évolution individuelle, et spécifique, disjointe) et leur relation avec lui (échanges divers). Pour le moment, retenons donc cette disjonction d'évolution (individuelle et spécifique), en laissant de côté l'explication qu'en pourrait donner le néo-darwinisme.
La disjonction d'évolution (individuelle et spécifique) a deux conséquences principales: le caractère temporel de la définition de l'être vivant et l'écart physico-chimique entre lui et son milieu.
Voyons d'abord la première, le caractère temporel de la définition de l'être vivant. C'est seulement dans le mouvement de son évolution individuelle, et donc dans le temps, que l'être vivant est défini comme une entité distincte. En effet, puisqu'il est ouvert sur son milieu, c'est seulement dans ce mouvement qu'est rompue la continuité physico-chimique entre eux, et c'est donc seulement dans le temps de cette évolution individuelle disjointe qu'il est défini (en tant qu'entité distincte). Cette évolution individuelle, par son mouvement, rassemble les parties de l'être en un tout qui est ainsi distingué du milieu. Il ne faut pas inverser la proposition en disant que l'être évolue de manière disjointe parce qu'il est distinct de son milieu; c'est, au contraire, en évoluant de manière disjointe qu'il se distingue de ce milieu. Il faut comprendre l'individu à partir de l'individuation, et non pas l'individuation à partir de l'individu (2).
Pour mieux le saisir, on peut comparer cette définition de l'être vivant par son évolution disjointe à la définition d'un groupe de personnes se déplaçant ensemble. Sur une place publique, de nombreuses personnes marchent en tous sens; si certaines d'entre elles se déplacent ensemble, elles vont se distinguer des autres comme constituant une entité, un groupe. D'autres personnes peuvent se joindre à ce groupe et certaines le quitter. Il n'est pas nécessaire que ces personnes regroupées soient toujours dans le même rapport (la même position relative, comme des soldats qui marchent au pas), mais leurs mouvements les unes par rapport aux autres doivent être limités, sinon elles sortent du groupe. Il n'est pas nécessaire non plus qu'elles soient reliées entre elles (en se donnant la main, en entourant leur groupe par une corde qui le délimite, etc.). Enfin, le fait qu'elles se meuvent (et la manière dont elles le font) peut donner à leur groupe une certaine structure et une certaine forme; cette structure peut d'ailleurs se modifier au cours du temps sans que cela altère le caractère de groupe (il y a un remaniement interne au groupe, mais celui-ci reste un groupe). Ce qu'il faut bien saisir ici, c'est que le regroupement de ces personnes est inhérent à leur mouvement commun qui les distingue des autres personnes qui ont des déplacements non coordonnés, ni entre eux ni avec le leur.
Le fait que les personnes ainsi regroupées puissent constituer une structure particulière ne suffit pas à définir leur groupe comme une entité distincte. On pourrait certes évaluer la probabilité thermodynamique de cette structure, et ainsi la distinguer du "désordre" des mouvements non coordonnés des autres occupants de la place; mais un renseignement de cette nature n'a pas grand intérêt ici. Ainsi. si l'observateur fait des photographies aériennes de cette place, il ne pourra pas distinguer une structure "morte" et une structure "vivante" (quelle que soit sa probabilité thermodynamique). Seule la dimension temporelle l'indique: les éléments constituant une structure "vivante" poursuivent leur mouvement commun, tandis que ceux constituant une structure "morte" vont se disperser en adoptant des mouvements individuels non coordonnés (ainsi le cadavre qui se décompose). Cet ensemble d'éléments n'est donc défini comme un groupe que dans le temps et non de manière atemporelle (cf. l'expression bergsonienne selon laquelle "la forme de l'être vivant est le contour d'un mouvement").
On comparera un tel groupe, défini par son mouvement, à l'être vivant défini comme entité par son évolution individuelle disjointe; il faut alors remplacer le simple déplacement dans l'espace par une transformation physico-chimique. Le problème est évidemment de comprendre comment sont coordonnés les "mouvements physico-chimiques" des éléments formant ce groupe. Chacun d'entre eux a-t-il reçu dès le départ un ordre de marche articulé avec celui des autres en un plan d'ensemble prédéfini (un programme) ?
