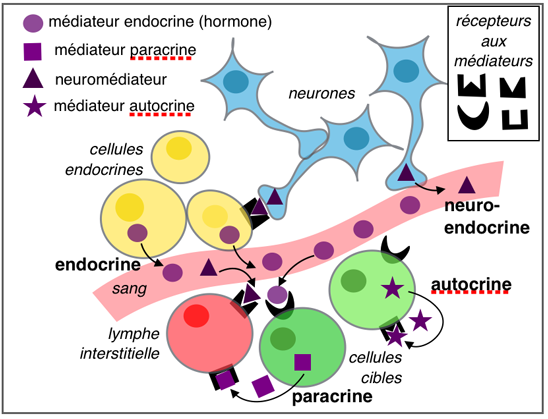
 Dans
cette page et celle sur les hormones
stéroïdes et les prostaglandines, nous verrons les
quatres types de médiateurs ou substances
chimiques informatives.
Dans
cette page et celle sur les hormones
stéroïdes et les prostaglandines, nous verrons les
quatres types de médiateurs ou substances
chimiques informatives.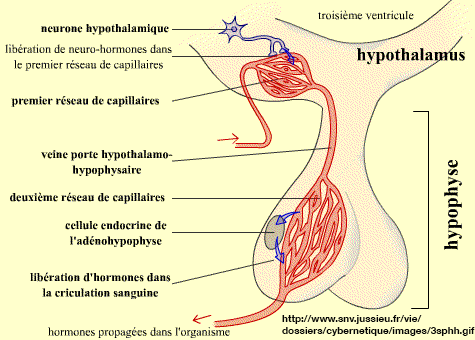 On désigne par système hypothalamo-hypophysaire un complexe
formé par des centres nerveux de l'hypothalamus
(appelés "noyaux") et la partie antérieure de l'hypophyse (antehypophyse)
ou adénohypophyse qui est une glande endocrine
(c'est-à-dire qui sécréte des médiateurs
endocrines ou hormones).
On désigne par système hypothalamo-hypophysaire un complexe
formé par des centres nerveux de l'hypothalamus
(appelés "noyaux") et la partie antérieure de l'hypophyse (antehypophyse)
ou adénohypophyse qui est une glande endocrine
(c'est-à-dire qui sécréte des médiateurs
endocrines ou hormones).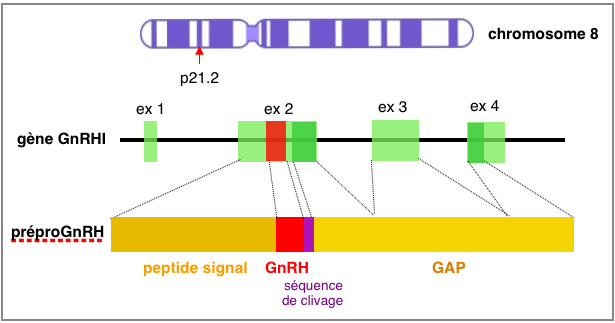 La
GnRH ou LHRH est un décapeptide qui est libéré
à partir d'un peptide de bien plus grande taille (92 aa): la
préproGnRH, synthétisée à partir d'un
gène situé sur le bras court du chromosome 8 humain (⎋).
Le gène codant la pré-pro-GnRH est situé sur le
brin reverse de l'ADN et est formé de 4 exons et 3 introns. La
pré-pro-GnRH est constituée d’une séquence
peptide signal, la séquence de la GnRH et une séquence de
clivage GKR (Gly-Lys-Arg), toutes codées par l’exon 2, et
du GnRH associated peptide (GAP, codé par une partie de
l’exon 2, l’exon 3 et une partie de l’exon 4). Parmi
les mammifères, seul le cobaye présente un acide
aminé différent dans la séquence de la GnRH. La
séquence du GAP est en revanche beaucoup plus variable.
Après clivage du peptide signal et du GAP, la GnRH subit deux
modifications importantes : la cyclisation de la glutamine N terminale
en pyro-glutamine et l’amidation de l’acide glutamique C
terminal. Ces deux modifications post-traductionnelles empêchent
l’action des exopeptidases. La structure de la GnRH lui
confère une conformation spatiale en fer à cheval
importante pour la liaison à son récepteur. La GnRH et le
GAP sont tous les deux présents dans les vésicules de
sécrétion des neurones à GnRH. Les neurones
à GnRH ont une origine embryonnaire extracérébrale
et migrent précocement depuis la zone antérieure nasale
vers l'aire préoptique de l'hypothalamus mais restent sous forme
d'amas - de quelques centaines de cellules - dispersés,
mais dont les terminaisons nerveuses se situent au niveau de
l'éminence médiane de l'hypothalamus. Les neurones
à GnRH comportent un grand nombre de dendrites qui semblent
fluctuer - en nombre et en taille - au cours des cycles sexuels,
notamment sous l'action des œstrogènes. Des cellules
gliales spécialisées (tanicytes) semblent faciliter la
réorganisation périodique du réseau des
prolongements axonaux en liaison avec l'endothélium
fenestré des vaisseaux du réseau capillaire qui prend en
charge la GnRH libérée. C'est probablement à ce
niveau et non au niveau des corps cellulaires, que se fait le
contrôle de la sécrétion pulsatile, encore
mystérieuse.
