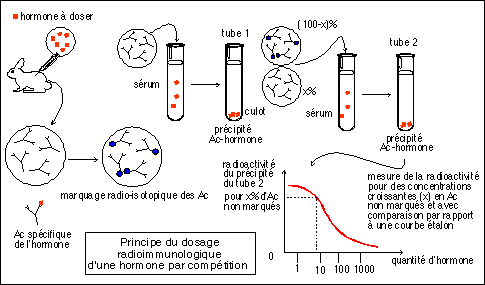
|
(ce tableau, incomplet, vous permet de situer les cellules étudiées en TP, il n'est bien évidemment pas à mémoriser) (RNC = rapport nucléocytoplasmique, qui est un indice de différentiation: une cellule a un RNC d'autant plus élevé qu'elle est peu différenciée) |
||||
|
Si certaines cellules comme des cellules épithéliales ou folliculaires des organes lymphoïdes périphériques (comme les ganglions lymphatiques) participent aux réactions immunitaires, on peut tout de même affirmer que toutes les cellules immunitaires se forment dans la moelle rouge des os longs (on dit qu'elles ont une origine hématopoïétique, l'hématopoïèse étant le processus de formation des cellules sanguines dans la moelle) à partir d'une seule lignée de cellules dites cellules souches. L'hématopoïèse est estimée à environ 100 milliards de globules rouges par jour par exemple. |
||||
|
|
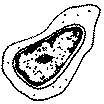 |
5 µm de Ø, RNC élevé, localisées dans le foie pendant la vie fœtale, elles colonisent ensuite la moelle osseuse où elles se divisent activement, ce sont des cellules peu différenciées (pluripotentes : elles ont plusieurs destinées possibles mais toutes inscrites dans la lignée hématopoïétique) |
||
|
|
|
|
||
|
|
|
|
7 µm de Ø, pas de noyau (chez les oiseaux et les mammifères), biconcave, durée de vie environ 120 jours (phagocyté) |
|
|
|
(thrombocyte) |
fragment cytoplasmiques anucléés |
||
|
|
(fait partie des globules blancs: leucocytes) |
 |
10 à 14 µm de Ø, RNC moyen, doué de diapédèse, circule dans le sang |
|
|
|
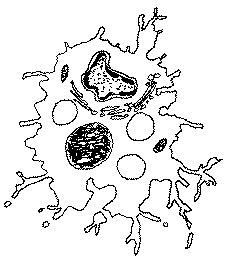 |
Ø >> 20 µm, RNC très faible, très spécialisé (phagocytose) avec de nombreuses vacuoles de phagocytose (phagosomes) des lysosomes, des pseudopodes et un Golgi développé |
||
|
|
Remarque: |
|||
|
|
(fait partie des globules blancs: leucocytes) |
 |
10 à 12 µm de Ø, 45 à 70% des leucocytes (3 à 6000 cellules/µL), doués de diapédèse et de pouvoir de phagocytose (lysosomes), très mobiles ce sont les principaux phagocytes intervenant dans la réponse immunitaire non spécifique, leur demi-vie est de l'ordre de 4 à 10 heures une fois qu'ils ont quitté la moelle osseuse, sous l'action de substances chimiotactiques, ils quittent le sang et se rendent sur les lieux de l'inflammation, RNC assez élevé |