Est-ce qu'il existe des forces qui lient les éléments, des contraintes qui coordonnent leurs mouvements, etc. ? Sans doute tout cela à la fois, mais dans des proportions difficiles à préciser; quelques éléments sont vraisemblablement déterminants, et entraînent les autres en fonction de diverses contraintes. Nous ne développerons pas cette question ici ; nous voulions seulement expliciter par cette comparaison la primauté du caractère dynamique de l'être vivant.
Quoi qu'il en soit du détail de ce processus totalisateur, il ne faut donc pas considérer l'être vivant comme une entité prédéfinie qui doit rester en vie (?) en maintenant sa structure et sa composition constantes grâce à des régulations. On doit abandonner ce paradigme cybernétique de la physiologie bernardienne et le remplacer par une conception dynamique. Au lieu de maintenir un certain nombre de constantes définies de manière absolue (à l'image de la constance du milieu intérieur de CI. Bernard), il s'agit pour l'être vivant de maintenir son évolution (individuelle, au moins) disjointe de celle de l'environnement. La vie est alors caractérisée par cette évolution disjointe. On garde bien la conception classique qui veut que l'être vivant se caractérise par sa persévérance dans son être; mais cette persévérance de l'être vivant dans son être est comprise comme le maintien d'une disjonction d'évolution (et donc le maintien de la distinction d'avec le milieu), et non comme le maintien de certaines constantes (d'ailleurs difficiles à préciser, et même assez imaginaires car la plupart des paramètres habituellement retenus varient selon des rythmes plus ou moins réguliers (3). L'être reste en vie tant qu'il maintient son évolution individuelle disjointe de celle de l'environnement, quelles que soient les variations qualitatives et quantitatives de sa matière et de sa structure. Mourir, pour l'être, c'est rejoindre son environnement et évoluer avec lui (c'est-à-dire cesser de s'en différencier). Il s'agit ici de "re-dynamiser" l'être vivant, de réintroduire en lui la dimension temporelle et dynamique que le statisme bernardien avait éliminée.
Outre la temporalité de la définition de l'être vivant, la conséquence la plus remarquable de la disjonction d'évolution est l'"écart" qui en résulte entre l'être et son milieu: si la première entité vivante au temps zéro de sa vie était en parfaite continuité physico-chimique avec le milieu prébiotique (puisqu'elle y est apparue spontanément, par le seul libre jeu des lois physiques), les êtres vivants d'aujourd'hui ne sont plus dans une telle continuité avec leur environnement (la génération spontanée de ces êtres n'y est pas possible). En effet, à partir de la continuité primitive, êtres vivants et environnement, malgré leurs relations (et grâce à elles), ont évolué de manières disjointes, asynchrones. Ce déphasage entre leurs évolutions a entraîné l'installation entre eux d'une discontinuité qui croît avec le temps.
Cette discontinuité physico-chimique se traduit par le fait que l'être vivant actuel, lorsqu'on le considère dans sa globalité, n'est pas directement reliable à son milieu selon les lois physico-chimiques. Il faut remonter à l'origine de la vie pour faire ainsi se rejoindre l'être et son milieu; d'où l'importance, en biologie, de l'explication historique que constitue l'évolution des espèces. Chacune des étapes, dans l'évolution des espèces comme dans l'évolution de l'environnement, respecte parfaitement les lois physiques, et ceci depuis l'origine de la vie où la première entité était en continuité avec son environnement; mais, comme ces évolutions ont divergé sitôt achevée la phase prébiotique, la discontinuité s'est tout de suite installée entre eux. On a donc deux histoires divergentes autonomes; à l'intérieur de chacune d'elles les lois physico-chimiques sont respectées, mais en déphasage, si bien que ces histoires (et leurs produits, êtres vivants et environnement) ne sont pas directement reliables selon ces lois. La nécessité d'une théorie de l'évolution en biologie (pour compléter l'explication biochimique des êtres vivants) tient à cette non-relation-directement-selon-Ies-Iois-physico-chimiques de l'être actuel et de son environnement actuel (et donc à l'impossibilité d'expliquer la présence de celui-là dans celui-ci par ces seules lois), bien plus qu'à l'existence de preuves paléontologiques ou autres (ici encore l'analyse de Lamarck est bien supérieure à celle des biologistes modernes qui, comme Darwin, n'ont jamais véritablement compris la nécessité de l'évolution).