La
GnRH ou LHRH est un décapeptide qui est libéré
à partir d'un peptide de bien plus grande taille (92 aa): la
préproGnRH, synthétisée à partir d'un
gène situé sur le bras court du chromosome 8 humain (⎋).
Le gène codant la pré-pro-GnRH est situé sur le
brin reverse de l'ADN et est formé de 4 exons et 3 introns. La
pré-pro-GnRH est constituée d’une séquence
peptide signal, la séquence de la GnRH et une séquence de
clivage GKR (Gly-Lys-Arg), toutes codées par l’exon 2, et
du GnRH associated peptide (GAP, codé par une partie de
l’exon 2, l’exon 3 et une partie de l’exon 4). Parmi
les mammifères, seul le cobaye présente un acide
aminé différent dans la séquence de la GnRH. La
séquence du GAP est en revanche beaucoup plus variable.
Après clivage du peptide signal et du GAP, la GnRH subit deux
modifications importantes : la cyclisation de la glutamine N terminale
en pyro-glutamine et l’amidation de l’acide glutamique C
terminal. Ces deux modifications post-traductionnelles empêchent
l’action des exopeptidases. La structure de la GnRH lui
confère une conformation spatiale en fer à cheval
importante pour la liaison à son récepteur. La GnRH et le
GAP sont tous les deux présents dans les vésicules de
sécrétion des neurones à GnRH. Les neurones
à GnRH ont une origine embryonnaire extracérébrale
et migrent précocement depuis la zone antérieure nasale
vers l'aire préoptique de l'hypothalamus mais restent sous forme
d'amas - de quelques centaines de cellules - dispersés,
mais dont les terminaisons nerveuses se situent au niveau de
l'éminence médiane de l'hypothalamus. Les neurones
à GnRH comportent un grand nombre de dendrites qui semblent
fluctuer - en nombre et en taille - au cours des cycles sexuels,
notamment sous l'action des œstrogènes. Des cellules
gliales spécialisées (tanicytes) semblent faciliter la
réorganisation périodique du réseau des
prolongements axonaux en liaison avec l'endothélium
fenestré des vaisseaux du réseau capillaire qui prend en
charge la GnRH libérée. C'est probablement à ce
niveau et non au niveau des corps cellulaires, que se fait le
contrôle de la sécrétion pulsatile, encore
mystérieuse.
 L'implication
de ce gène dans la fonction de réproduction ne sera
revélée que 10 ans plus tard. En 2003 on a
découvert qu'aussi bien chez la souris que dans
l’espèce humaine, une mutation perte de fonction ou une
délétion de KISS1R entraîne une absence de
puberté, des gonades de petite taille, des concentrations
plasmatiques des hormones sexuelles (œstrogènes et
testostérone) et des gonadotrophines basses, et une
infertilité chez l’adulte. Toutefois, un traitement par les
gonadotrophines ou par GnRH permet une ovulation et une grossesse
normale chez les femmes mutées et restaure la
spermatogenèse chez les hommes mutés. Cela démontre
que les kisspeptines sont nécessaires au bon
déroulement de la puberté et de la fonction de
reproduction chez l’adulte, mais ne participent pas au
développement des gonades. On a rapidement
montré que les kisspeptines contrôlent la
libération de la GnRH.
L'implication
de ce gène dans la fonction de réproduction ne sera
revélée que 10 ans plus tard. En 2003 on a
découvert qu'aussi bien chez la souris que dans
l’espèce humaine, une mutation perte de fonction ou une
délétion de KISS1R entraîne une absence de
puberté, des gonades de petite taille, des concentrations
plasmatiques des hormones sexuelles (œstrogènes et
testostérone) et des gonadotrophines basses, et une
infertilité chez l’adulte. Toutefois, un traitement par les
gonadotrophines ou par GnRH permet une ovulation et une grossesse
normale chez les femmes mutées et restaure la
spermatogenèse chez les hommes mutés. Cela démontre
que les kisspeptines sont nécessaires au bon
déroulement de la puberté et de la fonction de
reproduction chez l’adulte, mais ne participent pas au
développement des gonades. On a rapidement
montré que les kisspeptines contrôlent la
libération de la GnRH. 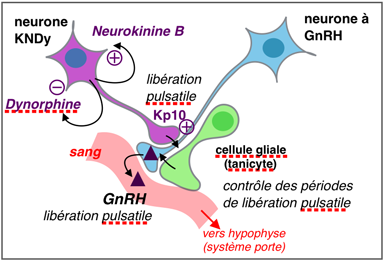 Chez
l’homme, l'unique gène KISS-1 est traduit en une
pro-hormone de 145 acides aminés qui est ensuite clivée
pour donner la forme longue des kisspeptines, une molécule de 54
AA, dont l’extrémité C-terminale est amidée.