|
|
|
(fait partie des globules blancs: leucocytes mais peut aussi être tissulaire) |
10 à 12 µm de Ø, 1 à 3% des leucocytes, doués de diapédèse, contiennent de nombreux lysosomes, interviennent essentiellement dans la réponse contre les parasites par leurs propriétés cytotoxiques |
||
|
(fait partie des globules blancs: leucocytes) |
8 à 10 µm de Ø, moins de 0,5% des leucocytes, noyau rond, doués de diapédèse, possèdent de grosses granulations pouvant libérer de l'histamine (voir inflammation), jouent un rôle dans l'allergie |
|||
|
|
8 à 10 µm de Ø, localisés dans les tissus (tissu conjonctif et muqueuses) |
|||
|
lignée lymphoïde |
(fait partie des globules blancs: leucocytes au sens large) |
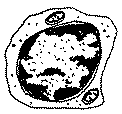 |
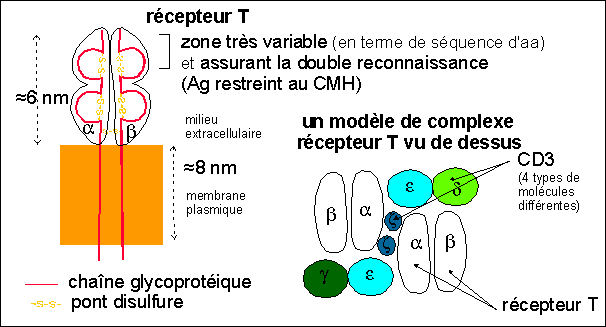
7 à 8 µm de Ø, 15 à 35% des leucocytes, doués de diapédèse, RNC élevé (peut se diviser activement), responsables de l'immunité à médiation cellulaire Remarque: |
|
|
|
(fait partie des globules blancs: leucocytes) |
 |
7 à 8 µm de Ø, 5 à 15% des leucocytes, doués de diapédèse, RNC élevé (peut se diviser activement) |
|
|
|
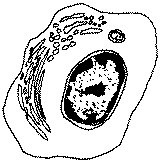 |
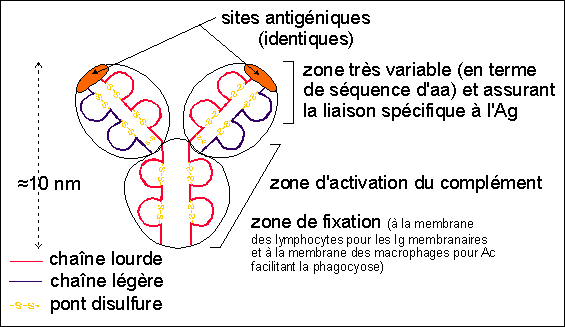
issu de la différenciation des lymphocytes B, Ø > 20µm, RNC faible (cellule différenciée, spécialisée : REG et Golgi hyperdéveloppés), cellule sécrétrice d'Ig (immunoglobulines) ou Ac (anticorps), responsables de l'immunité à médiation humorale |
||
|
cellule souche |
|
cellule tueuse K (killer) ou NK (natural killer)... |
||
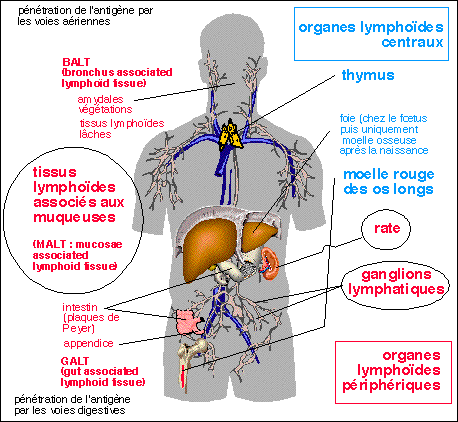
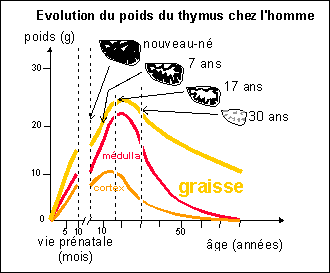
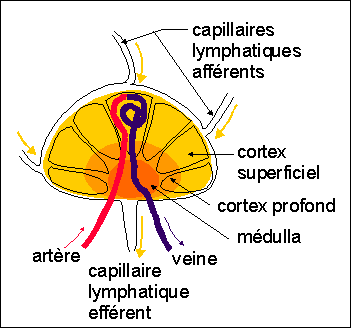
Le système immunitaire
comprend classiquement les organes centraux ou primaires (thymus
et moelle rouge des os longs) et les organes
périphériques ou secondaires (ganglions
lymphatiques, rate et organes lymphoïdes
associés aux muqueuses) mais aussi les appareils
circulatoires (sanguin et lymphatique).