Cette "non-relation directement selon les lois physico-chimiques" n'a rien de mystérieux; ce n'est qu'une autre manière de nommer la disjonction des évolutions. Et ceci pour deux raisons. Tout d'abord, comme on vient de le voir, le fait qu'ils soient produits par des évolutions disjointes, implique que l'être et l'environnement actuels sont en discontinuité. D'autre part, dire que l'être actuel évolue de manière disjointe de son environnement actuel, revient à dire que son évolution actuelle n'est pas directement reliée à celle de son environnement selon les lois physico-chimiques. En effet, puisque ces lois sont, pour la plupart, des lois d'évolution du type "si A au temps t1. alors B au temps t2 ", la disjonction des évolutions n'est rien d'autre que l'impossibilité de relier cet être et son environnement par de telles lois temporelles. Par conséquent, qu'on le considère "instantanément" (en tant qu'être actuel) ou dans son évolution (individuelle et spécifique), l'être vivant n'est pas directement reliable à son environnement selon les lois physico-chimiques ; il Y a une discontinuité, un écart, entre eux (4).
Cette "non-relation directement selon les lois physico-chimiques" se manifeste par le caractère physico-chimiquement improbable de la structure et de l'occurrence de l'être vivant dans son environnement La biologie moléculaire essaye de l'expliquer en recourant à la notion d'information (qu'on peut relier à celle d'improbabilité) : plus l'être est évolué, plus son écart avec l'environnement est marqué, plus il y est donc physico-chimiquement improbable, et plus il devra posséder une grande quantité d'information génétique pour que son occurrence et sa structure y soient explicables (que cela se traduise ou non par une plus grande quantité d'ADN, car ce qui compte ici est la valeur informationnelle de celui-ci). L'information génétique pallie en quelque sorte l'impossibilité d'expliquer l'être vivant actuel dans son milieu actuel par les seules lois physicochimiques. Cette information, en ce qu'elle est une sorte de "mémoire" de l'espèce (une réification de l'histoire de cette espèce sous la forme physique du génome), sert ainsi à articuler (plus ou moins bien) l'explication historique et l'explication physico-chimique de l'être vivant (5).
Que l'être vivant considéré globalement ne soit pas directement reliable à son milieu selon les lois physico-chimiques, n'est pas incompatible avec le fait qu'il entretienne avec son milieu des relations dont chacune, considérée isolément des autres, respecte parfaitement ces lois, Ce qui s'explique ainsi.
La discontinuité physico-chimique entre l'être actuel et son environnement actuel constitue une sorte de séparation synchronique entre eux, mais elle ne se comprend qu'en tant que moment artificiellement isolé de la disjonction d'évolution individuelle, sans laquelle elle ne peut se maintenir. À elle seule, cette discontinuité ne suffit pas à définir l'être en tant qu'entité distincte puisque, laissée à elle-même, elle tend à s'effacer, en raison de la tendance spontanée au retour à l'équilibre des systèmes physico-chimiques. Il faut qu'elle soit sans cesse entretenue par une évolution (individuelle) qui contrarie cette tendance au retour à l'équilibre. Ainsi, un cadavre "frais" n'a pas une discontinuité physico-chimique avec son environnement beaucoup moins marquée que celle d'un être vivant de même espèce; mais cette discontinuité va en s'atténuant au fur et à mesure de la décomposition. L'évolution du cadavre n'est pas disjointe de celle de l'environnement, elle suit le seul jeu libre des lois physico-chimiques. La discontinuité qui reste entre eux n'a alors aucun intérêt puisqu'elle va évoluer, elle aussi, selon ce seul libre jeu, et disparaître.