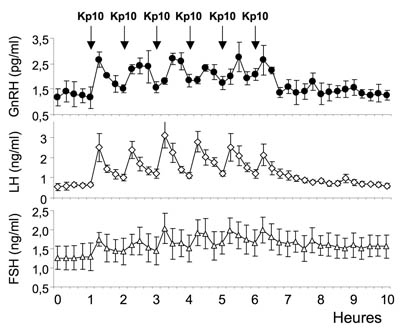
Kp54 peut donner naissance à des fragments plus courts (Kp14,
Kp13 et Kp10), biologiquement actifs. (<--- image ci-contre :
Chez
l’homme, l'unique gène KISS-1 est traduit en une
pro-hormone de 145 acides aminés qui est ensuite clivée
pour donner la forme longue des kisspeptines, une molécule de 54
AA, dont l’extrémité C-terminale est amidée.
Kp54 peut donner naissance à des fragments plus courts (Kp14,
Kp13 et Kp10), biologiquement actifs. (<--- image ci-contre : La GnRH ( 1YY1 ⎋ le fichier contient 21 modèles synthétiques accessibles dans le menu JSmol) est une neurohormone de type décapeptide (10aa) dont le premier aa - l'acide pyroglutamique (pGlu) remplace l'acide glutamique (Glu) -, et le dernier aa - la glycine possède un groupement amide terminal - sont modifiés :
Chez les différents vertébrés la séquence du GnRH (qui est un facteur de croissance : growth factor plutôt qu'un hormone au sen strict) varie de quelques aa et est sous le contrôle de 3 gènes. La séquence de la GnRH intervenant dans la reproduction est :
pGlu - His - Trp - Ser - Tyr - Gly - Leu - Arg - Pro - Gly-NH2
pGlu indique la pyroglutamine et Gly-NH2 indique l'amidation de la glycine terminale (extrêmité -COOH du polypeptide qui devient -CO-NH2).
La GnRH se fixe a un récepteur membranaire de type glycoprotéine à la surface des cellules gonadotropes. Le complexe hormone-récepteur est endocyté après micro-agrégation (regroupement des récepteurs deux par deux) et la GnRH est libérée dans la cellule. Elle semble agir au niveau du cytoplasme et du noyau et provoque rapidement une libération des gonadotropines et, à plus long terme, une stimulation de la synthèse des gonadotropines (par l'AMPc).
Les gonadotrophines (LH et FSH) sont toutes deux des glycoprotéines formées de deux sous-unités* : la sous-unité* alpha est commune aux deux molécules (ainsi qu'à l'HCG, qui intervient lors de la grossesse, et à d'autres hormones...). La sous-unité* béta est donc spécifique de chaque hormone (pour des données évolutives sur la famille multigénique voir access-ens lyon).
On ne connaît pas toutes les structures spatiales de toutes ces molécules : voici celles trouvées.
- unique sous-unité* α (comportant deux chaînes* A et B) de l''HCG (Human chorionic gonadotrophin, voir ci-dessous, 1HCN); cette sous-unité α est identique pour la LH et la FSH ;
- modèle théorique du précuseur de la sous-unité* β d'une LH (construit artificiellement et non protéine cristallisée, 1M92) ;
- FSH complète (avec la sous-unité* α (chaînes* A et C) et sous-unité* β (chaînes* B et D), 1FL7) et FSH avec son récepteur (avec la sous-unité* α (chaînes* A et D), la sous-unité* β (chaînes* B et E), et le récepteur (avec ses deux chaînes* C et F ), 1XWD).
* en anglais les sous-unités ET les chaînes sont apellées "chains", seules les lettres grecques (pour les sous-unités) et romanes (pour les chaînes) permettent de les différencier.
| Quelques
commandes pour JSmol: Couleurs des groupes non aa (glucides modifiés : hétérosides): cpk, rouge Modèles des chaînes peptidiques :, squelette, boules et bâtons, cartoons Couleurs des aa: cpk - a.a. colorés, chaînes colorées (couleurs de références page JSmol) |