La discontinuité physico-chimique entre l'être vivant et l'environnement n'a donc d'intérêt que dans son devenir, dans sa dynamique, et non lorsqu'on la considère de manière instantanée ou atemporelle (tout comme la structure de cet être n'a d'intérêt que dans son devenir, voir ci-dessus la comparaison au groupe de personnes qui marchent). Elle ne nous importe que dans la mesure où elle distingue l'être de son environnement, et cette distinction existe seulement dans le temps.
Pour que se maintienne une telle discontinuité, il faut que l'évolution du rapport global de l'être et de l'environnement ne se fasse pas selon le libre jeu des lois physico-chimiques, au contraire de ce qui se passe dans le cas d'un objet inanimé ou d'un cadavre. Cela ne signifie pas qu'alors cette évolution ne respecte pas ces lois, mais que, parmi l'ensemble des relations physico-chimiques locales possibles entre l'être et son environnement, seules quelques-unes (sélectionnées) se réalisent: celles qui, organisées entre elles (et souvent possibles seulement l'une par l'autre au sein de cette organisation (6), s'accordent avec la poursuite de la disjonction d'évolution. Un objet inanimé, ou un cadavre, aurait, lui, toutes les relations qu'il lui est possible d'avoir avec son environnement. Une membrane à perméabilité variable est une manière simple de sélectionner ces relations, puisqu'elle permet ou ne permet pas à telle ou telle substance d'entrer ou de sortir, à tel ou tel moment.
Ce processus est donc une sorte de "canalisation" du jeu des lois physico-chimiques dans le rapport global entre l'être et son environnement ; et cette "canalisation" est indissociable de celle qui se produit lors de l'évolution de la structure de cet être (au niveau individuel, mais aussi, indirectement, spécifique). Tout comme cette structure est constituée par un ensemble organisé de réactions physico-chimiques sélectionnées parmi les possibles, ce rapport global est constitué par un ensemble de rapports physico-chimiques locaux sélectionnés parmi les possibles et organisés entre eux. Ces deux "canalisations" n'existent que l'une par l'autre et sont les deux faces d'un même processus résolvant le déséquilibre né de la disjonction d'évolution: interne pour la structuration de l'être, externe pour la structuration de ses relations avec le milieu.
Ce jeu canalisé des lois physico-chimiques dans l'évolution du rapport global de l'être et de son environnement constitue le comportement. C'est une définition large puisqu'elle englobe toutes leurs interactions physico-chimiques, qu'elles soient ou non différenciées en une sensibilité, une motricité, une absorption, une excrétion, etc. En ce sens, on peut parler de comportement pour tout être vivant, y compris les végétaux.
Une forme minimale de comportement consiste, pour l'être, à ne prendre en compte dans l'environnement qu'un certain nombre d'éléments. C'est là une première manière de préserver la discontinuité qui le sépare de cet environnement, puisque l'indifférence à l'égard d'une partie de celui-ci est une sorte d'isolement partiel (grâce, par exemple, à une membrane à perméabilité sélective ).
Ces éléments externes sélectionnés constituent le milieu extérieur de l'être vivant, c'est-à-dire la partie de l'environnement dont il tient compte. Sont ainsi distingués le milieu extérieur et l'environnement: le milieu extérieur est la partie de l'environnement avec laquelle l'être vivant a des relations, partie qui est alors structurée, pour l'être vivant, en fonction de (et par) ces relations (donc par les capacités d'action de l'être vivant, alors que l'environnement, lui. est structuré par les lois physiques).
Du fait qu'il est le produit d'une évolution disjointe de celle de l'environnement, l'être vivant se trouve dans un état d'''errance physico-chimique" au sein de celui-ci (il ne lui est pas relié directement selon les lois physico-chimiques). Il doit alors organiser ses relations (physico-chimiques) avec cet environnement: sélectionner certains de ses éléments avec lesquels il interagira, et coordonner ses actions et réactions avec ces éléments sélectionnés (bien souvent, les relations que l'être a avec chacun d'entre eux n'est possible que grâce à celles qu'il a avec les autres au même moment; ce qui donne au comportement un aspect de totalité). De la sorte l'être vivant se constitue, au sein de l'environnement, un milieu extérieur structuré auquel il se relie par un comportement structuré. À une errance physico-chimique dans l'environnement (laquelle conduit tôt ou tard à la disparition de l'entité errante) est substituée une action comportementale déterminée au sein d'un milieu extérieur (constitué et structuré de la sorte). Et ce comportement fait partie intégrante de la disjonction d'évolution.
En résumé, la disjonction des évolutions se traduit par une autonomie (croissante) des êtres vivants par rapport à leur environnement Cette autonomie n'est pas une indépendance, mais une "non-relation globale à l'environnement directement selon les lois physico-chimiques" ; à celles-ci sont substituées des lois de transformation interne et de comportement propres à l'être (c'est le sens étymologique du mot autonome: qui a ses lois propres). des lois propres qui doivent néanmoins s'accorder avec les lois physiques.
Entre l'être vivant et son milieu, il n'y a donc ni séparation radicale (nombreux échanges entre eux), ni continuité physico-chimique (disjonction des évolutions) ; et ceci sans que les lois naturelles soient le moins du monde violées, et sans recours à une quelconque force vitale.
On tient là une différence fondamentale entre les êtres vivants et les objets inanimés: les objets inanimés, même ceux qui s'auto-construisent (cristaux, structures dissipatives), sont séparés de leur environnement selon l'espace (ils ont des formes définies) mais reliés à lui selon la physique (ils évoluent avec lui) ; les êtres vivants sont séparés de leur environnement non seulement selon l'espace (ils ont des formes définies) mais aussi selon la physique (leur évolution disjointe a créé un écart, une discontinuité). Reste à relier leur séparation selon l'espace et leur séparation selon la physique, à relier leur forme et la dynamique de leur évolution disjointe.
Nous avons expliqué ci-avant que l'être vivant ne reste en vie que tant qu'il maintient son évolution disjointe de celle de l'environnement. Il s'agissait alors essentiellement de l'évolution individuelle (développement et sénescence). On ne peut cependant négliger ici le rôle de l'évolution des espèces. Celle-ci se traduit par le fait qu'à sa naissance, le nouvel être vivant a un certain écart avec son environnement, écart qui est fonction de l'espèce à laquelle il appartient (et donc de l'histoire de ses ancêtres), mais aussi de l'environnement où il naît (puisque cet environnement est l'autre "pôle" de l'écart). Une fois né, le nouvel être vivant va devoir subsister dans l'environnement avec lequel il est en discontinuité. Pour cela, il doit adopter une évolution individuelle disjointe de cet environnement, car il ne peut pas laisser sa discontinuité évoluer selon sa "pente naturelle" (cette pente est celle de la disparition, et donc de l'indifférenciation au sein de l'environnement). D'une certaine manière, la disjonction de l'évolution individuelle, avec ses deux faces, interne et externe (structurale et comportementale), est la réponse à la discontinuité imposée à l'être vivant, dès sa naissance, par la disjonction de l'évolution des espèces.
Plus l'être est évolué, plus est grand l'écart qui, à sa naissance, le sépare de son environnement Encore faudrait-il s'entendre sur ce qu'est un être plus évolué qu'un autre (voir dans les chapitres VII et IX les considérations de Lamarck et Darwin à ce sujet: complexification, différenciation et spécialisation des organes). Si l'on ne prend en considération que le temps, les êtres actuels sont tous aussi évolués les uns que les autres (la durée d'évolution de leurs espèces, depuis l'origine de la vie, est la même). Une solution simple à ce problème est de considérer que les êtres actuels sont séparés de leur environnement par un écart qui est fonction, non seulement de la durée de leur évolution, mais aussi du degré de disjonction de celle-ci, de sa "pente". L'évolution des espèces est en un effet un processus ramifié, et toutes ses branches n'ont pas une disjonction aussi forte (quoique, d'un point de vue théorique, la "pente" de cette disjonction doive toujours être positive). De sorte que l'écart entre les êtres vivants et leur environnement n'est pas le même pour tous (voir figure ci-dessous).
L'écart entre un être vivant est ainsi fonction de deux paramètres, la "pente" de la disjonction de son évolution, et la durée de cette évolution. À pente égale, plus un être est proche de l'origine de la vie, plus son écart est faible. A durée égale, plus la pente de disjonction d'évolution est forte, plus l'écart est grand. Un être actuel, qui a évolué avec une pente "faible", a un écart qui n'est pas beaucoup plus grand que celui d'un être fossile dont la pente d'évolution a été plus forte. On peut alors dire de l'un comme de l'autre que ce sont des êtres peu évolués, l'un parce qu'il vivait il y a très longtemps, l'autre parce que, bien qu'il vive aujourd'hui, son évolution n'a pas eu une "pente" très forte.
Si l'on tient compte de ce que, pour un écart donné, seul un nombre restreint de formes est sans doute possible, du fait des contraintes inhérentes au statut physique très particulier qui résulte de la disjonction d'évolution, alors il est très probable que des êtres actuels et des êtres fossiles ayant des écarts comparables avec l'environnement auront des formes comparables. De manière plus générale, la physique étant ce qu'elle est. il n'y a sans doute qu'un nombre limité de formes capables d'exister en disjonction avec leur environnement, les formes dans lesquelles l'écart physique est surmonté, résolu (plutôt que comme l'expression d'un programme, la morphogenèse serait à comprendre comme la solution à la tension inhérente à cet écart physique; les voies réactionnelles choisies dans cette morphogenèse seraient les voies "de moindre résistance", plutôt que des voies déterminées par une information génétique (7).
Ces formes ne pouvant être réalisées que par un processus historique (elles dépendent de l'engagement dans telle ou telle voie à tel ou tel moment de l'évolution), il n'est pas sur que toutes les formes possibles aient été réalisées. Ce qui signifie qu'il y a bien un parallélisme entre les formes qui se sont succédées dans le temps et les formes actuelles classées selon un écart croissant, mais que. du fait des contingences de l'histoire, ce parallélisme ne peut être qu'approximatif.
Nous ne développerons pas tout cela ici, car cela nous entraînerait trop loin (8). Il s'agissait seulement, pour conclure cet ouvrage, d'indiquer quelques voies possibles pour une biologie qui étudierait la vie. et non simplement la matière des êtres vivants. Il existe maintenant des outils mathématiques et physiques qui le permettent. Contrairement à ce que pourrait laisser penser la stagnation théorique de la biologie moléculaire (mal masquée par le développement d'applications pratiques et les opérations à grand spectacle du style "génome humain"), l'histoire de la biologie n'est pas finie.
(1) On pourrait reprendre ici l'expression de F. Jacob "le jeu des possibles", mais certainement pas pour aboutir à une théorie du bricolage de l'évolution. comme le fait cet auteur; l'important est plutôt dans la différence que le jeu des possibles installe entre l'évolution (individuelle et spécifique) des êtres vivants et l'évolution de l'environnement. Par ailleurs, dans la vie de l'individu, l'engagement dans une des voies possibles ne constitue jamais un fonctionnement machinique, mais une transformation (et cela tout le long de la vie: dans le développement, l 'âge adulte et la sénescence).
(2) Cf. G. Simondon, L'individu et sa genèse physico-biologique, P.U.F.. Paris 1964.
(3) Néanmoins, Ie maintien de constantes équivaut souvent à une évolution disjointe comparativement à un milieu où les facteurs correspondants sont variables (l'homéostasie opposée à la variabilité des conditions externes). D'autre part, le maintien de la disjonction d'évolution, s'il ne consiste pas en un maintien d'une structure. nécessite sans doute le maintien d'une organisation circulaire (commune à toutes les structures des êtres vivants), où chaque partie n'est physico-chimiquement possible que par les autres, dont chacune n'est possible que de la même manière (voir ce que propose Kant pour les corps organisés naturels dans la Critique du Jugement).
(4) Cet écart se mesure comme l'inverse d'une probabilité. Il suffit de considérer l'évolution (individuelle ou spécifique) comme une succession de n étapes, à chacune desquelles une voie est choisie parmi m voies possibles. En supposons, pour simplifier, que m est le même à chaque étape et qu'il y a une équiprobabilité pour les différentes voies, la probabilité du produit obtenu à la fin du processus évolutif est égale à 1/mn', et son écart à l'environnement est mn".
(5) 0n peut faire les critiques suivantes à l'utilisation de la théorie de l'information en biologie. D'une part, on a recours à une information génétique pour pallier l'impossibilité d'expliquer l'être vivant actuel dans son milieu actuel par le seul libre jeu des lois physico-chimiques. D'autre part, on prétend que ce recours à une information génétique ne fait appel qu'à ces lois (le "programme génétique" et son expression sont décrits en termes purement physico-chimiques). Il y a là une contradiction. La théorie de l'information est utilisée pour ramener l'explication historique à une explication physico-chimique; c'est-à-dire qu'on ramène la dimension historique à une mémoire, laquelle mémoire consiste en une information stockée sous une forme physique. De cette manière la dimension historique (passée) est rendue présente sous une forme physique (baptisée "information génétique'), et c'est cette forme physique que l'on fait intervenir en complément de l'explication physico-chimique actuelle. La notion d'information sert donc à articuler l'explication physico-chimique à l'explication historique qui doit nécessairement la compléter, et à faire passer leur amalgame pour une explication qui ne serait que physico-chimique. - Le darwinisme a débarrassé l'explication historique des contraintes physiques en utilisant la contingence (voir page 845). Mais cette utilisation de la contingence ne vaut que pour la formation des espèces (elle intervient à la faveur des discontinuités que la reproduction introduit entre les générations) ; elle ne vaut pas pour la formation des individus qui est un processus continu où il n'y a pas de "fissures" par où introduire le hasard (même si on a récemment cherché à expliquer le développement individuel, notamment celui du système nerveux, en ayant recours à un principe sélectif - cela en raison des difficultés à appliquer la notion de programme génétique dès qu'on cherche à dépasser le niveau de la simple métaphore informatique). L'histoire individuelle ne pouvant pas être libérée des lois physiques (au contraire de l'histoire spécifique). la biologie doit y canaliser le jeu de ces lois (elle reprend ici la démarche lamarckienne). C'est à cela que sert la théorie de l'information ou, plus exactement, la théorie du programme génétique. - S'il n'y avait que ce problème de "canalisation" du jeu des lois physiques, la solution serait admissible (y compris l'affirmation que cela ne fait intervenir que les lois physiques, car on peut parfaitement imaginer que le jeu de ces lois est orienté par une organisation qui est elle-même purement physique) ; parler d'information plutôt que d'organisation ne serait qu'une métaphore plus ou moins adéquate. Malheureusement, ce problème de "canalisation" rencontre une difficulté: le choix d'un possible aux dépens des autres n'est pas neutre d'un point de vue thermodynamique. ni dans l'évolution des espèces, ni dans le développement individuel. Et là, la théorie de l'information est utilisée de manière physique et non plus simplement métaphorique. On donne en effet à l'ordre du génome (la succession des nucléotides dans l'ADN) une valeur informationnelle en ce qu'il doit rendre compte d'une structure vivante que le seul libre jeu des lois ne peut expliquer. La manière dont cela se fait est très floue; il semble qu'on utilise une dissociation entre l'ordre des nucléotides (un arrangement parmi un très grand nombre d'arrangements possibles) et l'ordre thermodynamique (sauf cas exceptionnels, tous les arrangements de nucléotides d'une longueur donnée ont à peu près la même probabilité thermodynamique, mais tous les arrangements de nucléotides ne correspondent pas indifféremment à une structure vivante ordonnée). On suppose apparemment que, pour aboutir à un ordre cellulaire ou pluricellulaire, cette dissociation entre ordre des nucléotides et ordre thermodynamique est complétée par l'ouverture de l'être vivant, qui puise de l'énergie dans son milieu. C'est du moins ce qu'il semble au premier abord, car je n'ai jamais trouvé une explication convaincante de l'utilisation physique (et non simplement métaphorique) de la théorie de l'information en biologie (utilisation physique pourtant avérée dans un ouvrage comme L'organisation biologique et la théorie de l'information de H. Atlan). - La biologie utilise ainsi la notion d'information à deux niveaux. un niveau métaphorique dans la notion de programme génétique, et un niveau thermodynamique dans l'explication physique de la structure de l'être vivant Mais elle ne précise jamais quand elle passe de l'un à l'autre, et entretient le plus grand flou sur la question (voir aussi la note 7 pages 953-954).
(6) Ce qu'on peut comprendre par une organisation circulaire où chacun des rapports de l'être avec son milieu n'est possible que grâce à l'ensemble des autres, dont chacun n'est possible que de la même manière; exactement comme chaque partie de l'être n'est physico-chimiquement possible que par les autres parties, dont chacune n'est possible que de la même manière (voir note 3 page 945).
(7) Dans cette optique, ce qu'un être hérite de ses parents, ce n'est pas tant un génome qu'un certain écart avec l'environnement, écart qui est fonction à la fois de l'environnement et de ce qu'à sa naissance l'être possède en matière non seulement de génome mais aussi de structure générale. La vie de cet être va alors consister en une évolution disjointe de celle de l'environnement, à partir de cet écart initial. Pour que l'être se maintienne en vie, il faut qu'à tout moment il résolve sa situation physiquement instable, par une modification de sa structure interne et/ou de son comportement (son rapport avec le milieu, ce qui peut modifier ce milieu) ; ces modifications créent une nouvelle situation. d'où un nouvel écart, et ainsi de suite,jusqu'à ce qu'il n'y ait plus de solution possible (compte tenu de la structure de l'être et de l'environnement). La résolution de chacun de ces écarts consiste à s'engager dans une voie d'évolution parmi les possibles; à tout moment, la possibilité d'adopter telle ou telle solution dépend donc des voies empruntées antérieurement. Que le génome intervienne dans ce processus, c'est certain, qu'il soit un programme, certainement pas: selon sa nature, telles ou telles possibilités seront ou non permises à l'être vivant dans telle ou telle situation, telles ou telles voies d'évolution (individuelle. mais aussi spécifique) seront ouvertes ou non. La nature de l'être vivant, son "essence". est alors fonction de son écart par rapport à l'environnement, lequel écart détermine sa forme; elle n'est fonction de son génome que dans la mesure où celui-ci intervient dans cet écart (le génome représente surtout les moyens - et même seulement une "partie" des moyens - dont l'être dispose pour vivre en discontinuité avec l'environnement). C'est l'écart qui définit l'être, et non les moyens dont il dispose pour "résoudre" cet écart. - Cela permet de revenir sur la critique de l'utilisation de la théorie de l'information. En effet, l'information sert, nous l'avons dit, à ramener l'explication historique à une explication physique actuelle: l'histoire est rendue présente par son engrammation dans une sorte de mémoire physique. Or il n'est pas sûr qu'une telle engrammation soit nécessaire. En effet, il ne faut pas confondre histoire et mémoire. Dans la mémoire, le passé intervient en tant qu'il est rendu présent par son engrammation dans une structure physique actuelle (sous forme d'informations). Dans l'histoire, le passé intervient en tant qu'il est passé: telles et telles voies d'évolution ont été suivies parmi toutes celles qui étaient possibles, et le présent en est la conséquence. L'histoire se caractérise par ce choix d'une voie physique parmi les possibles; elle ne consiste pas en une libre application du jeu des lois physiques pendant un certain temps. réalisant tous les possibles contenus dans la situation de départ. Par conséquent, le simple fait que les êtres vivants aient une histoire implique que la situation présente porte la trace du passé, sans qu'il soit besoin d'une mémoire ayant engrammé ce passé. Cette trace est l'écart qui existe entre l'être et son environnement, écart qui est fonction à la fois de l'histoire de l'être (et de ses ancêtres) et de l'histoire de l'environnement (ce qui relativise considérablement toute la problématique de l'inné et de l'acquis développée depuis Weismann: l'écart est inné sans être "génétique"). Il est certain que le génome joue un rôle important dans cet écart, mais il le joue en relation avec le reste de la structure vivante et avec l'environnement.
(8) J'en ai exposé quelques conséquences dans le premier chapitre de ma Petite phénoménologie de la connaissance (Aubier, Paris 1991).